
Genomic Analysis of Non-B Nucleic Acids Structures in SARS-CoV-2: Potential Key Roles for These Structures in Mutability, Translation, and Replication?


Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Sequences
2.2. Detection of Mutations within IRs, Prediction of Pseudoknot Formation, and G4-Analysis
2.3. Statistical Analysis
3. Results
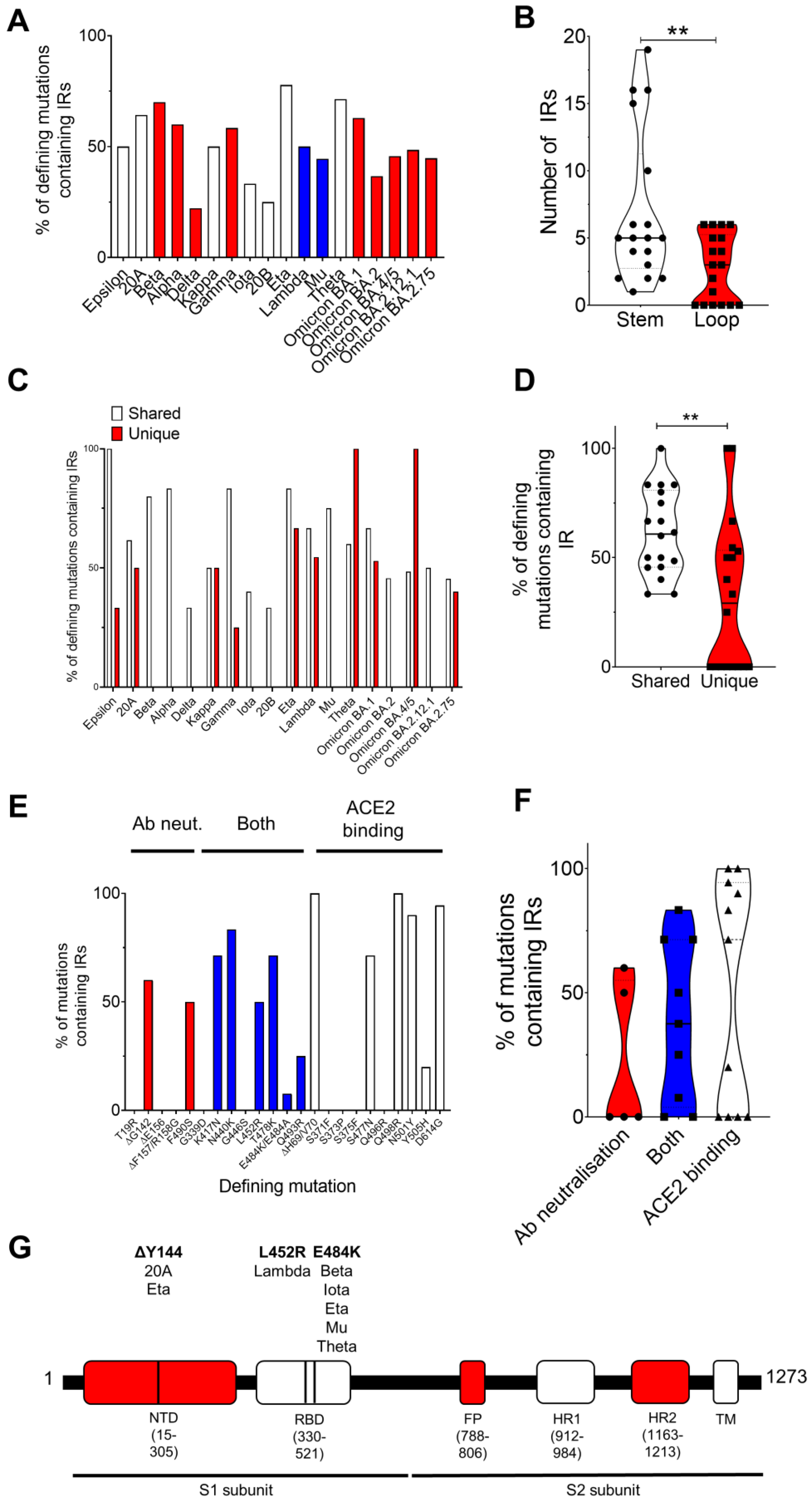
3.1. There Is a Large Variation in the Number of Defining Mutations Falling within IRs between SARS-CoV-2 Variants
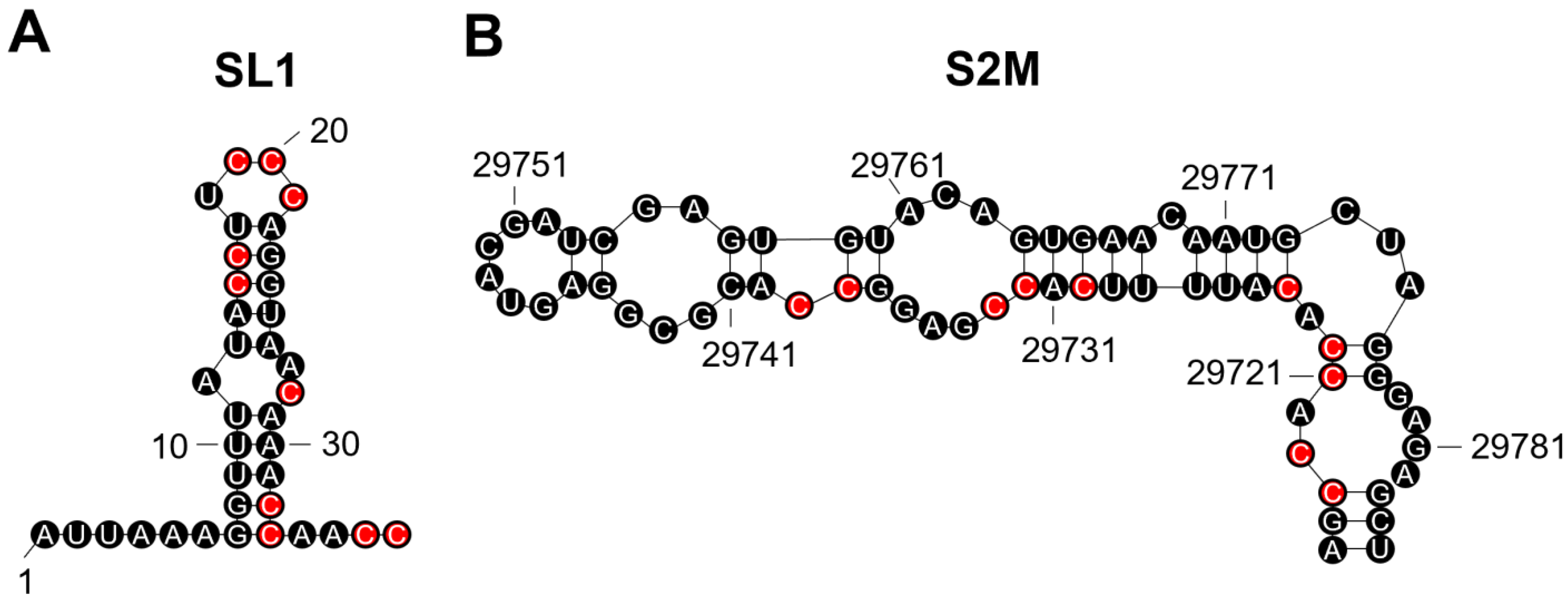
3.2. Pseudoknots Are Predicted to Occur near the Sites of Several Key Mutations
3.3. G4 Are Predicted to Form on the Negative Strand Genome in SARS-CoV-2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skourti-Stathaki, K.; Proudfoot, N.J. A Double-Edged Sword: R Loops as Threats to Genome Integrity and Powerful Regulators of Gene Expression. Genes Dev. 2014, 28, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Voineagu, I.; Narayanan, V.; Lobachev, K.S.; Mirkin, S.M. Replication Stalling at Unstable Inverted Repeats: Interplay between DNA Hairpins and Fork Stabilizing Proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 9936–9941. [Google Scholar] [CrossRef] [PubMed]
- Saranathan, N.; Vivekanandan, P. G-Quadruplexes: More Than Just a Kink in Microbial Genomes. Trends Microbiol. 2019, 27, 148–163. [Google Scholar] [CrossRef]
- Griffin, B.D.; Bass, H.W. Review: Plant G-Quadruplex (G4) Motifs in DNA and RNA; Abundant, Intriguing Sequences of Unknown Function. Plant Sci. 2018, 269, 143–147. [Google Scholar] [CrossRef]
- Brierley, I.; Pennell, S.; Gilbert, R.J.C. Viral RNA Pseudoknots: Versatile Motifs in Gene Expression and Replication. Nat. Rev. Microbiol. 2007, 5, 598–610. [Google Scholar] [CrossRef]
- Pearson, C.E.; Zorbas, H.; Price, G.B.; Zannis-Hadjopoulos, M. Inverted Repeats, Stem-Loops, and Cruciforms: Significance for Initiation of DNA Replication. J. Cell. Biochem. 1996, 63, 1–22. [Google Scholar] [CrossRef]
- del Solar, G.; Giraldo, R.; Ruiz-Echevarría, M.J.; Espinosa, M.; Díaz-Orejas, R. Replication and Control of Circular Bacterial Plasmids. Microbiol. Mol. Biol. Rev. 1998, 62, 434–464. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, G.; Bacolla, A.; Zhao, J.; Spitser, S.; Vasquez, K.M. Short Inverted Repeats Are Hotspots for Genetic Instability: Relevance to Cancer Genomes. Cell Rep. 2015, 10, 1674–1680. [Google Scholar] [CrossRef]
- Sadler, J.R.; Sasmor, H.; Betz, J.L. A Perfectly Symmetric Lac Operator Binds the Lac Repressor Very Tightly. Proc. Natl. Acad. Sci. USA 1983, 80, 6785–6789. [Google Scholar] [CrossRef]
- Butler, D.K.; Yasuda, L.E.; Yao, M.C. Induction of Large DNA Palindrome Formation in Yeast: Implications for Gene Amplification and Genome Stability in Eukaryotes. Cell 1996, 87, 1115–1122. [Google Scholar] [CrossRef]
- Okamura, K.; Chung, W.-J.; Lai, E.C. The Long and Short of Inverted Repeat Genes in Animals: MicroRNAs, Mirtrons and Hairpin RNAs. Cell Cycle 2008, 7, 2840–2845. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, T.; Matvienko, M.; Piskurewicz, U.; Xu, H.; Martineau, B.; Wong, J.; Govindarajulu, M.; Kozik, A.; Michelmore, R.W. Distinctive Profiles of Small RNA Couple Inverted Repeat-Induced Post-Transcriptional Gene Silencing with Endogenous RNA Silencing Pathways in Arabidopsis. RNA 2014, 20, 1987–1999. [Google Scholar] [CrossRef] [PubMed]
- Staple, D.W.; Butcher, S.E. Pseudoknots: RNA Structures with Diverse Functions. PLoS Biol. 2005, 3, e213. [Google Scholar] [CrossRef] [PubMed]
- Neupane, K.; Munshi, S.; Zhao, M.; Ritchie, D.B.; Ileperuma, S.M.; Woodside, M.T. Anti-Frameshifting Ligand Active against SARS Coronavirus-2 Is Resistant to Natural Mutations of the Frameshift-Stimulatory Pseudoknot. J. Mol. Biol. 2020, 432, 5843–5847. [Google Scholar] [CrossRef] [PubMed]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The Regulation and Functions of DNA and RNA G-Quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Cebrián, R.; Belmonte-Reche, E.; Pirota, V.; de Jong, A.; Morales, J.C.; Freccero, M.; Doria, F.; Kuipers, O.P. G-Quadruplex DNA as a Target in Pathogenic Bacteria: Efficacy of an Extended Naphthalene Diimide Ligand and Its Mode of Action. J. Med. Chem. 2022, 65, 4752–4766. [Google Scholar] [CrossRef] [PubMed]
- Abiri, A.; Lavigne, M.; Rezaei, M.; Nikzad, S.; Zare, P.; Mergny, J.-L.; Rahimi, H.-R. Unlocking G-Quadruplexes as Antiviral Targets. Pharmacol. Rev. 2021, 73, 897–923. [Google Scholar] [CrossRef]
- Cantara, A.; Luo, Y.; Dobrovolná, M.; Bohalova, N.; Fojta, M.; Verga, D.; Guittat, L.; Cucchiarini, A.; Savrimoutou, S.; Häberli, C.; et al. G-Quadruplexes in Helminth Parasites. Nucleic Acids Res. 2022, 50, 2719–2735. [Google Scholar] [CrossRef]
- Warner, E.F.; Bohálová, N.; Brázda, V.; Waller, Z.A.E.; Bidula, S. Analysis of Putative Quadruplex-Forming Sequences in Fungal Genomes: Novel Antifungal Targets? Microb. Genom. 2021, 7, 000570. [Google Scholar] [CrossRef]
- Goswami, P.; Bartas, M.; Lexa, M.; Bohálová, N.; Volná, A.; Červeň, J.; Červeňová, V.; Pečinka, P.; Špunda, V.; Fojta, M.; et al. SARS-CoV-2 Hot-Spot Mutations Are Significantly Enriched within Inverted Repeats and CpG Island Loci. Brief. Bioinform. 2021, 22, 1338–1345. [Google Scholar] [CrossRef]
- Bartas, M.; Goswami, P.; Lexa, M.; Červeň, J.; Volná, A.; Fojta, M.; Brázda, V.; Pečinka, P. Letter to the Editor: Significant Mutation Enrichment in Inverted Repeat Sites of New SARS-CoV-2 Strains. Brief. Bioinform. 2021, 22, bbab129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Qin, G.; Niu, J.; Wang, Z.; Wang, C.; Ren, J.; Qu, X. Targeting RNA G-Quadruplex in SARS-CoV-2: A Promising Therapeutic Target for COVID-19? Angew. Chem. Int. Ed. 2021, 60, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Brázda, V.; Kolomazník, J.; Lýsek, J.; Hároníková, L.; Coufal, J.; Štastný, J. Palindrome Analyser—A New Web-Based Server for Predicting and Evaluating Inverted Repeats in Nucleotide Sequences. Biochem. Biophys. Res. Commun. 2016, 478, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Hodcroft, E.B. CoVariants: SARS-CoV-2 Mutations and Variants of Interest. Available online: https://covariants.org/ (accessed on 19 December 2022).
- Shu, Y.; McCauley, J. GISAID: Global Initiative on Sharing All Influenza Data—From Vision to Reality. Euro Surveill. 2017, 22, 30494. [Google Scholar] [CrossRef] [PubMed]
- Smit, S.; Rother, K.; Heringa, J.; Knight, R. From Knotted to Nested RNA Structures: A Variety of Computational Methods for Pseudoknot Removal. RNA 2008, 14, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Bellaousov, S.; Mathews, D.H. ProbKnot: Fast Prediction of RNA Secondary Structure Including Pseudoknots. RNA 2010, 16, 1870–1880. [Google Scholar] [CrossRef]
- Kikin, O.; D’Antonio, L.; Bagga, P.S. QGRS Mapper: A Web-Based Server for Predicting G-Quadruplexes in Nucleotide Sequences. Nucleic Acids Res. 2006, 34, W676–W682. [Google Scholar] [CrossRef]
- Yang, T.-J.; Yu, P.-Y.; Chang, Y.-C.; Liang, K.-H.; Tso, H.-C.; Ho, M.-R.; Chen, W.-Y.; Lin, H.-T.; Wu, H.-C.; Hsu, S.-T.D. Effect of SARS-CoV-2 B.1.1.7 Mutations on Spike Protein Structure and Function. Nat. Struct. Mol. Biol. 2021, 28, 731–739. [Google Scholar] [CrossRef]
- Hirabara, S.M.; Serdan, T.D.A.; Gorjao, R.; Masi, L.N.; Pithon-Curi, T.C.; Covas, D.T.; Curi, R.; Durigon, E.L. SARS-CoV-2 Variants: Differences and Potential of Immune Evasion. Front. Cell. Infect. Microbiol. 2021, 11, 781429. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- McCallum, M.; Walls, A.C.; Sprouse, K.R.; Bowen, J.E.; Rosen, L.E.; Dang, H.V.; De Marco, A.; Franko, N.; Tilles, S.W.; Logue, J.; et al. Molecular Basis of Immune Evasion by the Delta and Kappa SARS-CoV-2 Variants. Science 2021, 374, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.E.; Zhang, X.; Case, J.B.; Winkler, E.S.; Liu, Y.; VanBlargan, L.A.; Liu, J.; Errico, J.M.; Xie, X.; Suryadevara, N.; et al. Resistance of SARS-CoV-2 Variants to Neutralization by Monoclonal and Serum-Derived Polyclonal Antibodies. Nat. Med. 2021, 27, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.I.; MacGowan, S.A.; Kutuzov, M.A.; Dushek, O.; Barton, G.J.; van der Merwe, P.A. Effects of Common Mutations in the SARS-CoV-2 Spike RBD and Its Ligand, the Human ACE2 Receptor on Binding Affinity and Kinetics. Elife 2021, 10, e70658. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, J.A.; Cunningham, C.L.; Pellegrene, K.A.; Brinson, R.G.; Marino, J.P.; Evanseck, J.D.; Mihailescu, M.R. Highly Conserved S2m Element of SARS-CoV-2 Dimerizes via a Kissing Complex and Interacts with Host MiRNA-1307-3p. Nucleic Acids Res. 2022, 50, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Tandel, D.; Gupta, D.; Sah, V.; Harshan, K.H. N440K Variant of SARS-CoV-2 Has Higher Infectious Fitness. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bate, N.; Savva, C.G.; Moody, P.C.E.; Brown, E.A.; Evans, S.E.; Ball, J.K.; Schwabe, J.W.R.; Sale, J.E.; Brindle, N.P.J. In Vitro Evolution Predicts Emerging SARS-CoV-2 Mutations with High Affinity for ACE2 and Cross-Species Binding. PLoS Pathog. 2022, 18, e1010733. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence That D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef]
- Tian, F.; Tong, B.; Sun, L.; Shi, S.; Zheng, B.; Wang, Z.; Dong, X.; Zheng, P. N501Y Mutation of Spike Protein in SARS-CoV-2 Strengthens Its Binding to Receptor ACE2. Elife 2021, 10, e69091. [Google Scholar] [CrossRef]
- Meng, B.; Kemp, S.A.; Papa, G.; Datir, R.; Ferreira, I.A.T.M.; Marelli, S.; Harvey, W.T.; Lytras, S.; Mohamed, A.; Gallo, G.; et al. Recurrent Emergence of SARS-CoV-2 Spike Deletion H69/V70 and Its Role in the Alpha Variant B.1.1.7. Cell Rep. 2021, 35, 109292. [Google Scholar] [CrossRef]
- Iketani, S.; Liu, L.; Guo, Y.; Liu, L.; Chan, J.F.-W.; Huang, Y.; Wang, M.; Luo, Y.; Yu, J.; Chu, H.; et al. Antibody Evasion Properties of SARS-CoV-2 Omicron Sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef]
- Zhou, H.; Dcosta, B.M.; Landau, N.R.; Tada, T. Resistance of SARS-CoV-2 Omicron BA.1 and BA.2 Variants to Vaccine-Elicited Sera and Therapeutic Monoclonal Antibodies. Viruses 2022, 14, 1334. [Google Scholar] [CrossRef] [PubMed]
- Yamasoba, D.; Kosugi, Y.; Kimura, I.; Fujita, S.; Uriu, K.; Ito, J.; Sato, K. Neutralisation Sensitivity of SARS-CoV-2 Omicron Subvariants to Therapeutic Monoclonal Antibodies. Lancet Infect. Dis. 2022, 22, 942–943. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 Escape Antibodies Elicited by Omicron Infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.-H.; Michailidis, E.; et al. Escape from Neutralizing Antibodies by SARS-CoV-2 Spike Protein Variants. Elife 2020, 9, e61312. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.D.; Chang, R.Y.; Brian, D.A. A Phylogenetically Conserved Hairpin-Type 3’ Untranslated Region Pseudoknot Functions in Coronavirus RNA Replication. J. Virol. 1999, 73, 8349–8355. [Google Scholar] [CrossRef] [PubMed]
- Belmonte-Reche, E.; Serrano-Chacón, I.; Gonzalez, C.; Gallo, J.; Bañobre-López, M. Potential G-Quadruplexes and i-Motifs in the SARS-CoV-2. PLoS ONE 2021, 16, e0250654. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, L. G-Quadruplexes Are Present in Human Coronaviruses Including SARS-CoV-2. Front. Microbiol. 2020, 11, 567317. [Google Scholar] [CrossRef] [PubMed]
- Dinan, A.M.; Lukhovitskaya, N.I.; Olendraite, I.; Firth, A.E. A Case for a Negative-Strand Coding Sequence in a Group of Positive-Sense RNA Viruses. Virus Evol. 2020, 6, veaa007. [Google Scholar] [CrossRef]
- Liu, G.; Du, W.; Sang, X.; Tong, Q.; Wang, Y.; Chen, G.; Yuan, Y.; Jiang, L.; Cheng, W.; Liu, D.; et al. RNA G-Quadruplex in TMPRSS2 Reduces SARS-CoV-2 Infection. Nat. Commun. 2022, 13, 1444. [Google Scholar] [CrossRef] [PubMed]
- Moraca, F.; Marzano, S.; D’Amico, F.; Lupia, A.; Di Fonzo, S.; Vertecchi, E.; Salvati, E.; Di Porzio, A.; Catalanotti, B.; Randazzo, A.; et al. Ligand-Based Drug Repurposing Strategy Identified SARS-CoV-2 RNA G-Quadruplex Binders. Chem. Commun. 2022, 58, 11913–11916. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Zhao, C.; Liu, Y.; Zhang, C.; Yang, G.; Yang, J.; Wang, Z.; Wang, C.; Tu, C.; Guo, Z.; et al. RNA G-Quadruplex Formed in SARS-CoV-2 Used for COVID-19 Treatment in Animal Models. Cell Discov. 2022, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.M.; Fontana, P.; Mao, T.; Leger, V.; Zhang, Y.; Fu, T.-M.; Lieberman, J.; Gehrke, L.; Shi, M.; Wang, L.; et al. Targeting Stem-Loop 1 of the SARS-CoV-2 5’ UTR to Suppress Viral Translation and Nsp1 Evasion. Proc. Natl. Acad. Sci. USA 2022, 119, e2117198119. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Wang, J.; Nuccio, S.P.; Mao, H.; Di Antonio, M. Short LNA-Modified Oligonucleotide Probes as Efficient Disruptors of DNA G-Quadruplexes. Nucleic Acids Res. 2022, 50, 7247–7259. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.-Y.; Contreras, G.P. Emerging Viral Mutants in Australia Suggest RNA Recombination Event in the SARS-CoV-2 Genome. Med. J. Aust. 2020, 213, 44–44.e1. [Google Scholar] [CrossRef]
- Xu, Z.; Choi, J.-H.; Dai, D.L.; Luo, J.; Ladak, R.J.; Li, Q.; Wang, Y.; Zhang, C.; Wiebe, S.; Liu, A.C.H.; et al. SARS-CoV-2 Impairs Interferon Production via NSP2-Induced Repression of MRNA Translation. Proc. Natl. Acad. Sci. USA 2022, 119, e2204539119. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variant | Pango Lineage | Defining Spike Protein Mutations | Defining Mutations within IRs |
|---|---|---|---|
| Epsilon | B.1.427 | S13I W152C L452R D614G | W152C D614G |
| 20A | B.1.620 | ΔP26, ΔH69/V70, V126A, ΔY144, ΔL241-A243, H245Y, S477N, E484K, D614G, P681H, T1027I, D1118H | ΔH69/V70, V126A, ΔY144, ΔL241-A243, D614G, D1118H |
| Beta | B.1.351 | D80A, D215G, ΔL241-A243, K417N, E484K, N501Y, D614G, A701V | ΔL241-A243, K417N, N501Y, D614G, A701V |
| Alpha | B.1.1.7 | ΔH69/V70, ΔY144, N501Y, A570D, D614G, P681H, T716I, S982A, D1118H | ΔH69/V70, ΔY144, N501Y, D614G, D1118H |
| Delta | B.1.617.2 | T19R, ΔE156/F157, R158G, L452R, T478K, D614G, P681R, D950N | D614G, D950N |
| Kappa | B.1.617.1 | E154K, L452R, E484Q, D614G, P681R, Q1071H | E484Q, D614G, Q1071H |
| Gamma | P.1 | L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, D614G, H655Y, T1027I, V1176F | L18F, T20N, R190S, K417T, N501Y, D614G, V1176F |
| Iota | B.1.526 | L5F, T95I, D253G, E484K, D614G, A701V | D614G, A701V |
| 20B | B.1.1.519 | T478K, D614G, P681R, T732A | D614G |
| Eta | B.1.525 | Q52R, A67V, ΔH69/V70, ΔY144, E484K, D614G, Q677H, F888L | Q52R, A67V, ΔH69/V70, ΔY144, D614G, F888L |
| Lambda | C.37 | G75V, T76I, ΔR246-G252, D253N, L452Q, F490S, D614G, T859N | G75V, T76I, ΔR246-G252, L452Q, F490S, D614G |
| Mu | B.1.621 | T95I, Y144S, Y145N, R346K, E484K, N501Y, D614G, P681R, D950N | Y144S, Y145N, D614G, D950N |
| Theta | P.3 | E484K, N501Y, D614G, P681R, E1092K, H1101Y, V1176F | N501Y, D614G, E1092K, H1101Y, V1176F |
| Omicron | BA.1 | A67V, ΔH69/V70, T95I, ΔG142-Y144, Y145D, ΔN211, L212I, G339D, S371L, S373P, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, Q496R, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F | A67V, ΔH69/V70, ΔG142-Y144, Y145D, ΔN211, L212I, N440K, S477N, T478K, Q498R, N501Y, Y505H, D614G, N679K, N764K, D796Y, Q954H, N969K, L981F |
| Omicron | BA.2 | T19I, ΔL24-P26, A27S, G142D, V213G, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, S477N, T478K, E484A, Q493R, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, Q954H, N969K | G142D, V213G, N440K, S477N, T478K, Q498R, N501Y, N764K, D796Y, Q954H, N969K |
| Omicron | BA.4/BA.5 | T19I, ΔL24-P26, A27S, ΔH69/V70, G142D, V213G, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452R, S477N, T478K, E484A, F486V, Q498R, N501Y, Y505H, D614G, H665Y, N679K, P681H, N764K, D796Y, Q954H, N969K | ΔH69/V70, G142D, V213G, N440K, L452R, S477N, T478K, F486V, Q498R, N501Y, H665Y, N764K, D796Y, Q954H, N969K |
| Omicron | BA.2.12.1 | T19I, ΔL24-P26, A27S, G142D, V213G, G339D, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, L452Q, S477N, T478K, E484A, Q493A, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, S704L, N764K, D796Y, Q954H, N969K | G142D, V213G, D405N, K417N, N440K, L452Q, S477N, T478K, Q498R, N501Y, D614G, N679K, N764K, D796Y, Q954H, N969K |
| Omicron | BA.2.75 | T19I, ΔL24-P26, A27S, G142D, K147E, W152R, F157L, I210V, V213G, G257S, G339H, S371F, S373P, S375F, T376A, D405N, R408S, K417N, N440K, G446S, N460K, S477N, T478K, E484A, R493Q, Q498R, N501Y, Y505H, D614G, H655Y, N679K, P681H, N764K, D796Y, Q954H, N969K | K147E, V213G, D405N, K417N, N440K, N460K, S477N, T478K, R493Q, Q498R, N501Y, D614G, N679K, N764K, D796Y, Q954H, N969K |
| Gene Name/Region | Highest Scoring Sequence | + or − Strand |
|---|---|---|
| nsp1 | GGCTTTGGAGACTCCGTGGAGGAGG | + |
| nsp2 | GGTGTTGTTGGAGAAGGTTCCGAAGG | + |
| nsp3 | GGATATGGTTGGTTTGG | − |
| nsp4 | GGTGATAGAGGTTTGTGGTGGTTGG | − |
| nsp10 | GGTATGTGGAAAGGTTATGG | + |
| nsp12 | GGAACCACTAAATTTTATGGTGGTTGG | + |
| nsp14 | GGTTGGGTTGGTTTTGATGTTGAAGG | + |
| nsp15 | GGAGCCCACAAGGTAATCCAGGTGG | + |
| nsp16 | GGAGAAATAGTACAACATGGAATGGCGG | + |
| S | GGCTTATAGGTTTAATGGTATTGG | + |
| N | GGCTGGCAATGGCGG | + |
| 3′UTR | GGUGGUGTAAAAGUGGCUCCGG | − |
| 5′UTR | GGAAGGGUCCAUUGUUUGGUUGG | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bidula, S.; Brázda, V. Genomic Analysis of Non-B Nucleic Acids Structures in SARS-CoV-2: Potential Key Roles for These Structures in Mutability, Translation, and Replication? Genes 2023, 14, 157. https://doi.org/10.3390/genes14010157
Bidula S, Brázda V. Genomic Analysis of Non-B Nucleic Acids Structures in SARS-CoV-2: Potential Key Roles for These Structures in Mutability, Translation, and Replication? Genes. 2023; 14(1):157. https://doi.org/10.3390/genes14010157
Chicago/Turabian StyleBidula, Stefan, and Václav Brázda. 2023. "Genomic Analysis of Non-B Nucleic Acids Structures in SARS-CoV-2: Potential Key Roles for These Structures in Mutability, Translation, and Replication?" Genes 14, no. 1: 157. https://doi.org/10.3390/genes14010157
APA StyleBidula, S., & Brázda, V. (2023). Genomic Analysis of Non-B Nucleic Acids Structures in SARS-CoV-2: Potential Key Roles for These Structures in Mutability, Translation, and Replication? Genes, 14(1), 157. https://doi.org/10.3390/genes14010157