Author Contributions
Conceptualization, S.K.D.; design of the experiment, S.M, S.K.D. and M.H.A.; methodology, S.M., S.K.D. and M.H.A.; data analysis, S.K.D., M.A.S., A.G. and A.H.; data curation, S.K.D. and M.A.S., writing—original draft preparation, S.K.D., M.A.S., and P.R.R.; critical review and editing, S.K.D., M.A.S., A.G., M.B. and A.H.; funding, M.B. and A.H. All authors have read and agreed to the published version of the manuscript.
Figure 1.
Correlation coefficient matrix, scatter plot, and phenotypic frequency distribution among grain yield and yield-related traits over the locations. Each variable’s distribution is displayed diagonally. The bivariate scatter plots with a trend line are shown at the bottom of the diagonal. The correlation coefficient and the level of significance are displayed as stars at the top of the diagonal. * p ≤ 0.05, ** p ≤ 0.01, and *** p > 0.001 show significance level; Note: X50F = days to 50% flowering, PH = plant height (cm), TILL = number of tillers per hill, PPH = number of panicles per hill, PL = panicle length (cm), GP = grains per panicle, Fertility = fertility (%), YPP = grain yield per plant (gm), TGW = thousand grain weight (gm), and YTH = grain yield (t/ha).
Figure 1.
Correlation coefficient matrix, scatter plot, and phenotypic frequency distribution among grain yield and yield-related traits over the locations. Each variable’s distribution is displayed diagonally. The bivariate scatter plots with a trend line are shown at the bottom of the diagonal. The correlation coefficient and the level of significance are displayed as stars at the top of the diagonal. * p ≤ 0.05, ** p ≤ 0.01, and *** p > 0.001 show significance level; Note: X50F = days to 50% flowering, PH = plant height (cm), TILL = number of tillers per hill, PPH = number of panicles per hill, PL = panicle length (cm), GP = grains per panicle, Fertility = fertility (%), YPP = grain yield per plant (gm), TGW = thousand grain weight (gm), and YTH = grain yield (t/ha).
Figure 2.
Scree plot on variability explained by each component of 34 elite rice genotypes.
Figure 2.
Scree plot on variability explained by each component of 34 elite rice genotypes.
Figure 3.
Biplot for the 34 rice genotypes and 10 agronomic traits along the first 2 principal components. Note: X50F = days to 50% flowering, PH = plant height (cm), TILL = number of tillers per hill, PPH = number of panicles per hill, PL = panicle length (cm), GP = grains per panicle, Fertility = fertility (%), YPP = grain yield per plant (gm), TGW = thousand grain weight (gm), and YTH = grain yield (t/ha).
Figure 3.
Biplot for the 34 rice genotypes and 10 agronomic traits along the first 2 principal components. Note: X50F = days to 50% flowering, PH = plant height (cm), TILL = number of tillers per hill, PPH = number of panicles per hill, PL = panicle length (cm), GP = grains per panicle, Fertility = fertility (%), YPP = grain yield per plant (gm), TGW = thousand grain weight (gm), and YTH = grain yield (t/ha).
Figure 4.
Graphical representation on (A) percent of contribution and (B) quality of representation of top 5 variables to principal components 1-2-3-4-5.
Figure 4.
Graphical representation on (A) percent of contribution and (B) quality of representation of top 5 variables to principal components 1-2-3-4-5.
Figure 5.
Graphical representation of (A) percent of contribution and (B) quality of representation of top 10 genotypes to principal components 1-2-3-4-5.
Figure 5.
Graphical representation of (A) percent of contribution and (B) quality of representation of top 10 genotypes to principal components 1-2-3-4-5.
Figure 6.
Graphical method (A) Hubert index and (B) Dindex for detecting the optimum number of clusters.
Figure 6.
Graphical method (A) Hubert index and (B) Dindex for detecting the optimum number of clusters.
Figure 7.
The heatmap displays the clustering pattern of 34 rice genotypes with 10 quantitative characters. Each column indicates a character, whereas each row represents a genotype. The various colours and intensities (−2 to 3) were adjusted based on the genotype–characters relationship. The orange colour represents a lower value, the white colour for mid value, and the dark red indicates a higher value.
Figure 7.
The heatmap displays the clustering pattern of 34 rice genotypes with 10 quantitative characters. Each column indicates a character, whereas each row represents a genotype. The various colours and intensities (−2 to 3) were adjusted based on the genotype–characters relationship. The orange colour represents a lower value, the white colour for mid value, and the dark red indicates a higher value.
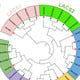
Figure 8.
The circular preview indicates the ranking of genotypes based on the MGIDI selection index as well as the best rice genotypes with the associated locations (A) Satkhira, (B) Kushtia, and (C) Barishal. The selected ones are marked in red colour.
Figure 8.
The circular preview indicates the ranking of genotypes based on the MGIDI selection index as well as the best rice genotypes with the associated locations (A) Satkhira, (B) Kushtia, and (C) Barishal. The selected ones are marked in red colour.
Figure 9.
The strengths and weaknesses view of the selected genotypes represent the proportion of each factor on the computed MGIDI index. (A) Satkhira, (B) Kushtia, and (C) Barishal.
Figure 9.
The strengths and weaknesses view of the selected genotypes represent the proportion of each factor on the computed MGIDI index. (A) Satkhira, (B) Kushtia, and (C) Barishal.
Table 1.
Monthly mean agro-climatic data during the field experiment period in Barishal, Kushtia, and Satkhira.
Table 1.
Monthly mean agro-climatic data during the field experiment period in Barishal, Kushtia, and Satkhira.
| Locations | Month | Maxi. (°C) | Min. (°C) | Wind (km/h) | Rainfall (mm) | Humidity (%) | Cloud (%) | Pressure (mb) |
|---|
| Barishal | February | 27.0 | 17.7 | 10.5 | 0 | 43.7 | 0 | 1014.5 |
| March | 31.7 | 21.5 | 9.5 | 0 | 54.0 | 9.7 | 1013.5 |
| April | 31.5 | 22.7 | 11.7 | 1.0 | 61.0 | 13.7 | 1013.2 |
| May | 35.2 | 26.7 | 13.7 | 2.4 | 67.7 | 37.5 | 1007.7 |
| Kushtia | February | 28.0 | 16.5 | 10.0 | 0 | 39.5 | 0 | 1014.7 |
| March | 33.0 | 20.5 | 9.5 | 0 | 38.0 | 3.7 | 1011.7 |
| April | 35.2 | 21.7 | 9.2 | 1.2 | 42.5 | 5.2 | 1009.7 |
| May | 38.5 | 26.7 | 12.2 | 7.0 | 55.5 | 33.0 | 1005.5 |
| Satkhira | February | 27.5 | 17.0 | 12.2 | 0 | 38.5 | 0 | 1015.5 |
| March | 33.7 | 20.7 | 8.5 | 0.0 | 43.7 | 8.7 | 1013.2 |
| April | 34.5 | 22.5 | 10.2 | 0.3 | 50.2 | 4.7 | 1010.7 |
| May | 37.2 | 26.2 | 15.2 | 0.1 | 63.5 | 36.2 | 1007.0 |
Table 2.
The designation and parentages of 34 elite rice cultivars utilized in the current investigation.
Table 2.
The designation and parentages of 34 elite rice cultivars utilized in the current investigation.
| Code | Designation | Parentage |
|---|
| Gen1 | BR7671-37-2-2-37-3-P3 | BRRI dhan29/IR68144 |
| Gen2 | BR8626-19-5-1-2 | BR7166-5B-6/SHEW WAR TUN//BRRI dhan47 |
| Gen3 | BR8626-10-5-1 | BR7166-5B-6/SHEW WAR TUN//BRRI dhan47 |
| Gen4 | BR8109-29-2-2-3 | BM9821/Parija |
| Gen5 | BR(Bio)8333-BC5-1-1 | BRRI dhan29/IRBB60 |
| Gen6 | BR(Bio)8333-BC5-1-20 | BRRI dhan29/IRBB60 |
| Gen7 | BR(Bio)8333-BC5-2-16 | BRRI dhan29/IRBB60 |
| Gen8 | BR(Bio)8333-BC5-2-22 | BRRI dhan29/IRBB60 |
| Gen9 | BR(Bio)8333-BC5-3-10 | BRRI dhan29/IRBB60 |
| Gen10 | BR8631-12-3-5-P2 | BR7166-5B-5/BG305//BRRI dhan29 |
| Gen11 | BR8631-12-3-6-P3 | BR7166-5B-5/BG305//BRRI dhan29 |
| Gen12 | BR7831-59-1-1-4-5 | BR7166-5B-5/BG305//BRRI dhan29 |
| Gen13 | BR8253-9-3-3-1 | BR7305-21-6-1/BRRI dhan29//BR7305-21-6-1 |
| Gen14 | BR8609-2-B-9-1-B5 | BRRIdhan29/BR7166-5B-1 |
| Gen15 | BR7815-18-1-3-2-1 | BRRI dhan36/BR6725-15-1-2 |
| Gen16 | BR7671-37-2-2-3-7-3-P10 | BRRI dhan29/IR68144 |
| Gen17 | BR7671-37-2-2-3-7-3-P11 | BRRI dhan29/IR68144 |
| Gen18 | BR8079-19-1-5-1 | IR71730-51-2/Nayanmoni |
| Gen19 | BR8590-5-2-5-2-2 | BR6902-51-3-2-1/BRRI dhan HR7 |
| Gen20 | BR8590-5-3-3-4-2 | BR6902-51-3-2-1/BRRI dhan HR7 |
| Gen21 | BR8608-39-2-1 | BRRI dhan 29/Shaheen Basmati/BR6902-51-3-2-1 |
| Gen22 | BRC266-5-1-1-1 | BR16/90060-TR1252-8-2-1 |
| Gen23 | BRC266-5-1-2-1 | BR16/90060-TR1252-8-2-1 |
| Gen24 | BR8523-36-2-2-6 | IR 77512-128-2-1-2/BR 6817-25-2-2 |
| Gen25 | BR8938-19-4-3-1-1 | IRBB60/BRRI dhan29 |
| Gen26 | BR8333-15-3-2-2 | BRRI dhan29/IRBB60 |
| Gen27 | IRBB60 | Near isogenic line of IR24 |
| Gen28 | BRRI dhan28 | BR6(IR28)/Purbachi |
| Gen29 | BRRI dhan58 | Somaclonal line of BRRI dhan29 |
| Gen30 | BRRI dhan29 | BG90-2/BR51-46-5 |
| Gen31 | BR16 | IR1416-131-5/IR1364-37-3-1//IR1544A-E666 |
| Gen32 | BRRI dhan50 | BR30/IR67684B |
| Gen33 | BRRI dhan63 | Amol-3/BRRI dhan28 |
| Gen34 | BRRI dhan74 | BRRI dhan29/IR68144 |
Table 3.
Combined analysis of variance, mean sum of square (MSS) and F-value of the ten traits, and their interaction in elite rice breeding lines in studied locations.
Table 3.
Combined analysis of variance, mean sum of square (MSS) and F-value of the ten traits, and their interaction in elite rice breeding lines in studied locations.
| Traits a | Barishal | Kushtia | Satkhira | Pooled b | Interaction c |
|---|
| MSS | F-Value | MSS | F-Value | MSS | F-Value | MSS | F-Value | MSS | F-Value |
|---|
| X50F | 196.2 | 160.3 ** | 66.9 | 62.7 ** | 51.7 | 49.2 ** | 224.2 | 203.9 ** | 44.7 | 40.7 ** |
| PH | 391.3 | 23.1 ** | 225.8 | 21.2 ** | 258.2 | 26.8 ** | 783.4 | 63.1 ** | 45.9 | 3.7 ** |
| TILL | 18.7 | 5.9 ** | 31.4 | 4.3 ** | 10.2 | 11.2 ** | 36.3 | 9.5 ** | 12 | 3.2 ** |
| PPH | 21.8 | 7.3 ** | 8.4 | 2.6 ** | 12.3 | 8.4 ** | 16.3 | 6.4 ** | 13 | 5.2 ** |
| PL | 4.8 | 5.7 ** | 8.8 | 3.7 ** | 5.4 | 6.1 ** | 11.3 | 8.2 ** | 3.8 | 2.8 ** |
| GP | 27,549.9 | 7.5 ** | 9620.6 | 11.6 ** | 7675.9 | 7.9 ** | 31,784.3 | 17.4 ** | 6531 | 3.6 ** |
| Ferti | 258.1 | 14.1 ** | 161.6 | 6.4 ** | 124.7 | 5.4 ** | 171.3 | 7.7 ** | 186.8 | 8.4 ** |
| YPP | 27.2 | 12.2 ** | 43.7 | 7.9 ** | 16.2 | 10.4 ** | 48.9 | 15.8 ** | 19 | 6.2 ** |
| TGW | 37.2 | 19.2 ** | 27.3 | 36.8 ** | 20.7 | 6.3 ** | 72.7 | 36.5 ** | 6.2 | 3.2 ** |
| YTH | 1.9 | 10.5 ** | 2.9 | 9.3 ** | 1.8 | 16.8 ** | 4.8 | 22.6 ** | 1 | 5.2 ** |
Table 4.
Estimation of variance components, broad sense heritability, and genetic advance in elite rice genotypes.
Table 4.
Estimation of variance components, broad sense heritability, and genetic advance in elite rice genotypes.
| Traits | Location | X50F | PH | TILL | PPH | PL | GP | Ferti | YPP | TGW | YTH |
|---|
| Range | Barishal | 103–135 | 80–148 | 8–23 | 8–23 | 20–27.9 | 151–607 | 51–87.3 | 16.9–32.5 | 15.4–33 | 4.5–8.6 |
| Kushtia | 110–129 | 77–131 | 8–29 | 5–18 | 19–30 | 160–484 | 57.4–94.9 | 18–37.5 | 16.9–31.2 | 4.4–9.3 |
| Satkhira | 106–124 | 81–124 | 7–18 | 6.3–16.5 | 18.5–29.8 | 79–375 | 54.3–94.5 | 17.6–28.9 | 13.8–28.9 | 4.6–7.9 |
| Mean ± SE | Barishal | 118.2 ± 0.9 | 105.7 ± 3.4 | 13.8 ± 1.5 | 12.5 ± 1.4 | 23.9 ± 0.7 | 307.3 ± 49.6 | 66.1 ± 3.4 | 24.2 ± 1.2 | 21.8 ± 1.2 | 6.41 ± 0.4 |
| Kushtia | 121.0 ± 0.8 | 99.0 ± 2.67 | 15.7 ± 2.2 | 10.4 ± 1.5 | 24.4 ± 1.3 | 267.5 ± 23.5 | 79.7 ± 4.2 | 28.6 ± 1.9 | 22.1 ± 0.7 | 7.04 ± 0.5 |
| Satkhira | 114.7 ± 0.8 | 101.2 ± 2.5 | 11.2 ± 0.7 | 10.7 ± 0.9 | 23.8 ± 0.7 | 210 ± 25.5 | 80.2 ± 3.9 | 22.7 ± 1.1 | 20.7 ± 1.5 | 6.24 ± 0.2 |
| ẟ2g | Barishal | 64.9 | 124.7 | 5.2 | 6.3 | 1.3 | 7951.9 | 88.9 | 8.30 | 11.7 | 0.6 |
| Kushtia | 21.9 | 71.7 | 8.1 | 1.7 | 2.2 | 2929.7 | 45.4 | 12.7 | 8.8 | 0.8 |
| Satkhira | 16.8 | 82.8 | 3.1 | 3.6 | 1.5 | 2234.5 | 33.9 | 4.8 | 5.8 | 0.5 |
| ẟ2p | Barishal | 65.4 | 130.4 | 6.2 | 7.3 | 1.6 | 9183.3 | 95.0 | 9.1 | 12.4 | 0.6 |
| Kushtia | 22.3 | 75.3 | 10.5 | 2.8 | 2.9 | 3206.8 | 53.8 | 14.6 | 9.1 | 0.9 |
| Satkhira | 17.3 | 89.0 | 3.4 | 4.1 | 1.8 | 2558.6 | 41.5 | 5.4 | 6.9 | 0.6 |
| ẟ2e | Barishal | 1.3 | 16.9 | 3.2 | 3.0 | 0.8 | 3694.2 | 18.3 | 2.3 | 1.9 | 0.2 |
| Kushtia | 1.1 | 10.7 | 7.4 | 3.2 | 2.4 | 831.5 | 25.5 | 5.5 | 0.7 | 0.3 |
| Satkhira | 1.1 | 9.6 | 0.9 | 1.5 | 0.8 | 972.4 | 22.9 | 1.5 | 3.3 | 0.1 |
| GCV (%) | Barishal | 54.9 | 117.9 | 37.3 | 50.3 | 5.6 | 2588.0 | 134.5 | 34.4 | 53.6 | 9.3 |
| Kushtia | 3.8 | 8.5 | 17.9 | 12.7 | 5.9 | 20.3 | 8.5 | 12.5 | 13.4 | 13.2 |
| Satkhira | 3.5 | 8.9 | 15.7 | 17.6 | 5.2 | 22.5 | 7.3 | 9.7 | 11.6 | 11.9 |
| PCV (%) | Barishal | 6.8 | 10.5 | 16.5 | 20.0 | 4.8 | 29.0 | 13.5 | 11.9 | 15.6 | 12.0 |
| Kushtia | 3.96 | 9.2 | 24.8 | 8.7 | 22.9 | 10.5 | 14.9 | 13.9 | 15.4 | 8.7 |
| Satkhira | 3.6 | 9.5 | 17.8 | 6.4 | 26.9 | 9.4 | 11.1 | 14.5 | 13.0 | 6.4 |
| ECV (%) | Barishal | 6.8 | 11.3 | 20.9 | 6.1 | 35.1 | 14.9 | 13.4 | 16.9 | 13.7 | 6.1 |
| Kushtia | 0.8 | 3.3 | 17.2 | 17.1 | 6.3 | 10.7 | 6.3 | 8.1 | 3.8 | 7.9 |
| Satkhira | 0.8 | 3.0 | 8.5 | 11.2 | 3.9 | 14.8 | 5.9 | 5.4 | 8.7 | 5.2 |
| RD (%) | Barishal | 0.9 | 3.9 | 13.0 | 13.8 | 3.8 | 19.7 | 6.4 | 6.1 | 6.3 | 6.7 |
| Kushtia | 1.6 | 4.7 | 23.3 | 37.5 | 27.2 | 8.6 | 15.7 | 12.5 | 2.7 | 10.7 |
| Satkhira | 2.2 | 6.9 | 8.8 | 11.9 | 16.4 | 12.7 | 18.4 | 9.5 | 16.7 | 5.9 |
| h2b (%) | Barishal | 99.4 | 95.6 | 83.2 | 86.2 | 82.6 | 86.6 | 93.5 | 91.7 | 94.8 | 90.5 |
| Kushtia | 98.4 | 95.3 | 76.7 | 62.5 | 72.8 | 91.4 | 84.2 | 87.4 | 97.3 | 89.2 |
| Satkhira | 97.9 | 93.0 | 91.2 | 88.0 | 83.6 | 87.3 | 81.6 | 90.5 | 84.1 | 94.0 |
| GA (%) | Barishal | 14.0 | 21.3 | 30.7 | 38.4 | 9.1 | 55.6 | 28.4 | 23.5 | 31.4 | 23.5 |
| Kushtia | 7.9 | 17.2 | 32.5 | 20.7 | 10.5 | 39.8 | 15.9 | 24.1 | 27.3 | 25.7 |
| Satkhira | 7.3 | 17.8 | 30.9 | 34.0 | 9.7 | 43.3 | 13.5 | 19.0 | 21.9 | 23.9 |
Table 5.
List of the best number of clusters and their index value of 30 indices implemented in the NbClust package in R.
Table 5.
List of the best number of clusters and their index value of 30 indices implemented in the NbClust package in R.
| | KL | CH | Hartigan | CCC | Scott | Marriot |
| No. of clusters | 8 | 3 | 3 | 10 | 3 | 3 |
| Index value | 2.1197 | 7.8206 | 3.051 | −0.7325 | 87.9885 | 2.49 × 1012 |
| | TrCovW | TraceW | Friedman | Rubin | Cindex | DB |
| No. of clusters | 3 | 3 | 8 | 8 | 8 | 10 |
| Index value | 310.342 | 23.6767 | 61.487 | −0.1678 | 0.4 | 1.0447 |
| | Silhouette | Duda | PseudoT2 | Beale | Ratkowsky | Ball |
| No. of clusters | 10 | 2 | 2 | 2 | 4 | 3 |
| Index value | 0.2796 | 1.0678 | −1.6513 | −0.4058 | 0.3152 | 61.7704 |
| | PtBiserial | Gap | McClain | γ | Gplus | Tau |
| No. of clusters | 5 | 2 | 2 | 10 | 10 | 5 |
| Index value | 0.5448 | −0.7636 | 0.8023 | 0.8943 | 2.8324 | 79.1515 |
| | Frey | Hubert | Dindex | Dunn | SDindex | SDbw |
| No. of clusters | 6 | 9 | 3 | 10 | 7 | 10 |
| Index value | 0.6385 | 0.0111 | 2.4227 | 0.5084 | 0.8697 | 0.3125 |
Table 6.
Clustering of 34 elite rice breeding lines.
Table 6.
Clustering of 34 elite rice breeding lines.
| Cluster | Genotypes Number | Name of Genotypes Code |
|---|
| I | 8 | Gen1, Gen2, Gen3, Gen4, Gen8, Gen9, Gen23, Gen29 |
| II | 13 | Gen6, Gen7, Gen12, Gen15, Gen18, Gen19, Gen24, Gen28, Gen30, Gen31, Gen32, Gen33 Gen34 |
| III | 13 | Gen5, Gen10, Gen11, Gen13, Gen14, Gen16, Gen17, Gen20, Gen21, Gen22, Gen25, Gen26, Gen27 |
Table 7.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Satkhira.
Table 7.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Satkhira.
| Variable | Factor | Xo | Xs | SDperc | h2 | SGperc | Sense | Goal |
|---|
| PPH | FA1 | 10.7 | 11.8 | 9.88 | 0.88 | 8.7 | increase | 100 |
| GP | FA1 | 210 | 244 | 16.3 | 0.873 | 14.3 | increase | 100 |
| TGW | FA1 | 20.7 | 20.4 | −1.88 | 0.841 | −1.58 | increase | 0 |
| X50F | FA2 | 115 | 115 | −0.117 | 0.98 | −0.115 | increase | 0 |
| PH | FA2 | 101 | 101 | −0.276 | 0.963 | −0.266 | increase | 0 |
| TILL | FA2 | 11.2 | 11.5 | 2.82 | 0.91 | 2.57 | increase | 100 |
| YPP | FA3 | 22.8 | 25.6 | 12.4 | 0.904 | 11.2 | increase | 100 |
| YTH | FA3 | 6.24 | 6.67 | 6.84 | 0.941 | 6.43 | increase | 100 |
| PL | FA4 | 23.8 | 25.1 | 5.29 | 0.836 | 4.42 | increase | 100 |
| Fertility | FA4 | 80.2 | 76.9 | −4.12 | 0.816 | −3.36 | increase | 0 |
Table 8.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Kushtia.
Table 8.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Kushtia.
| Variable | Factor | Xo | Xs | SDperc | h2 | SGperc | Sense | Goal |
|---|
| YPP | FA1 | 28.6 | 31.9 | 11.4 | 0.874 | 9.96 | increase | 100 |
| YTH | FA1 | 7.04 | 7.88 | 11.9 | 0.892 | 10.6 | increase | 100 |
| TILL | FA2 | 15.7 | 16 | 1.89 | 0.767 | 1.45 | increase | 100 |
| PPH | FA2 | 10.4 | 10.4 | 0.0471 | 0.624 | 0.0294 | increase | 100 |
| GP | FA2 | 267 | 267 | −0.108 | 0.914 | −0.0989 | increase | 0 |
| X50F | FA3 | 121 | 124 | 2.31 | 0.984 | 2.28 | increase | 100 |
| PH | FA3 | 99.1 | 105 | 6.01 | 0.952 | 5.72 | increase | 100 |
| TGW | FA4 | 22.2 | 24.1 | 8.6 | 0.973 | 8.36 | increase | 100 |
| PL | FA5 | 24.4 | 25.2 | 3.48 | 0.728 | 2.53 | increase | 100 |
| Fertility | FA5 | 79.8 | 78.1 | −2.03 | 0.842 | −1.71 | increase | 0 |
Table 9.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Barishal.
Table 9.
Original value (Xo), selected value (Xs), selection differential in percentage (SDperc), heritability (h2), and selection gain in percent (SGperc) for the MGIDI in 34 rice genotypes in Barishal.
| Variable | Factor | Xo | Xs | SDperc | h2 | SGperc | Sense | Goal |
|---|
| YPP | FA1 | 24.2 | 26 | 7.75 | 0.918 | 7.11 | increase | 100 |
| YTH | FA1 | 6.41 | 6.93 | 8.20 | 0.905 | 7.42 | increase | 100 |
| TILL | FA2 | 13.9 | 15 | 8.31 | 0.827 | 6.87 | increase | 100 |
| PPH | FA2 | 12.5 | 14.1 | 12.6 | 0.863 | 10.8 | increase | 100 |
| PH | FA3 | 106 | 112 | 5.92 | 0.956 | 5.66 | increase | 100 |
| GP | FA3 | 307 | 323 | 5.07 | 0.866 | 4.39 | increase | 100 |
| PL | FA3 | 23.9 | 24.9 | 4.15 | 0.826 | 3.43 | increase | 100 |
| Fertility | FA3 | 66.1 | 60.1 | −9.06 | 0.929 | −8.41 | increase | 0 |
| X50F | FA4 | 118 | 114 | −3.79 | 0.994 | −3.77 | increase | 0 |
| TGW | FA4 | 21.9 | 23.4 | 6.96 | 0.948 | 6.6 | increase | 100 |



 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}