
Genetic Variants Associated with Elevated Plasma Ceramides in Individuals with Metabolic Syndrome

Abstract
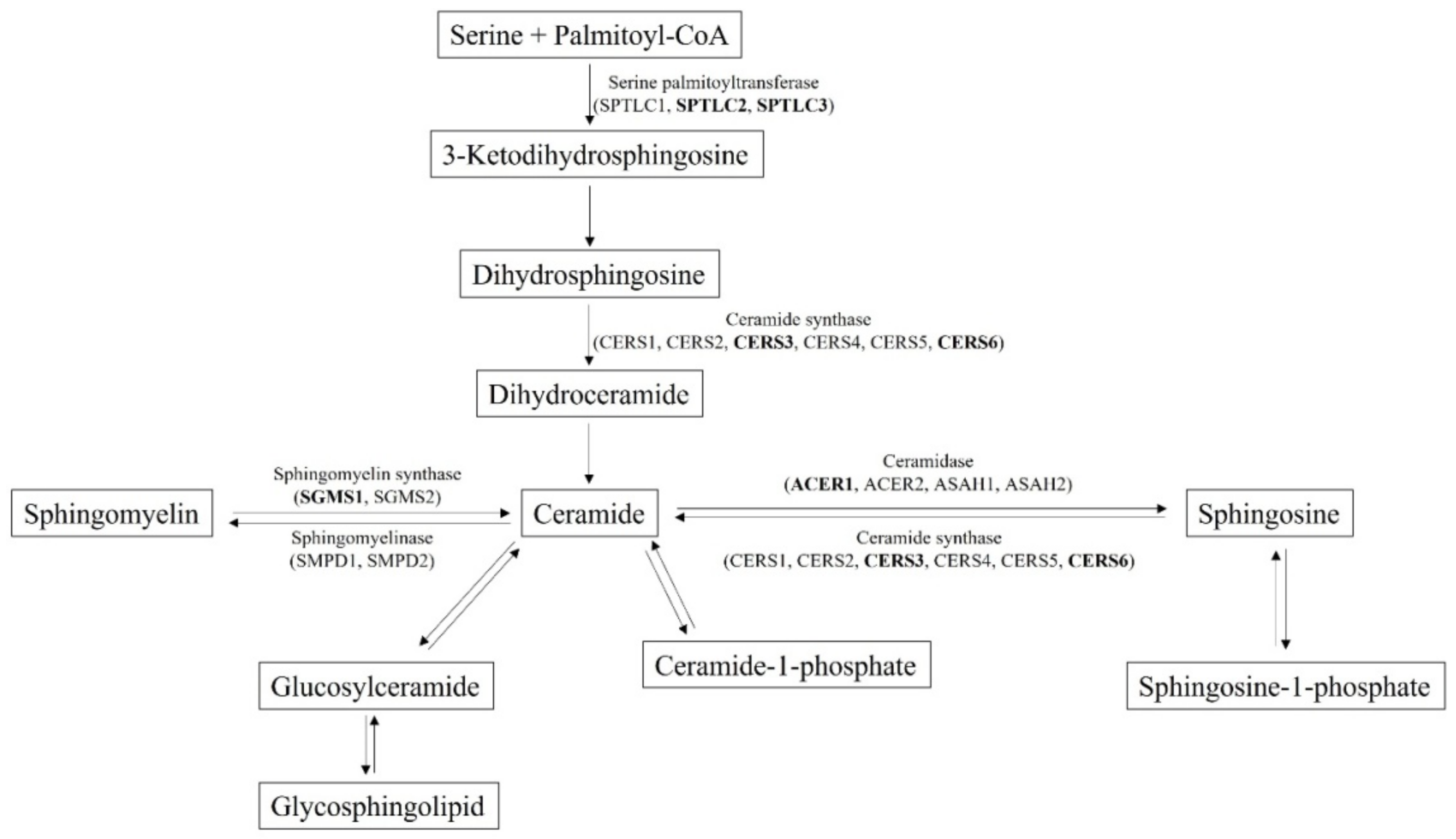
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Their Classification
2.2. Selection of Ceramides and Dihydroceramides
2.3. Sample Preparation for LC-MS/MS Analysis
2.4. LC-MS/MS Analysis of Plasma Ceramide Species
2.5. SNP Genotyping
2.6. Association Analysis
3. Results
3.1. Clinical Characteristics of Study Participants
3.2. LC-MS/MS Measurement of Plasma Level of Ceramides
3.3. Identification of SNPs Associated with Ceramide Biosynthesis Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [PubMed] [Green Version]
- Lim, S.; Jang, H.C.; Lee, H.K.; Kim, K.C.; Cho, N.H. A rural-urban comparison of the characteristics of the metabolic syndrome by gender in Korea: The Korean Health and Genome Study (KHGS). J. Endocrinol. Investig. 2006, 29, 313–319. [Google Scholar]
- Rakugi, H.; Ogihara, T. The metabolic syndrome in the Asian population. Curr. Hypertens. Rep. 2005, 7, 103–109. [Google Scholar]
- Moore, J.X.; Chaudhary, N.; Akinyemiju, T. Metabolic syndrome prevalence by race/ethnicity and sex in the United States, national health and nutrition examination survey, 1988–2012. Prev. Chronic Dis. 2017, 14, E24. [Google Scholar] [PubMed] [Green Version]
- Williams, V.F.; Oh, G.T.; Stahlman, S. Incidence and prevalence of the metabolic syndrome using ICD-9 and ICD-10 diagnostic codes, active component, U.S. Armed Forces, 2002–2017. MSMR 2018, 25, 20–25. [Google Scholar] [PubMed]
- Ahmadian, M.; Duncan, R.E.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Triacylglycerol metabolism in adipose tissue. Future Lipidol. 2007, 2, 229–237. [Google Scholar]
- Bonen, A.; Dohm, G.L.; van Loon, L.J. Lipid metabolism, exercise and insulin action. Essays Biochem. 2006, 42, 47–59. [Google Scholar]
- Hannun, Y.A. Functions of ceramide in coordinating cellular responses to stress. Science 1996, 274, 1855–1859. [Google Scholar]
- Hannum, Y.A.; Obeid, L.M. Principles of bioactive lipid signaling: Lessons from sphingolipids. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar]
- McGurk, K.A.; Keavney, B.D.; Nicolaoou, A. Circulating ceramides as biomarkers of cardiovascular disease: Evidence from phenotypic and genomic studies. Atherosclerosis 2021, 327, 18–30. [Google Scholar]
- Turpin-Nolan, S.M.; Bruning, J.C. The role of ceramides in metabolic disorders: When size and localization matters. Nat. Rev. Endocrinol. 2020, 16, 224–233. [Google Scholar]
- Consitt, L.A.; Bell, J.A.; Houmard, J.A. Intramuscular lipid metabolism, insulin action, and obesity. IUBMB Life 2009, 61, 47–55. [Google Scholar] [PubMed] [Green Version]
- Oh, S.-H.; Lee, J.E.; Shin, E.; Kwon, H.; Choe, E.K.; Choi, S.Y.; Rhee, H.; Choi, S.H. Genome-wide association study of metabolic syndrome in Korean populations. PLoS ONE 2020, 15, e0227357. [Google Scholar]
- Brown, A.E.; Walker, M. Genetics of insulin resistance and the metabolic syndrome. Curr. Cardiol. Rep. 2016, 18, 75. [Google Scholar] [PubMed] [Green Version]
- Kleinendorst, L.; Massink, M.P.G.; Cooiman, M.I.; Savas, M.; van der Baan-Slootweg, O.H.; Roelants, R.J.; Janssen, I.C.M.; Meijers-Heijboer, H.J.; Knoers, N.V.A.M.; van Amstel, H.K.P.; et al. Genetic obesity: Next-generation sequencing results of 1230 patients with obesity. J. Med. Genet. 2018, 55, 578–586. [Google Scholar]
- Foucan, L.; Larifla, L.; Durand, E.; Rambhojan, C.; Armand, C.; Michel, C.T.; Billy, R.; Dhennin, V.; De Graeve, F.; Rabearivelo, I.; et al. High prevalence of rare monogenic forms of obesity in obese Guadeloupean Afro-Caribbean children. J. Clin. Endocrinol. Metab. 2018, 103, 539–545. [Google Scholar]
- Sung, J.; Lee, K.; Song, Y.M. Heritabilities of the metabolic syndrome phenotypes and related factors in Korean twins. J. Clin. Endocrinol. Metab. 2009, 94, 4946–4952. [Google Scholar]
- Hicks, A.A.; Pramstaller, P.P.; Johansson, A.; Vitart, V.; Rudan, I.; Ugocsai, P.; Aulchenko, Y.; Franklin, C.S.; Liebisch, G.; Erdmann, J.; et al. Genetic determinants of circulating sphingolipid concentrations in European populations. PLoS Genet. 2009, 5, e1000672. [Google Scholar]
- Moon, S.; Kim, Y.J.; Han, S.; Hwang, M.Y.; Shin, D.M.; Park, M.Y.; Lu, Y.; Yoon, K.; Jang, H.M.; Kim, Y.K.; et al. The Korea Biobank Array: Design and identification of coding variants associated with blood biochemical traits. Sci. Rep. 2019, 9, 1382. [Google Scholar]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C., Jr.; et al. Harmonizing the Metabolic Syndrome. Circulation 2009, 120, 1640–1645. [Google Scholar]
- Kang, H.T.; Kim, S.Y.; Kim, J.; Kim, J.; Kim, J.; Park, H.A.; Shin, J.; Cho, S.H.; Choi, Y.; Shim, J.Y. Clinical practice guideline of prevention and treatment for metabolic syndrome. Korean J. Fam. Pract. 2015, 5, 375–420. [Google Scholar]
- Basit, A.; Piomelli, D.; Armirotti, A. Rapid evaluation of 25 key sphingolipids and phosphosphingolipids in human plasma by LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 5189–5198. [Google Scholar] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [PubMed] [Green Version]
- Mao, C.; Obeid, L.M. Ceramidases: Regulators of cellular responses mediated by ceramide, sphingosine, and sphingosine- 1-phosphate. Biochim. Biophy. Acta 2008, 1781, 424–434. [Google Scholar]
- Bilal, F.; Montfort, A.; Gilhodes, J.; Garcia, V.; Riond, J.; Carpentier, S.; Filleron, T.; Colacios, C.; Levade, T.; Daher, A.; et al. Sphingomyelin synthase 1 (SMS1) downregulation is associated with sphingolipid reprogramming and a worse prognosis in melanoma. Front. Pharmacol. 2019, 30, 443. [Google Scholar]
- Choromañska, B.; Myśliwiec, P.; Hady, H.R.; Dadan, J.; Myśliwiec, H.; Chabowski, A.; Mikłosz, A. Metabolic syndrome is associated with ceramide accumulation in visceral adipose tissue of women with morbid obesity. Obesity 2019, 27, 444–453. [Google Scholar]
- Yuyama, K.; Mitsutake, S.; Igarashi, Y. Pathological roles of ceramide and its metabolites in metabolic syndrome and Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1841, 793–798. [Google Scholar]
- Sen, P.; Hyötyläinen, T.; Orešic, M. 1-Deoxyceramides—Key players in lipotoxicity and progression to type 2 diabetes? Acta Physiol. 2021, 232, e13635. [Google Scholar]
- Kolter, T.; Sandhoff, K. Sphingolipid metabolism diseases. Biochim. Biophys. Acta 2006, 1758, 2057–2079. [Google Scholar]
- Morad, S.A.; Cabot, M.C. Ceramide-orchestrated signalling in cancer cells. Nat. Rev. Cancer 2013, 13, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.M.; Bandaru, V.V.R.; Haughey, N.J.; Xia, J.; Fried, L.P.; Yasar, S.; Albert, M.; Varma, V.; Harris, G.; Schneider, E.B.; et al. Serum ceramides increase the risk of Alzheimer disease: The Women’s Health and Aging Study II. Neurology 2012, 79, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, M.M.; Haughey, N.J.; Bandaru, V.V.R.; Schech, S.; Carrick, R.; Carlson, M.C.; Mori, S.; Miller, M.I.; Ceritoglu, C.; Brown, T.; et al. Plasma ceramides are altered in mild cognitive impairment and predict cognitive decline and hippocampal volume loss. Alzheimers Dement. 2010, 6, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, C.; Lin, X.; Sun, H.; Xu, R.; Li, Q.; Wei, Y.; Li, Y.; Qian, J.; Liu, C.; et al. Targeting alkaline ceramidase 3 alleviates the severity of nonalcoholic steatohepatitis by reducing oxidative stress. Cell Death Dis. 2020, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Fan, Y.; Liu, J.; Li, Y.; Quan, C.; Bui, H.H.; Kuo, M.S.; Park, T.S.; Cao, G.; Jiang, X.C. The impact of sphingomyelin synthase 1 deficiency on sphingolipid metabolism and atherosclerosis in mice. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, M.J.; Barnett, A.C.; Bruce, C.R.; Schenk, S.; Horowitz, J.F.; Hoy, A.J. Regulation of plasma ceramide levels with fatty acid oversupply: Evidence that the liver detects and secretes de novo synthesized ceramide. Diabetologia 2012, 55, 2741–2746. [Google Scholar] [CrossRef] [Green Version]
- Tidhar, R.; Futerman, A.H. The complexity of sphingolipid biosynthesis in the endoplasmic reticulum. Biochim. Biophy. Acta 2013, 1833, 2511–2518. [Google Scholar] [CrossRef] [Green Version]
- Hornemann, T.; Penno, A.; Rütti, M.F.; Ernst, D.; Kivrak-Pfiffner, F.; Rohrer, L.; von Eckardstein, A. The SPTLC3 subunit of serine palmitoyltransferase generates short chain sphingoid bases. J. Biol. Chem. 2009, 284, 26322–26330. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Control | MetS | p Value | |
|---|---|---|---|
| Number of study participants | 38 | 37 | |
| Age (years) | 38.9 ± 11.7 | 46.6 ± 10.3 | 3.5 × 10−3 |
| Height (cm) | 165.1 ± 8.1 | 170.3 ± 8.9 | 9.9 × 10−3 |
| Weight (kg) | 58.1 ± 9.7 | 82.5 ± 16.1 | <1.0 × 10−4 |
| Waist measurement (cm) | 75.2 ± 6.9 | 94.6 ± 9.7 | <1.0 × 10−4 |
| BMI (kg/m3) | 21.2 ± 2.1 | 27.9 ± 3.4 | <1.0 × 10−4 |
| SBP (mmHg) | 112.5 ± 9.1 | 126.2 ± 9.7 | <1.0 × 10−4 |
| DBP (mmHg) | 67.1 ± 6.5 | 76.7 ± 8.1 | <1.0 × 10−4 |
| Blood glucose (mg/dL) | 91.0 ± 5.4 | 106.5 ± 13.0 | <1.0 × 10−4 |
| Total cholesterol (mg/dL) | 189.5 ± 30.2 | 202.3 ± 39.5 | 0.1 |
| HDL-C (mg/dL) | 71.4 ± 12.6 | 48.4 ± 12.8 | <1.0 × 10−4 |
| LDL-C (mg/dL) | 102.9 ± 31.5 | 116.8 ± 34.3 | 6.8 × 10−2 |
| TG (mg/dL) | 81.3 ± 29.0 | 205.3 ± 102.5 | <1.0 × 10−4 |
| AST (U/L) | 20.2 ± 5.0 | 31.3 ± 10.2 | <1.0 × 10−4 |
| ALT (U/L) | 16.3 ± 7.1 | 42.1 ± 21.9 | <1.0 × 10−4 |
| γ-GTP (U/L) | 17.5 ± 8.5 | 59.8 ± 64.2 | <1.0 × 10−4 |
| Serum creatinine (mg/dL) | 0.8 ± 0.2 | 0.9 ± 0.2 | 4.0 × 10−4 |
| GFR (mL/min/1.73 m2) | 101.1 ± 21.7 | 92.1 ± 13.7 | 3.5 × 10−2 |
| Leptin (pg/mL) | 5089.8 ± 4803.1 | 7866.6 ± 7743.7 | 6.4 × 10−2 |
| Adiponectin (ng/mL) | 6464.2 ± 3806.9 | 2537.2 ± 1633.2 | <1.0 × 10−4 |
| hs-CRP (mg/L) | 0.9 ± 1.3 | 1.5 ± 1.2 | 4.0 × 10−2 |
| Insulin (µIU/mL) | 4.7 ± 2.6 | 12.4 ± 6.5 | <1.0 × 10−4 |
| HOMA-IR | 1.1 ± 0.6 | 3.3 ± 1.9 | <1.0 × 10−4 |
| Control (ng/mL) | MetS (ng/mL) | p Value | |
|---|---|---|---|
| Cer-16 | 61.8 ± 12.7 | 80.0 ± 17.6 | <1.0 × 10−4 |
| Cer-18 | 34.9 ± 12.9 | 65.2 ± 25.4 | <1.0 × 10−4 |
| Cer-20 | 45.3 ± 14.1 | 79.1 ± 24.9 | <1.0 × 10−4 |
| DhCer-16 | 7.6 ± 3.6 | 8.8 ± 3.4 | 0.13 |
| DhCer-18 | 9.9 ± 8.8 | 18.4 ± 12.4 | 9.0 × 10−4 |
| DhCer-24 | 122.1 ± 70.7 | 163.8 ± 60.6 | 7.2 × 10−3 |
| DhCer24:1 | 99.5 ± 54.0 | 136.9 ± 62.4 | 6.6 × 10−3 |
| Gene | dbSNP ID | Chr | Position | Ref † allele | Alt ǂ allele | Associated Ceramide | FDR-Adjusted p Value |
|---|---|---|---|---|---|---|---|
| CERS6 | rs75397325 | 2 | 169607910 | G | C | Cer-16 Cer-20 | 5.2 × 10−3 2.1 × 10−2 |
| CERS3 | rs72759132 | 15 | 101023194 | G | A | Cer-16 | 5.0 × 10−2 |
| CERS3 | rs4246316 | 15 | 101066561 | C | T | Cer-16 | 3.1 × 10−2 |
| CERS6 | rs80165332 | 2 | 169516096 | C | T | Cer-16 | 2.8 × 10−2 |
| ACER1 | rs62106618 | 19 | 6314542 | A | G | DhCer-18 DhCer-24 DhCer-24:1 | 3.4 × 10−2 4.7 × 10−2 2.6 × 10−2 |
| SGMS1 | rs12358192 | 10 | 52364839 | T | C | Cer-16 Cer-18 Cer-20 | 2.6 × 10−3 2.3 × 10−2 1.5 × 10−2 |
| SGMS1 | rs11006229 | 10 | 52350006 | T | C | DhCer-24 Cer-16 Cer-18 Cer-20 | 3.9 × 10−2 7.8 × 10−3 1.8 × 10−2 1.0 × 10−2 |
| SGMS1 | rs10826014 | 10 | 52209580 | T | C | Cer-18 | 4.2 × 10−2 |
| SPTLC2 | rs149162405 | 14 | 78076411 | T | C | Cer-20 | 3.6 × 10−2 |
| SPTLC3 | rs6109681 | 20 | 13047504 | T | C | Cer-16 | 4.4 × 10−2 |
| SPTLC3 | rs3906631 | 20 | 13064936 | C | A | Cer-16 | 1.3 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Kim, S.-A.; Kim, Y.; Kim, J.; Hong, G.; Hong, J.; Choi, K.; Eom, C.-S.; Baik, S.; Lee, M.-K.; et al. Genetic Variants Associated with Elevated Plasma Ceramides in Individuals with Metabolic Syndrome. Genes 2022, 13, 1497. https://doi.org/10.3390/genes13081497
Lee S, Kim S-A, Kim Y, Kim J, Hong G, Hong J, Choi K, Eom C-S, Baik S, Lee M-K, et al. Genetic Variants Associated with Elevated Plasma Ceramides in Individuals with Metabolic Syndrome. Genes. 2022; 13(8):1497. https://doi.org/10.3390/genes13081497
Chicago/Turabian StyleLee, Sanghoo, Seol-A Kim, Yejin Kim, Juhoon Kim, Gayeon Hong, Jeonghoon Hong, Kyeonghwan Choi, Chun-Sick Eom, Saeyun Baik, Mi-Kyeong Lee, and et al. 2022. "Genetic Variants Associated with Elevated Plasma Ceramides in Individuals with Metabolic Syndrome" Genes 13, no. 8: 1497. https://doi.org/10.3390/genes13081497
APA StyleLee, S., Kim, S.-A., Kim, Y., Kim, J., Hong, G., Hong, J., Choi, K., Eom, C.-S., Baik, S., Lee, M.-K., & Lee, K.-R. (2022). Genetic Variants Associated with Elevated Plasma Ceramides in Individuals with Metabolic Syndrome. Genes, 13(8), 1497. https://doi.org/10.3390/genes13081497