
Genome-Wide Identification and Analysis of the NAC Transcription Factor Gene Family in Garden Asparagus (Asparagus officinalis)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the NAC Gene Family in A. officinalis
2.2. Phylogenetic Analysis and Classification of AoNAC Genes
2.3. Chromosomal Mapping and Cis-Acting Regulatory Analysis of AoNAC Genes
2.4. Gene Structure and Motif Analysis of AoNAC Genes
2.5. Physicochemical Properties and Subcellular Localization Analysis of AoNAC Genes
2.6. Gene Duplication and Syntenic Analysis of AoNAC Genes
2.7. Differential Expression of AoNAC Genes under Different Types of Salinity Stress
3. Results
3.1. Identification of NAC Members in A. officinalis
3.2. Phylogenetic Analysis of AoNAC and AtNAC
3.3. Gene Structure, Conserved Motifs, and Domain Analysis of AoNAC Genes
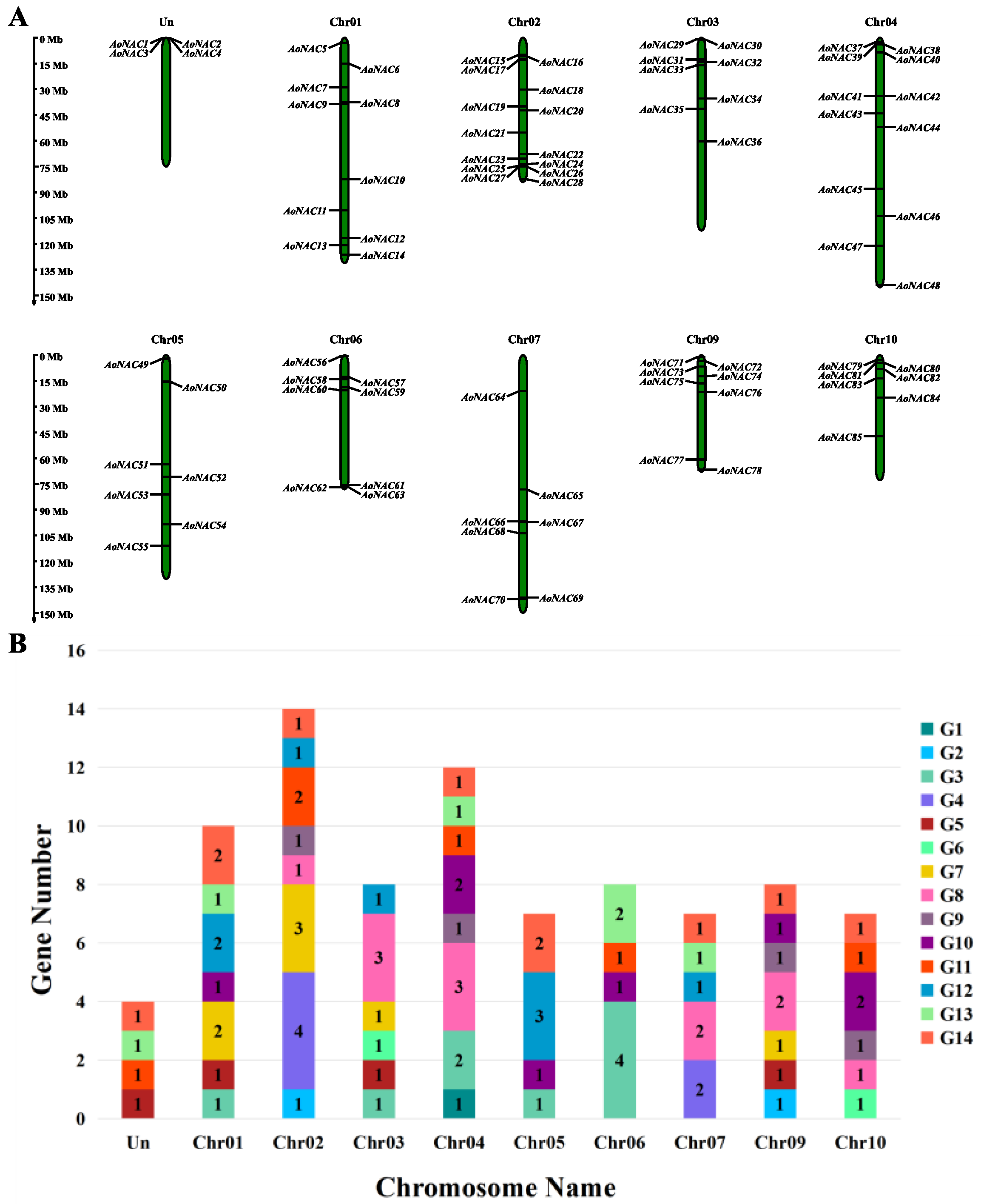
3.4. Chromosomal Mapping and Cis-Acting Regulatory Analysis of AoNAC Genes
3.5. Gene Duplication and Syntenic Analysis of AoNAC Genes
3.6. Differentially Expressed Genes Analysis of AoNAC Genes under Different Types of Salinity Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495. [Google Scholar] [CrossRef] [PubMed]
- Meshi, T.; Iwabuchi, M. Plant transcription factors. Plant Cell Physiol. 1995, 36, 1405–1420. [Google Scholar] [PubMed]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.P.; Zhang, H.; Kong, L.; Gao, G.; Luo, J.C. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2014, 12, D1182–D1187. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.S.; Li, H.L.; Grierson, D.; Fu, D.Q. NAC transcription factor family regulation of fruit ripening and quality: A review. Cells 2022, 11, 525. [Google Scholar] [CrossRef]
- Tran, L.S.; Quach, T.N.; Guttikonda, S.K.; Aldrich, D.L.; Kumar, R.; Neelakandan, A.; Valliyodan, B.; Nguyen, H.T. Molecular characterization of stress–inducible GmNAC genes in soybean. Mol. Genet. Genom. 2009, 281, 647–664. [Google Scholar] [CrossRef]
- Zhu, T.T.; Nevo, E.; Sun, D.F.; Peng, J.H. Phylogenetic analyses unravel the evolutionary history of NAC proteins in plants. Evolution 2012, 66, 1833–1848. [Google Scholar] [CrossRef]
- Duval, M.; Hsieh, T.F.; Kim, S.Y.; Thomas, T.L. Molecular characterization of AtNAM: A member of the Arabidopsis NAC domain superfamily. Plant Mol. Biol. 2002, 50, 237–248. [Google Scholar] [CrossRef]
- Shahzad, R.; Jamil, S.; Ahmad, S.; Nisar, A.; Amina, Z.; Saleem, S.; Iqbal, M.Z.; Atif, R.M.; Wang, X. Harnessing the potential of plant transcription factors in developing climate resilient crops to improve global food security: Current and future perspectives. Saudi J. Biol. Sci. 2021, 28, 2323–2341. [Google Scholar] [CrossRef]
- Hegedus, D.; Yu, M.; Baldwin, D.; Gruber, M.; Sharpe, A.; Parkin, I.; Whitwill, S.; Lydiate, D. Molecular characterization of Brassica napus NAC domain transcriptional activators induced in response to biotic and abiotic stress. Plant Mol. Biol. 2003, 53, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; Galberg, P.; Petersen, K.; O’shea, C.; Skriver, K. The Arabidopsis thaliana NAC transcription factor family: Structure–function relationships and determinants of ANAC019 stress signalling. Biochem. J. 2010, 426, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Kjaersgaard, T.; Jensen, M.K.; Christiansen, M.W.; Gregersen, P.; Kragelund, B.B.; Skriver, K. Senescence–associated barley NAC (NAM, ATAF1,2, CUC) transcription factor interacts with radical–induced cell death 1 through a disordered regulatory domain. J. Biol. Chem. 2011, 286, 35418–35429. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.J.; Kim, S.G.; Park, C.M. Membrane–bound transcription factors in plants. Trends Plant Sci. 2008, 13, 550–556. [Google Scholar] [CrossRef]
- Sun, Q.W.; Huang, J.F.; Guo, Y.F.; Yang, M.M.; Guo, Y.J.; Li, J.; Zhang, J.; Xu, W.L. A cotton NAC domain transcription factor, GhFSN5, negatively regulates secondary cell wall biosynthesis and anther development in transgenic Arabidopsis. Plant Physiol. Bioch. 2020, 146, 303–314. [Google Scholar] [CrossRef]
- Vroemen, C.W.; Mordhorst, A.P.; Albrecht, C.; Kwaaitaal, M.A.; de Vries, S.C. The CUP-SHAPED COTYLEDON3 gene is required for boundary and shoot meristem formation in Arabidopsis. Plant Cell 2003, 15, 1563–1577. [Google Scholar] [CrossRef]
- Zimmermann, R.; Werr, W. Pattern formation in the monocot embryo as revealed by NAMand CUC3 orthologues from Zea mays L. Plant Mol. Biol. 2005, 58, 669–685. [Google Scholar] [CrossRef]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef]
- Guo, Y.; Gan, S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC Gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Kim, S.G.; Park, J.E.; Park, H.Y.; Lim, M.H.; Chua, N.H.; Park, C.M. A membrane-bound NAC transcription factor regulates cell division in Arabidopsis. Plant Cell 2006, 18, 3132–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, V.; Bauch, M.; Bennett, T.; Campilho, A.; Wolkenfelt, H.; Jian, X.; Scheres, B. The NAC domain transcription factors FEZ and SOMBRERO control the orientation of cell division plane in Arabidopsis root stem cells. Dev. Cell 2008, 15, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Mao, C.J.; He, J.M.; Liu, L.N.; Deng, Q.M.; Yao, X.F.; Liu, C.M.; Qiao, Y.L.; Li, P.; Ming, F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol. J. 2020, 18, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, S.; Wang, Y.; Zhang, X.; Ming, F. OsNAC2 encoding a NAC transcription factor that affects plant height through mediating the gibberellic acid pathway in rice. Plant J. 2015, 82, 302–314. [Google Scholar] [CrossRef]
- Jensen, M.K.; Lindemose, S.; Masi, F.D.; Reimer, J.J.; Nielsen, M.; Perera, V.; Workman, C.T.; Turck, F.; Grant, M.R.; Mundy, J.; et al. ATAF1 transcription factor directly regulates abscisic acid biosynthetic gene NCED3 in Arabidopsis thaliana. FEBS Open Bio. 2013, 3, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.L.; Chen, N.Z.; Rui, A.; Su, Z.; Qi, B.S.; Ren, F.; Chen, J.; Wang, X.C. A novel drought-inducible gene, ATAF1, encodes a NAC family protein that negatively regulates the expression of stress-responsive genes in Arabidopsis. Plant Mol. Biol. 2007, 63, 289–305. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, B.; Lu, G.; Han, B. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem. Biophys. Res. Commun. 2009, 379, 985–989. [Google Scholar] [CrossRef]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Deyholos, M.K. Comprehensive transcriptional profiling of NaCl-stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol. 2006, 6, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collinge, M.; Boller, T. Differential induction of two potato genes, Stprx2 and StNAC, in response to infection by Phytophthora infestans and to wounding. Plant Mol. Biol. 2001, 46, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Qu, F.; Morris, T.J. HRT gene function requires interaction between a NAC protein and viral capsid protein to confer resistance to turnip crinkle virus. Plant Cell 2000, 12, 1917–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delessert, C.; Kazan, K.; Wilson, I.W.; Straeten, D.V.D.; Manners, J.; Dennis, E.S.; Dolferus, R. The transcription factor ATAF2 represses the expression of pathogenesis-related genes in Arabidopsis. Plant J. 2005, 43, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Basnayake, B.M.; Zhang, H.; Li, G.; Li, W.; Virk, N.; Mengiste, T.; Song, F. The Arabidopsis ATAF1, a NAC transcription factor, is a negative regulator of defense responses against necrotrophic fungal and bacterial pathogens. Mol. Plant-Microbe Interact. 2009, 22, 1227–1238. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.F.R.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef]
- Xue, G.P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Fang, Y.J.; Jun, Y.; Xie, K.B.; Xiong, L.Z. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.B.; Guang, Q.; Kong, Y.Z.; Kong, D.J.; Gao, Q.; Zhou, G.K. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, C.V.; Esfahani, M.N.; Watanabe, Y.; Tran, U.T.; Sulieman, S.; Mochida, K.; Nguyen, D.V.; Tran, L.S. Genome-wide identification and expression analysis of the CaNAC family members in chickpea during development, dehydration and ABA treatments. PLoS ONE 2014, 9, e114107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Li, D.H.; Wang, Y.Y.; Zhou, R.; Wang, L.H.; Zhang, Y.X.; Yu, J.Y.; Gong, H.H.; You, J.; Zhang, X.R.; et al. Genome-wide identification and comprehensive analysis of the NAC transcription factor family in Sesamum indicum. PLoS ONE 2018, 13, e0199262. [Google Scholar] [CrossRef]
- Gong, X.; Zhao, L.Y.; Song, X.F.; Lin, Z.K.; Gu, B.J.; Yan, J.X.; Zhang, S.L.; Tao, S.T.; Huang, X.S. Genome-wide analyses and expression patterns under abiotic stress of NAC transcription factors in white pear (Pyrus bretschneideri). BMC Plant Biol. 2019, 19, 161. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lin, X.; Zhang, D.; Li, Q.; Chen, S. Genome-wide analysis of NAC gene family in Betula pendula. Forests 2019, 10, 741. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Liu, Y.; Zhang, M.; Bai, M.; Qin, Y. Genome-wide identification and expression analysis of the NAC transcription factor family in pineapple. Trop. Plant Biol. 2019, 12, 255–267. [Google Scholar] [CrossRef]
- Wang, N.; Liu, H.; Li, Z.; Lu, L.; Wang, C. The bioactive compounds and biological functions of Asparagus officinalis L.—A review. J. Funct. Foods. 2019, 65, 03727. [Google Scholar]
- Pegiou, E.; Mumm, R.; Acharya, P.; de Vos, R.; Hall, R.D. Green and White asparagus (Asparagus officinalis): A Source of Developmental, Chemical and Urinary Intrigue. Metabolites 2019, 10, 17. [Google Scholar] [CrossRef]
- Global Production of Vegetables in 2020, by Type. 2022. Available online: https://www.statista.com/statistics/264065/global-production-of-vegetables-by-type (accessed on 22 April 2022).
- Sriyab, S.; Laosirisathian, N.; Punyoyai, C.; Anuchapreeda, S.; Tima, S.; Chiampanichayakul, S.; Chaiyana, W. Nutricosmetic effects of Asparagus officinalis: A potent matrix metalloproteinase-1 inhibitor. Sci. Rep. 2021, 11, 8772. [Google Scholar] [CrossRef]
- Rui, M.; Ricardo, S. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 13. [Google Scholar]
- Zhang, X.; Han, C.; Cao, Y. Transcriptomic and Physiological analyses reveal the dynamic response to salinity stress of the garden asparagus (Asparagus officinalis L.). Plant Mol. Biol. Report. 2020, 38, 613–627. [Google Scholar] [CrossRef]
- Harkess, A.; Zhou, J.; Xu, C.; Bowers, J.E.; Van der Hulst, R.; Ayyampalayam, S.; Mercati, F.; Riccardi, P.; McKain, M.R.; Kakrana, A.; et al. The asparagus genome sheds light on the origin and evolution of a young Y chromosome. Nat. Commun. 2017, 8, 1279. [Google Scholar] [CrossRef] [PubMed]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Yu, G. Using ggtree to visualize data on tree-like structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Wang, D.P.; Zhang, Y.B.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Han, C.; Gao, H.; Cao, Y. Comparative transcriptome analysis of the garden asparagus (Asparagus officinalis L.) reveals the molecular mechanism for growth with arbuscular mycorrhizal fungi under salinity stress. Plant Physiol. Biochem. 2019, 141, 20–29. [Google Scholar] [CrossRef]
- Andrew, S. FastQC: A quality control tool for high throughput sequence data. Babraham Bioinform. 2014, 532, 1. [Google Scholar]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Bioch. 2018, 129, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Pressman, E.; Schaffer, A.A.; Compton, D.; Zmski, E. Carbohydrate content of young asparagus plants affected by temperature regimes. J. Plant Physiol. 1994, 143, 621–624. [Google Scholar] [CrossRef]
- Murase, K.; Shigenobu, S.; Fujii, S.; Ueda, K.; Murata, T.; Sakamoto, A.; Wada, Y.; Yamaguchi, K.; Osakabe, Y.; Osakabe, K.; et al. MYB transcription factor gene involved in sex determination in Asparagus officinalis. Genes Cells 2017, 22, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Dong, T.; Han, R.; Yu, J.; Zhu, M.; Zhang, Y.; Gong, Y.; Li, Z. Anthocyanins accumulation and molecular analysis of correlated genes by metabolome and transcriptome in green and purple asparaguses (Asparagus officinalis L.). Food Chem. 2019, 271, 18–28. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Guo, J.; Wang, S.; Yu, X.; Dong, R.; Li, Y.; Mei, X.; Shen, Y. Polyamines regulate strawberry fruit ripening by abscisic acid, auxin, and ethylene. Plant Physiol. 2018, 177, 339–351. [Google Scholar] [CrossRef] [Green Version]
- An, X.H.; Tian, Y.; Chen, K.Q.; Liu, X.J.; Liu, D.D.; Xie, X.B.; Cheng, C.G.; Cong, P.H.; Hao, Y.J. MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 2015, 56, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.B.; Bianchetti, R.E.; Alves, F.R.R.; Purgatto, E.; Peres, L.E.P.; Rossi, M.; Freschi, L. Light, Ethylene and Auxin Signaling Interaction Regulates Carotenoid Biosynthesis During Tomato Fruit Ripening. Front Plant Sci. 2018, 9, 1370. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, R.; Jiang, Q.; Sun, X.; Zhang, H.; Hu, Z. GmNAC06, a NAC domain transcription factor enhances salt stress tolerance in soybean. Plant Mol. Biol. 2021, 105, 333–345. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor enhances salt stress tolerance by modulating the ethylene response. Physiol. Plant. 2018, 164, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | pI | MW (kDa) | Length (aa) | Instability Index | Aliphatic Index | Subcellular Localization | ORF | Duplications |
|---|---|---|---|---|---|---|---|---|
| AoNAC1 | 4.49 | 50.35 | 449 | 45.65 | 63.63 | Nucleus | 1350 | Dispersed |
| AoNAC2 | 5.18 | 46.29 | 412 | 48.10 | 70.49 | Nucleus | 1239 | Dispersed |
| AoNAC3 | 7.68 | 28.42 | 249 | 40.89 | 70.12 | Nucleus | 750 | Dispersed |
| AoNAC4 | 5.51 | 26.46 | 232 | 24.75 | 57.11 | Nucleus | 699 | Dispersed |
| AoNAC5 | 5.42 | 99.26 | 872 | 57.24 | 71.69 | Nucleus | 2619 | Dispersed |
| AoNAC6 | 5.38 | 29.65 | 263 | 39.13 | 60.72 | Nucleus | 792 | Dispersed |
| AoNAC7 | 7.38 | 36.74 | 330 | 44.47 | 65.00 | Nucleus | 993 | Dispersed |
| AoNAC8 | 8.36 | 35.47 | 318 | 43.20 | 61.32 | Nucleus | 957 | Dispersed |
| AoNAC9 | 7.03 | 37.60 | 338 | 44.81 | 69.56 | Nucleus | 1017 | Dispersed |
| AoNAC10 | 7.35 | 43.48 | 389 | 47.84 | 61.13 | Nucleus | 1167 | Dispersed |
| AoNAC11 | 7.71 | 35.90 | 312 | 39.64 | 63.46 | Nucleus | 939 | Dispersed |
| AoNAC12 | 8.54 | 32.34 | 284 | 30.05 | 58.31 | Nucleus | 855 | Dispersed |
| AoNAC13 | 9.33 | 18.93 | 161 | 35.46 | 55.78 | Nucleus | 486 | Dispersed |
| AoNAC14 | 4.82 | 69.33 | 621 | 54.87 | 69.73 | Endomembrane System | 1866 | Dispersed |
| AoNAC15 | 5.95 | 35.97 | 313 | 57.04 | 68.18 | Nucleus | 942 | Dispersed |
| AoNAC16 | 6.36 | 31.38 | 279 | 41.39 | 55.63 | Nucleus | 840 | Dispersed |
| AoNAC17 | 9.17 | 21.54 | 197 | 61.85 | 60.46 | Nucleus | 594 | WGD/Segmental |
| AoNAC18 | 5.49 | 17.86 | 153 | 72.43 | 74.51 | Nucleus | 462 | Dispersed |
| AoNAC19 | 4.94 | 48.28 | 425 | 60.37 | 64.05 | Nucleus | 1278 | Dispersed |
| AoNAC20 | 5.30 | 36.19 | 318 | 44.07 | 64.69 | Nucleus | 957 | Dispersed |
| AoNAC21 | 6.47 | 31.90 | 280 | 44.93 | 58.57 | Nucleus | 843 | Dispersed |
| AoNAC22 | 4.84 | 66.29 | 589 | 53.84 | 69.56 | Endomembrane System | 1770 | Dispersed |
| AoNAC23 | 6.91 | 30.50 | 273 | 56.59 | 57.58 | Nucleus | 822 | Dispersed |
| AoNAC24 | 8.94 | 30.04 | 261 | 45.37 | 62.03 | Nucleus | 786 | Dispersed |
| AoNAC25 | 5.81 | 29.51 | 261 | 61.51 | 68.70 | Nucleus | 786 | Dispersed |
| AoNAC26 | 5.18 | 17.86 | 156 | 55.88 | 76.79 | Nucleus | 471 | Dispersed |
| AoNAC27 | 5.43 | 43.32 | 386 | 54.38 | 63.73 | Nucleus | 1161 | Proximal |
| AoNAC28 | 6.44 | 131.61 | 1187 | 53.51 | 70.73 | Nucleus | 3564 | Tandem |
| AoNAC29 | 5.93 | 72.81 | 652 | 46.37 | 78.70 | Plasma Membrane | 1959 | WGD/Segmental |
| AoNAC30 | 6.47 | 42.88 | 383 | 35.15 | 60.34 | Nucleus | 1152 | Dispersed |
| AoNAC31 | 9.73 | 23.79 | 209 | 45.91 | 56.08 | Chloroplast | 630 | Dispersed |
| AoNAC32 | 5.76 | 35.25 | 304 | 42.87 | 59.61 | Nucleus | 915 | Dispersed |
| AoNAC33 | 4.67 | 13.56 | 121 | 46.42 | 43.55 | Nucleus | 366 | Dispersed |
| AoNAC34 | 5.84 | 127.90 | 1139 | 58.08 | 74.12 | Nucleus | 3420 | Dispersed |
| AoNAC35 | 9.08 | 23.61 | 205 | 35.37 | 70.83 | Nucleus | 618 | Dispersed |
| AoNAC36 | 10.05 | 27.51 | 261 | 40.66 | 59.20 | Nucleus | 786 | Dispersed |
| AoNAC37 | 5.78 | 29.86 | 263 | 54.69 | 61.86 | Nucleus | 792 | Dispersed |
| AoNAC38 | 5.28 | 48.40 | 423 | 49.84 | 62.53 | Nucleus | 1272 | Tandem |
| AoNAC39 | 6.49 | 47.18 | 415 | 53.29 | 57.42 | Nucleus | 1248 | Tandem |
| AoNAC40 | 4.52 | 42.57 | 373 | 49.98 | 66.89 | Nucleus | 1122 | Dispersed |
| AoNAC41 | 5.71 | 64.44 | 563 | 46.69 | 73.53 | Nucleus | 1692 | Proximal |
| AoNAC42 | 4.93 | 27.97 | 243 | 41.99 | 62.55 | Nucleus | 732 | Proximal |
| AoNAC43 | 9.02 | 25.18 | 221 | 44.43 | 56.88 | Nucleus | 666 | WGD/Segmental |
| AoNAC44 | 5.04 | 42.46 | 377 | 47.30 | 60.24 | Nucleus | 1134 | Dispersed |
| AoNAC45 | 4.70 | 25.48 | 224 | 49.37 | 77.50 | Nucleus | 675 | Dispersed |
| AoNAC46 | 7.07 | 33.31 | 293 | 56.01 | 60.31 | Nucleus | 882 | Dispersed |
| AoNAC47 | 4.91 | 21.99 | 191 | 47.58 | 63.30 | Nucleus | 576 | Dispersed |
| AoNAC48 | 9.11 | 24.52 | 205 | 30.90 | 58.44 | Nucleus | 618 | Dispersed |
| AoNAC49 | 4.83 | 37.71 | 338 | 67.48 | 69.56 | Nucleus | 1017 | Dispersed |
| AoNAC50 | 7.02 | 32.15 | 278 | 33.72 | 56.47 | Nucleus | 837 | Dispersed |
| AoNAC51 | 5.03 | 32.27 | 290 | 41.81 | 74.38 | Nucleus | 873 | Dispersed |
| AoNAC52 | 4.67 | 57.78 | 516 | 43.74 | 60.31 | Nucleus | 1551 | Dispersed |
| AoNAC53 | 9.10 | 36.03 | 315 | 40.01 | 78.03 | Nucleus | 948 | Dispersed |
| AoNAC54 | 9.44 | 27.50 | 245 | 38.85 | 61.76 | Nucleus | 738 | Dispersed |
| AoNAC55 | 9.46 | 20.95 | 186 | 48.11 | 81.24 | Chloroplast | 561 | Dispersed |
| AoNAC56 | 8.04 | 34.76 | 308 | 47.17 | 61.43 | Nucleus | 927 | Dispersed |
| AoNAC57 | 5.85 | 28.97 | 253 | 52.15 | 68.58 | Nucleus | 762 | Dispersed |
| AoNAC58 | 8.84 | 46.54 | 412 | 53.06 | 59.13 | Nucleus | 1239 | Dispersed |
| AoNAC59 | 5.87 | 10.34 | 90 | 48.36 | 75.78 | Nucleus | 273 | Dispersed |
| AoNAC60 | 9.27 | 13.36 | 113 | 46.58 | 48.32 | Chloroplast | 342 | Dispersed |
| AoNAC61 | 8.44 | 32.25 | 287 | 51.53 | 58.75 | Nucleus | 864 | WGD/Segmental |
| AoNAC62 | 6.36 | 78.39 | 685 | 53.96 | 64.89 | Nucleus | 2058 | Tandem |
| AoNAC63 | 5.44 | 15.78 | 134 | 39.32 | 90.90 | Nucleus | 405 | Tandem |
| AoNAC64 | 7.61 | 34.01 | 299 | 29.06 | 52.81 | Nucleus | 900 | Dispersed |
| AoNAC65 | 6.98 | 29.38 | 260 | 38.02 | 70.50 | Nucleus | 783 | Dispersed |
| AoNAC66 | 5.47 | 35.20 | 313 | 57.92 | 61.79 | Nucleus | 942 | Proximal |
| AoNAC67 | 4.67 | 45.48 | 404 | 57.87 | 69.33 | Nucleus | 1215 | Proximal |
| AoNAC68 | 6.67 | 40.15 | 356 | 44.86 | 71.24 | Nucleus | 1071 | Dispersed |
| AoNAC69 | 7.04 | 39.23 | 344 | 46.40 | 57.50 | Extracellular Space | 1035 | WGD/Segmental |
| AoNAC70 | 7.71 | 34.28 | 298 | 49.56 | 59.30 | Nucleus | 897 | WGD/Segmental |
| AoNAC71 | 6.11 | 29.64 | 266 | 65.67 | 48.08 | Nucleus | 801 | Dispersed |
| AoNAC72 | 8.88 | 13.70 | 117 | 71.07 | 64.10 | Chloroplast | 354 | Dispersed |
| AoNAC73 | 5.50 | 29.51 | 264 | 52.09 | 62.05 | Nucleus | 795 | Dispersed |
| AoNAC74 | 8.39 | 23.07 | 201 | 48.54 | 65.87 | Nucleus | 606 | Dispersed |
| AoNAC75 | 9.71 | 20.87 | 186 | 44.04 | 66.18 | Nucleus | 561 | WGD/Segmental |
| AoNAC76 | 5.51 | 36.10 | 314 | 43.35 | 61.82 | Nucleus | 945 | Dispersed |
| AoNAC77 | 9.53 | 24.87 | 219 | 46.45 | 59.22 | Nucleus | 660 | WGD/Segmental |
| AoNAC78 | 5.61 | 44.10 | 392 | 42.41 | 76.12 | Chloroplast | 1179 | Dispersed |
| AoNAC79 | 8.85 | 30.95 | 272 | 46.24 | 76.69 | Nucleus | 819 | Dispersed |
| AoNAC80 | 5.43 | 24.00 | 208 | 36.83 | 53.41 | Nucleus | 627 | Tandem |
| AoNAC81 | 5.24 | 64.63 | 567 | 46.76 | 71.55 | Nucleus | 1704 | Tandem |
| AoNAC82 | 8.55 | 26.80 | 234 | 57.68 | 67.91 | Nucleus | 705 | WGD/Segmental |
| AoNAC83 | 5.58 | 32.47 | 278 | 49.19 | 64.89 | Nucleus | 837 | Dispersed |
| AoNAC84 | 8.19 | 27.05 | 232 | 49.19 | 68.10 | Nucleus | 699 | Dispersed |
| AoNAC85 | 4.63 | 50.48 | 451 | 40.36 | 62.86 | Nucleus | 1356 | Dispersed |
| Tandem Duplication | Chromosome Name | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| AoNAC38 and AoNAC39 | Chr04 | 0.35 | 0.70 | 0.51 |
| AoNAC62 and AoNAC63 | Chr06 | 0.06 | 0.10 | 0.60 |
| AoNAC80 and AoNAC81 | Chr10 | 0.29 | 3.69 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Zhang, J.; Zhang, Q.; Dong, A.; Wu, Q.; Zhu, X.; Zhu, X. Genome-Wide Identification and Analysis of the NAC Transcription Factor Gene Family in Garden Asparagus (Asparagus officinalis). Genes 2022, 13, 976. https://doi.org/10.3390/genes13060976
Li C, Zhang J, Zhang Q, Dong A, Wu Q, Zhu X, Zhu X. Genome-Wide Identification and Analysis of the NAC Transcription Factor Gene Family in Garden Asparagus (Asparagus officinalis). Genes. 2022; 13(6):976. https://doi.org/10.3390/genes13060976
Chicago/Turabian StyleLi, Caifeng, Jingyang Zhang, Qianqian Zhang, Ang Dong, Qiuhong Wu, Xingyu Zhu, and Xuli Zhu. 2022. "Genome-Wide Identification and Analysis of the NAC Transcription Factor Gene Family in Garden Asparagus (Asparagus officinalis)" Genes 13, no. 6: 976. https://doi.org/10.3390/genes13060976
APA StyleLi, C., Zhang, J., Zhang, Q., Dong, A., Wu, Q., Zhu, X., & Zhu, X. (2022). Genome-Wide Identification and Analysis of the NAC Transcription Factor Gene Family in Garden Asparagus (Asparagus officinalis). Genes, 13(6), 976. https://doi.org/10.3390/genes13060976