
Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker


Abstract
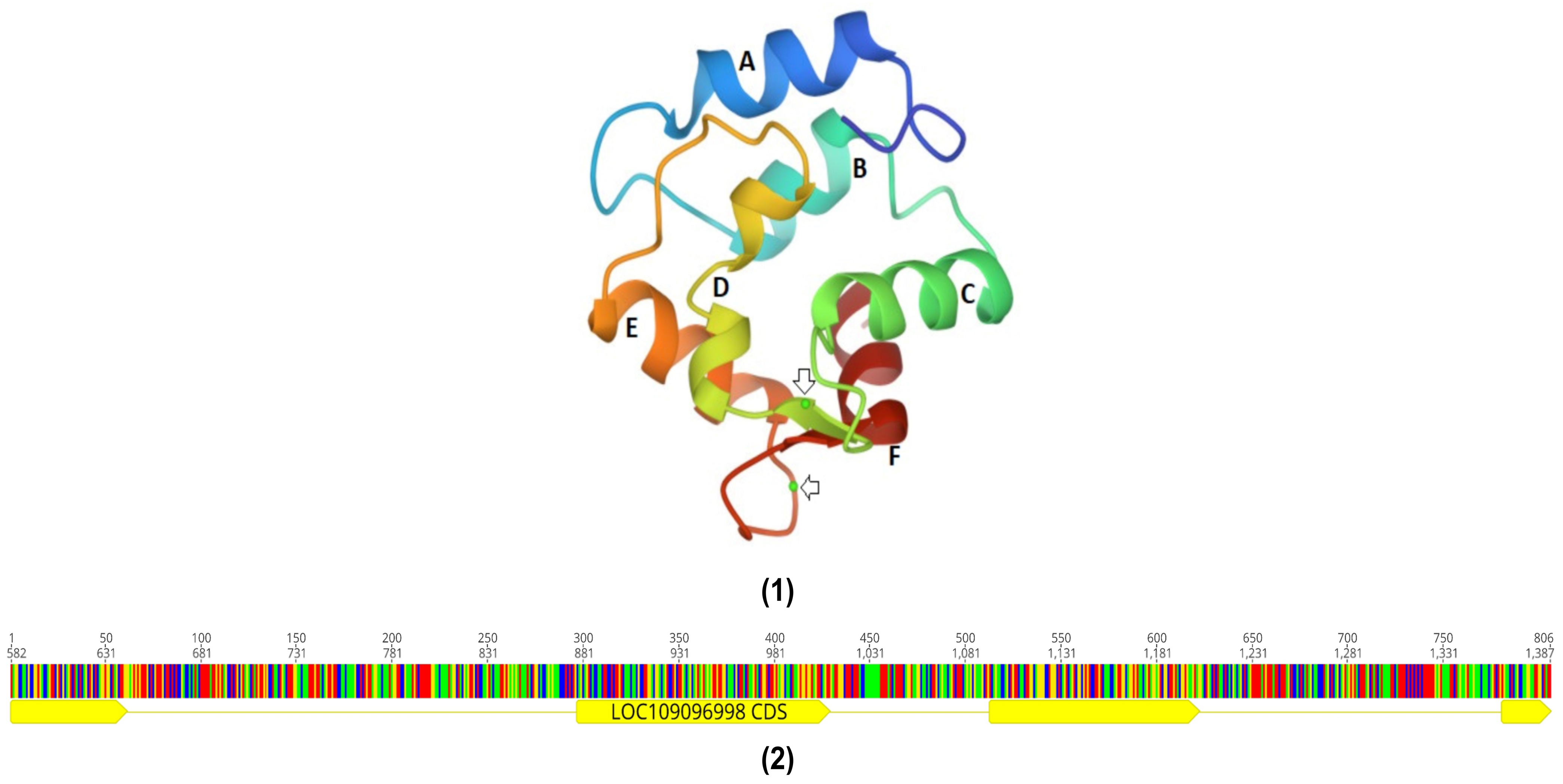
1. Introduction
2. Parvalbumin Gene Diversity
3. Parvalbumin—A Major Fish Allergen

{kind=link}
{kind=link}
{kind=link}
| Name of Allergen | Organism | Biochemical Name | Genbank Nucleotide Accession no. | Reference | |
|---|---|---|---|---|---|
| Scientific Name | Common Name | ||||
| Fish species | |||||
| Clu h 1 | Clupea harengus | Atlantic herring | β-parvalbumin | FM178220 | [61] |
| FM178221 | |||||
| FM178222 | |||||
| Cten i 1 | Ctenopharyngodon idella | Grass carp | β-parvalbumin | MK140606 | [62] |
| Cyp c 1 | Cyprinus carpio | Common carp | β-parvalbumin | AJ292211 | [63] |
| AJ292212 | |||||
| Gad c 1 | Gadus callarias | Baltic cod | β-parvalbumin | [64] | |
| Gad m 1 | Gadus morhua | Atlantic cod | β-parvalbumin | AY035584 | [65] |
| AM497927 | |||||
| AY035585 | |||||
| AM497928 | |||||
| lat c 1 | Lates calcarifer | Baramundi | β-parvalbumin | AY688372 | [66] |
| KF021278 | |||||
| AY626068 | |||||
| KF021279 | |||||
| AY688373 | |||||
| Lep w 1 | Lepidorhombus whiffiagonis | Megrim, whiff, turbot fish | β-parvalbumin | AM904681 | [19] |
| Onc m 1 | Oncorhynchus mykiss | Rainbow trout | β-parvalbumin | not specified | [67] |
| Pan h 1 | Pangasianodon hypophthalmus | Striped catfish | β-parvalbumin | XM_026916202 | [68] |
| XM_026947968 | |||||
| Ras k 1 | Rastrelliger kanagurta | Indian mackerel | parvalbumin | KX527884 | [69] |
| Sal s 1 | Salmo salar | Atlantic salmon | β-parvalbumin 1 | X97824 | [70] |
| Sar sa 1 | Sardinops sagax | Pacific pilchard | β-parvalbumin | FM177701 | [47] |
| Sco s 1 | Scomber scombrus | Atlantic mackerel | parvalbumin | FM994926 | [48] |
| Seb m 1 | Sebastes marinus | Ocean perch, redfish | β-parvalbumin | FM178218 | [71] |
| FM178219 | |||||
| Sole s 1 | Solea solea | Sole | parvalbumin | [72] | |
| Thu a 1 | Thunnus albacares | Yellow fin | β-parvalbumin | FM178217 | [73] |
| Xip g 1 | Xiphias gladius | Swordfish | β-parvalbumin | FM202668 | [19] |
| Non-fish Species | |||||
| Cro p 1 | Crocodylus porosus | Australian saltwater crocodile | β-parvalbumin | XM_019542160 | [74] |
| Cro p 2 | α-parvalbumin | XM_019544844 | |||
| Gal d 8 | Gallus domesticus | Chicken | α-parvalbumin | FM994924 | [75] |
| [76] | |||||
| Ran e 1 | Rana esculenta (Pelophylax esculentus) | Edible frog | α-parvalbumin | AJ315959 | [77] |
| Ran e 2 | Rana esculenta (Pelophylax esculentus) | Edible frog | β-parvalbumin | AJ414730 | |
4. Forensic Application of Parvalbumin
4.1. Protein-Based Assays
4.2. Using DNA-Based Assays for Fish Identification
| Organism | Primer Name | Primer Sequence (5′-3′) | Amplification Length (BP) | Amplification Region | Reference | |
|---|---|---|---|---|---|---|
| Scientific Name | Common Name | |||||
| Universal | IFF232 | GACAAGAGCGGCTTCATTGAGG | 268 | β-parvalbumin | [122] | |
| IFF233 | TCAACTCCAATCTTGCCATCACCAT | Exon 3 to Exon 4 | ||||
| Universal | IFF 233a | TCAATACCGATCTTGCCATCACCGT | NA | [123] | ||
| IFF 233b | TCAACTCCGATCATGCCATCACCAT | |||||
| Universal | SUN-F | CAGGACAAGAGTGGCTTCAT | 57 | β2-parvalbumin | [25] | |
| SUN-R | GAAGTTCTGCAGGAACAGCTT | Exon2 to Exon 3 | ||||
| probe | AGGAGGAYGAGCT | |||||
| C. harengus | Atlantic herring Pacific herring | CluHaPaF | CCGCTGATGATGTGAAGAAG | 189 | β2-parvalbumin | [124] |
| Clupea pallasii | CluHaPaR | GCAGGAACAGCCTGAGAGAG | Exon 2 to Exon 3 | |||
| Cyprinus carpio | Carp | MA-f | ACAAGCTTATGGCTTTCGCCGGAATTCTGA | β-parvalbumin | [125] | |
| MA-r | ATCGGATCCTATGCCTTGATCATGGC | |||||
| G. morhua | Atlantic cod | rGad m 1.01 | ATGGCATTCGCTGGAATTCTCG | 599 | ||
| rGad m 1.02 | ATGGCTTTCGCCGGAATTCTG | 797 | ||||
| Lophius piscatorius Lophius budegassa | White anglerfish | DAS-F | ACAACTTTCCCCGAGAAGC | 196 | β2-parvalbumin | [126] |
| Black-bellied anglerfish | DAS-R | ACAACATCACAGTTTAAGTTTTGC | Exon 2 to Exon 3 | |||
| Oncorhynchus mykiss | Rainbow trout | 601F9 forward | AGACAGAGACACAGGTTGGCTTACTATTCT | 75 | β-parvalbumin | [24] |
| 601G0 reverse | TTTACGACATAGGGAGCAGCTTACTATTCT | Intron 2 | ||||
| Paralichthys olivaceus | Japanese flounder | sunF | GATGACACCATATGTCTCTGGCATCTAAGCTGTCTG | 327 | β-parvalbumin | [127] |
| sunR | GTGTCCTCGAGTTACTGTTTCACCATCGCCGC | |||||
| S. salar | Atlantic salmon | 601F5 forward | AGACAGAGACACAGGTTGGCTTACTATTCT | 126 | β-parvalbumin | [24] |
| 601F6 reverse | TTTACGACATAGGGAGCAGCTTACTATTCT | Intron 2 | ||||
| S. salar | Atlantic salmon | Sense PV | AGYGGCTTYATHGARGARGAYGARYT | 430 | β2-parvalbumin | [70] |
| Antisense PV1 | YTGYTTNACNAANACNGCRAAYTC | Exon 2 to Exon 4 | ||||
| Antisense PV2 | GAATTCRTCRACHCCDATYTTHCC | |||||
| S. salar | Atlantic salmon | IFF 156 | ATGGCCTGTGCCCATCTGTGC | 300 | β1-parvalbumin | [122] |
| IFF 157 | GGACTTCGAGGCAAAGCCAAT | Exon 1 to Exon 2 | ||||
| S. salar | Atlantic salmon | Psal1 | CTGTGCCCATCTGTGCAAGG | 650 | β1-parvalbumin | [128] |
| Oncorhynchus mykiss | Rainbow trout | Psal2 | CCAATCATGCCATCACCATCG | Exon1 to Exon 3 | ||
| Salmo trutta | Brown trout | Psal3 | TACCGATGCAGAGACAAAGG | 931 | β1-parvalbumin | |
| Salvelinus alpinus | Arctic char | Psal4 | GTCTTGGGCAATATTGTTCC | 3′ end of Exon 3 to Exon 4 | ||
| Scomber japonicus | Mackerel | SJ9 | CCCTACAAAGCAAAAACATC | 1500 | β-parvalbumin | [129] |
| SJ487 | GCATAGGAGGAAAGGICTCT | |||||
| SJ106 | GTAGITTCGACCACAAAAAGTT | 190 | β-parvalbumin | |||
| SJG441r | ACTGCTGTATAGGTGATAGG | Exon 2 to Intron 2 | ||||
| SJG107f | AGCTATTCTGTATCGCTTCG | 284 | β-parvalbumin | |||
| SGG297r | GGTGTGAGTCTTACTTCAGC | Intron 1 to Intron 2 | ||||
| S. scombrus Trachurus trachurus | Atlantic mackerel Atlantic horse mackerel | Pval1Fw | CTGAAGCTGTTCCTGCAGAACTT | 87 | β-parvalbumin | [130] |
| Pval1Rev | GCTGTCACCGGCCTTGAG | |||||
| Pval1Probe | [6FAM]TCCGACGCCGAGACCAAGGC[TAM] | Intron 2 to Exon 3 | ||||
| Spondyliosoma cantharus | Black seabream | 1189B6 | TGAGCTGAAGTAAGACACTCAGGAA | 78 | β-parvalbumin | [23] |
| 1189B7 | TCTAAAATGTTGTCTTGGTGCCTTAG | |||||
| 1273H9(probe) | TGCACACTTGAGCAAGCAATGGCC | Intron 2 | ||||
| Spondyliosoma cantharus | Black seabream | 601F7 forward | AGACAGAGACACAGGTTGGCTTACTATTCT | 79 | β-parvalbumin | [24] |
| 601F8 reverse | TTTACGACATAGGGAGCAGCTTACTATTCT | Intron 2 | ||||
| Thunnus albacares | Yellowfin tuna | ALB4F | AGGATTGGATTTTCTGTCTTAGCTT | 227 | β-parvalbumin | [22] |
| ALB4R | TCAGTTTGTGTCAATTGGTCTGTAG | Intron 2 | ||||
| PARVT1F | GGGGTTGGAGATGAATGGCA | 785 | β-parvalbumin | |||
| PARVT1R | GAGTCACCGGCCATGAGAAA | Intron 1 to Exon 3 | ||||
| PARVT2F | ACAGCTGCCGACTCTTTCAA | 670 | Parvalbumin β | |||
| PARVT2R | CGGCCATGAGAAATGCCTTG | Intron 1/Exon 2 to Exon 3 | ||||
4.3. Methods of Parvalbumin Allergen Quantification
5. Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arif, S.H.; Jabeen, M.; Hasnain, A. Biochemical characterisation and thermostable capacity of parvalbumins: The major fish-food allergens. J. Food. Biochem. 2007, 31, 121–137. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bartoš, O.; Zdeňková, K.; Hanák, P.; Horká, P.; Musilova, Z. Evolution of the Parvalbumin Genes in Teleost Fishes after the Whole-Genome Duplication. Fishes 2021, 6, 70. [Google Scholar] [CrossRef]
- Deuticke, H.J. Űber die Sedimentationskonstante von Muskelproteinen. Physiol. Chem. 1934, 224, 216–228. [Google Scholar] [CrossRef]
- Henrotte, J.G. A crystalline constituent from myogen of carp muscles. Nature 1952, 169, 968–969. [Google Scholar] [CrossRef] [PubMed]
- Pechère, J.F. Muscular parvalbumins as homologous proteins. Comp. Biochem. Physiol. 1968, 24, 289–295. [Google Scholar] [CrossRef]
- Pechère, J.F.; Capony, J.; Ryden, L. The primary structure of the major parvalbumin from hake muscle. Eur. J. Biochem. 1971, 23, 421–428. [Google Scholar] [CrossRef]
- Nockolds, C.E.; Kretsinger, R.H.; Coffee, C.J.; Bradshaw, R.A. Structure of a calcium binding carp myogen. Proc. Natl. Acad. Sci. USA 1972, 69, 581–584. [Google Scholar] [CrossRef]
- Arif, S.H. A Ca2+-binding protein with numerous roles and uses: Parvalbumin in molecular biology and physiology. BioEssays 2009, 31, 410–421. [Google Scholar] [CrossRef]
- Heizmann, C.W. Ca2+-Binding proteins of the EF-hand superfamily: Diagnostic and prognostic biomarkers and novel therapeutic targets. Methods Mol. Biol. 2019, 1929, 157–186. [Google Scholar] [PubMed]
- Nogueira, L.; Gilmore, N.K.; Hogan, M.C. Role of parvalbumin in fatigue-induced changes in force and cytosolic calcium transients in intact single mouse myofibers. J. Appl. Physiol. 2022, 132, 1041–1053. [Google Scholar] [CrossRef]
- Ge, M.; Chen, S.; Huang, Y.; Chen, W.; He, L.; Zhang, Y. Role of calcium homeostasis in Alzheimer’s Disease. Neuropsychiatr. Dis. Treat. 2022, 18, 487. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, M.W.; Brinkmeier, H.; Műntener, M. Calcium ion in skeletal muscle: Its crucial role for muscle function, plasticity and disease. Physiol. Rev. 2000, 80, 1215–1265. [Google Scholar] [CrossRef] [PubMed]
- Gillis, J.M. Relaxation of vertebrate skeletal muscle. A synthesis of the biochemical and physiological approaches. Biochem. Biophys. Acta 1985, 811, 97–145. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, D.J. Aerobic muscle functions during steady swimming in fishes. Fish Fish. 2002, 3, 63–78. [Google Scholar] [CrossRef]
- Celio, M.R.; Heizmann, C.W. Calcium-binding protein parvalbumin as a neuronal marker. Nature 1981, 293, 300–302. [Google Scholar] [CrossRef]
- Radauer, C.; Bublin, M.; Wagner, S.; Mari, A.; Breiteneder, H. Allergens are distributed into few protein families and possess a restricted number of biochemical functions. J. Allergy Clin. Immunol. 2006, 121, 847–852. [Google Scholar] [CrossRef]
- Lopata, A.L.; Jeebhay, M.F. Airborne seafood allergens as a cause of occupational allergy and asthma. Curr. Allergy Asthma Rep. 2013, 13, 288–297. [Google Scholar] [CrossRef]
- Saptarshi, S.R.; Sharp, M.F.; Kamath, S.D.; Lopata, A.L. Antibody reactivity to the major fish allergen parvalbumin is determined by isoforms and impact of thermal processing. Food Chem. 2014, 148, 321–328. [Google Scholar] [CrossRef]
- Griesmeier, U.; Vázquez-Cortés, S.; Bublin, M.; Radauer, C.; Ma, Y.; Briza, P.; Fernández-Rivas, M.; Breiteneder, H. Expression levels of parvalbumins determine allergenicity of fish species. Allergy 2010, 65, 191–198. [Google Scholar] [CrossRef]
- Kubota, H.; Kobayashi, A.; Kobayashi, Y.; Shiomi, K.; Hamada-Sato, N. Reduction in IgE reactivity of Pacific mackerel parvalbumin by heat treatment. Food Chem. 2016, 206, 78–84. [Google Scholar] [CrossRef]
- Pérez-Tavarez, R.; Moreno, H.M.; Borderias, J.; Loli-Ausejo, D.; Pedrosa, M.; Hurtado, J.L.; Rodriguez-Pérez, R.; Gasset, M. Fish muscle processing into seafood products reduces β-parvalbumin allergenicity. Food Chem. 2021, 364, 130308. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, A.; Rehbein, H. The differentiation of tuna (family: Scombridae) products through the PCR-based analysis of the cytochrome b gene and parvalbumin introns. J. Sci. Food Agric. 2016, 96, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Akhatova, D.; Laknerova, I.; Zdenkova, K.; Olafsdottir, G.; Magnusdottir, S.; Piknova, L.; Kyrova, V.; Lerch, Z.; Hanak, P. International interlaboratory study on TaqMan real-time polymerase chain reaction authentication of black seabream (Spondyliosoma cantharus). J. Food Nutr. Res. 2018, 57, 27–37. [Google Scholar]
- Hanak, P.; Laknerova, I.; Svatora, M. Second intron in the protein-coding region of the fish parvalbumin gene-a promising platform for polymerase chain reaction-based discrimination of fish meat of various species. J. Food Nutr. Res. 2012, 51, 81–88. [Google Scholar]
- Sun, M.; Liang, C.; Gao, H.; Lin, C.; Deng, M. Detection of parvalbumin, a common fish allergen gene in food, by real-time polymerase chain reaction. J. AOAC Int. 2009, 92, 234–240. [Google Scholar] [CrossRef]
- Ross, C.; Tilghman, R.W.; Hartmann, J.X.; Mari, F. Distribution of parvalbumin isotypes in adult snook and their potential applications as species-specific biomarkers. J. Fish. Biol. 1997, 51, 561–572. [Google Scholar] [CrossRef]
- Huriaux, F.; Vandewalle, P.; Focant, B. Immunological study of muscle parvalbumin isotypes in three African catfish during development. Comp. Biochem. Physiol. 2002, 132B, 579–584. [Google Scholar] [CrossRef]
- Kuehn, A.; Swoboda, I.; Arumugam, K.; Hilger, C.; Hentges, F. Fish allergens at a glance: Variable allergenicity of parvalbumins, the major fish allergens. Front. Immunol. 2014, 5, 179. [Google Scholar] [CrossRef]
- Sharp, M.F.; Lopata, A.L. Fish allergy: In review. Clin. Rev. Allergy Immunol. 2014, 46, 258–271. [Google Scholar] [CrossRef]
- Goodman, M.; Pechére, J.F. The evolution of muscular parvalbumins investigated by the maximum parsimony method. J. Mol. Evol. 1977, 9, 131–158. [Google Scholar] [CrossRef]
- Leung, N.Y.; Wai, C.Y.; Shu, S.; Wang, J.; Kenny, T.P.; Chu, K.H.; Leung, P.S. Current immunological and molecular biological perspectives on seafood allergy: A comprehensive review. Clin. Rev. Allergy Immunol. 2014, 46, 180–197. [Google Scholar] [CrossRef] [PubMed]
- Rodenbaugh, D.W.; Wang, W.; Davis, J.; Edwards, T.; Potter, J.D.; Metzger, J.M. Parvalbumin isoforms differentially accelerate cardiac myocyte relaxation kinetics in an animal model of diastolic dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, 1705–1713. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, E.A.; Uversky, V.N. What Is Parvalbumin for? Biomolecules. 2022, 12, 656. [Google Scholar] [CrossRef] [PubMed]
- Haiech, J.; Moreau, M.; Leclerc, C.; Kilhoffer, M.C. Facts and conjectures on calmodulin and its cousin proteins, parvalbumin and troponin C. Biochim. Biophys. Acta-Mol. Cell Res. 2019, 1866, 1046–1053. [Google Scholar] [CrossRef]
- Cates, M.S.; Berry, M.B.; Ho, E.L.; Li, Q.; Potter, J.D.; Phillips, G.N., Jr. Metal-ion affinity and specificity in EF-hand proteins: Coordination geometry and domain plasticity in parvalbumin. Structure 1999, 7, 1269–1278. [Google Scholar] [CrossRef]
- Biomatters, Geneious version 2022.2. Available online: https://www.geneious.com (accessed on 6 December 2022).
- Cox, A.L.; Eigenmann, P.A.; Sicherer, S.H. Clinical relevance of cross-reactivity in food allergy. J. Allergy Clin. Immunol. Pract. 2021, 9, 82–99. [Google Scholar] [CrossRef]
- Meyer, A.; Van de Peer, Y. From 2R to 3R: Evidence for a fish-specific genome duplication (FSGD). Bioessays 2005, 27, 937–945. [Google Scholar] [CrossRef]
- Modrell, M.S.; Lyne, M.; Carr, A.R.; Zakon, H.H.; Buckley, D.; Campbell, A.S.; Davis, M.C.; Micklem, G.; Baker, C.V. Insights into electrosensory organ development, physiology and evolution from a lateral line-enriched transcriptome. eLife 2017, 6, e24197. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Uhlén, M. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Perez-Gordo, M.; Lin, J.; Bardina, L.; Pastor-Vargas, C.; Cases, B.; Vivanco, F.; Cuesta-Herranz, J.; Sampson, H.A. Epitope mapping of Atlantic salmon major allergen by peptide microarray immunoassay. Int. Arch. Allergy Immunol. 2012, 157, 31–40. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2006, 117, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Bannon, G.A. What makes a food protein an allergen? Curr. Allergy Asthma Rep. 2004, 4, 43–46. [Google Scholar] [CrossRef]
- Moreno, F.J. Gastrointestinal digestion of food allergens: Effect on their allergenicity. Biomed. Pharmacother. 2007, 61, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, C.; Zhu, Y.; Wang, Y. Seafood allergy: Occurrence, mechanisms and measures. Trends Food Sci. Technol. 2019, 88, 80–92. [Google Scholar] [CrossRef]
- Failler, P.; Van der Walle, G.; Lecrivain, N.; Himbes, A.; Lewins, R. Future Prospects for Fish and Fishery Products, European Overview; FAO Fisheries Circular No. 972/4, Part 1; FAO: Rome, Italy, 2007; 204p. [Google Scholar]
- Beale, J.E.; Jeebhay, M.F.; Lopata, A.L. Characterisation of purified parvalbumin from five fish species and nucleotide sequencing of this major allergen from Pacific pilchard, Sardinops sagax. Mol. Immunol. 2009, 46, 2985–2993. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Tanaka, H.; Ishizaki, S.; Ishida, M.; Nagashima, Y.; Shiomi, K. Purification, reactivity with IgE and cDNA cloning of parvalbumin as the major allergen of mackerels. Food Chem. Toxicol. 2003, 41, 1149–1156. [Google Scholar] [CrossRef]
- Helbling, A.; Heydet, R.; McCants, M.L.; Musmand, J.J.; El-Dahr, J.; Lehrer, S.G. Fish allergy: Is cross-reactivity among fish species relevant? Double-blind placebo-controlled food challenge studies or fish-allergic patients. Ann. Allergy Asthma Immunol. 1999, 83, 517–523. [Google Scholar] [CrossRef]
- Lim, D.L.C.; Neo, K.H.; Yi, F.C.; Chua, K.Y.; Goh, D.L.M.; Shek, L.P.C.; Giam, Y.C.; Van Bever, H.P.; Lee, B.W. Parvalbumin–the major tropical fish allergen. Pediatr. Allergy Immunol. 2008, 19, 399–407. [Google Scholar] [CrossRef]
- Borrego, J.T.; Cuevas, J.M.; Garcia, J.T. Cross reactivity between fish and shellfish. Allergol. Immunopathol. 2003, 31, 146–151. [Google Scholar]
- Madsen, C.B.; Hattersley, S.; Allen, K.J.; Beyer, K.; Chan, C.H.; Godefroy, S.B.; Hodgson, R.; Mills, E.N.; Muñoz-Furlong, A.; Schnadt, S.; et al. Can we define a tolerable level of risk in food allergy? Report from a EuroPrevall/UK food standards agency workshop. Clin. Exp. Allergy 2012, 42, 30–37. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.C.; Johnston, E.B.; Nugraha, R.; Le, T.T.; Kalic, T.; McLean, T.R.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef] [PubMed]
- Van Hengel, A.J. Introduction. In Food Allergens: Analysis Instrumentation and Methods; Nollet, L.M.L., van Hengel, A.J., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 1–11. [Google Scholar]
- Bernhisel-Broadbent, J.; Strause, D.; Sampson, H.A. Fish hypersensitivity. II. Clinical relevance of altered fish allergenicity caused by various preparation methods. J. Allergy Clin. Immunol. 1992, 90, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Pascual, C.Y.; Reche, M.; Fiandor, A.; Valbuena, T.; Cuevas, T.; Martin-Esteban, M.M. Fish allergy in childhood. Pediatr. Allergy Immunol. 2008, 19, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.W.; Nordlee, J.A.; Koppelman, S.J.; Baumert, J.L.; Taylor, S.L. Measuring parvalbumin levels in fish muscle tissue: Relevance of muscle locations and storage conditions. Food Chem. 2012, 135, 502–507. [Google Scholar] [CrossRef]
- Costa, J.; Ansari, P.; Mafra, I.; Oliveira, M.B.P.P.; Baumgartner, S. Assessing hazelnut allergens by protein- and DNA-based approaches: LC-MS/MS, ELISA and real-time PCR. Anal. Bioanal. Chem. 2014, 406, 2581–2590. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Directive 2007/68/EC of 27 November 2007 amending Annex IIIa to Directive 2000/13/EC of the European Parliament and of the Council as regards certain food ingredients. Off. J. Eur. Union 2007, 310, 11–14. [Google Scholar]
- Mari, A.; Scala, E.; Palazzo, P.; Ridolfi, S.; Zennaro, D.; Carabella, G. Bioinformatics applied to allergy: Allergen databases, from collecting sequence information to data integration. The allergome platform as a model. Cell Immunol. 2006, 244, 97–100. [Google Scholar] [CrossRef]
- Swoboda, I.; Balic, N.; Klug, C.; Focke, M.; Weber, M.; Spitzauer, S.; Neubauer, A.; Quirce, S.; Douladiris, N.; Papadopoulos, N.G.; et al. A general strategy for the generation of hypoallergenic molecules for the immunotherapy of fish allergy. J. Allergy Clin. Immunol. 2013, 132, 979–981. [Google Scholar] [CrossRef]
- Leung, N.Y.H.; Leung, A.S.Y.; Xu, K.J.Y.; Wai, C.Y.Y.; Lam, C.Y.; Wong, G.W.K.; Leung, T.F. Molecular and immunological characterization of grass carp (Ctenopharyngodon idella) parvalbumin Cten i 1: A major fish allergen in Hong Kong. Pediatr. Allergy Immunol. 2020, 31, 792–804. [Google Scholar] [CrossRef]
- Bugajska-Schretter, A.; Grote, M.; Vangelista, L.; Valent, P.; Sperr, W.R.; Rumpold, H.; Pastore, A.; Reichelt, R.; Valenta, R.; Spitzauer, S. Purification, biochemical, and immunological characterisation of a major food allergen: Different immunoglobulin E recognition of the apo-and calcium-bound forms of carp parvalbumin. Gut 2000, 46, 661–669. [Google Scholar] [CrossRef]
- Bugajska-Schretter, A.; Elfman, L.; Fuchs, T.; Kapiotis, S.; Rumpold, H.; Valenta, R.; Spitzauer, S. Parvalbumin, a cross-reactive fish allergen, contains IgE-binding epitopes sensitive to periodate treatment and Ca2+ depletion. J. Allergy Clin. Immunol. 1998, 101, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Van Do, T.; Hordvik, I.; Endresen, C.; Elsayed, S. The major allergen (parvalbumin) of codfish is encoded by at least two isotypic genes: cDNA cloning, expression and antibody binding of the recombinant allergens. Mol. Immunol. 2003, 39, 595–602. [Google Scholar] [CrossRef]
- Sharp, M.F.; Kamath, S.D.; Koeberl, M.; Jerry, D.R.; O’Hehir, R.E.; Campbell, D.E.; Lopata, A.L. Differential IgE binding to isoallergens from Asian seabass (Lates calcarifer) in children and adults. Mol. Immunol. 2014, 62, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Ahn, J.; Komatsubara, R.; Terada, A.; Yasuda, T.; Tsuge, I.; Urisu, A. Comparison of allergenic properties of salmon (Oncorhynchus nerka) between landlocked and anadromous species. Allergol. Int. 2009, 58, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Karnaneedi, S.; Nie, S.; Kalic, T.; Dai, D.; Daduang, S.; Leeming, M.; Williamson, N.A.; Breiteneder, H.; et al. Expanding the allergen repertoire of salmon and catfish. Allergy 2021, 76, 1443–1453. [Google Scholar] [CrossRef]
- Ruethers, T.; Raith, M.; Sharp, M.F.; Koeberl, M.; Stephen, J.N.; Nugraha, R.; Le, T.T.; Quirce, S.; Nguyen, H.X.; Kamath, S.D.; et al. Characterization of Ras k 1 a novel major allergen in Indian mackerel and identification of parvalbumin as the major fish allergen in 33 Asia-Pacific fish species. Clin. Exp. Allergy 2018, 48, 452–463. [Google Scholar] [CrossRef]
- Lindstrøm, C.D.V.; Van Do, T.; Hordvik, I.; Endresen, C.; Elsayed, S. Cloning of two distinct cDNAs encoding parvalbumin, the major allergen of Atlantic salmon (Salmo salar). Scand. J. Immunol. 1996, 44, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, K.G.; Hsieh, Y.H. Monoclonal antibody specific to a major fish allergen: Parvalbumin. J. Food Prot. 2009, 72, 818–825. [Google Scholar] [CrossRef]
- Kuehn, A. Allergen Nomenclature. 2021. Available online: http://allergen.org/viewallergen.php?aid=655 (accessed on 17 October 2022).
- Van Do, T.; Elsayed, S.; Florvaag, E.; Hordvik, I.; Endresen, C. Allergy to fish parvalbumins: Studies on the cross-reactivity of allergens from 9 commonly consumed fish. J. Allergy Clin. Immunol. 2005, 116, 1314–1320. [Google Scholar] [CrossRef]
- Ruethers, T.; Nugraha, R.; Taki, A.C.; O’Malley, A.; Karnaneedi, S.; Zhang, S.; Kapingidza, A.B.; Mehr, S.; Kamath, S.D.; Chruszcz, M.; et al. The first reptilian allergen and major allergen for fish-allergic patients: Crocodile β-parvalbumin. Pediatr. Allergy Immunol. 2022, 33, e13781. [Google Scholar] [CrossRef]
- Kuehn, A.; Lehners, C.; Hilger, C.; Hentges, F. Food allergy to chicken meat with IgE reactivity to muscle α-parvalbumin. Allergy 2009, 64, 1557–1558. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Codreanu-Morel, F.; Lehners-Weber, C.; Doyen, V.; Gomez-André, S.A.; Bienvenu, F.; Van Hage, M.; Perotin, J.M.; Silcret-Grieu, S. Cross-reactivity to fish and chicken meat–a new clinical syndrome. Allergy 2016, 71, 1772–1781. [Google Scholar] [CrossRef] [PubMed]
- Hilger, C.; Grigioni, F.; Thill, L.; Mertens, L.; Hentges, F. Severe IgE-mediated anaphylaxis following consumption of fried frog legs: Definition of α-parvalbumin as the allergen in cause. Allergy 2002, 57, 1053–1058. [Google Scholar] [CrossRef]
- Hubalkova, Z.; Kralik, P.; Tremlova, B.; Rencova, E. Methods of gadoid fish species identification in food and their economic impact in the Czech Republic: A review. Vet. Med. 2007, 52, 273. [Google Scholar] [CrossRef]
- Kim, H.; Kumar, K.S.; Hwang, S.Y.; Kang, B.C.; Moon, H.B.; Shin, K.H. Utility of Stable Isotope and Cytochrome Oxidase I Gene Sequencing Analyses in Inferring Origin and Authentication of Hairtail Fish and Shrimp. J. Agric. Food Chem. 2015, 63, 5548–5556. [Google Scholar] [CrossRef]
- Schmidhuber, J.; Pound, J.; Qiao, B. COVID-19: Channels of Transmission to Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Whitworth, J. High Risk Firms May Miss an Inspection Due to COVID-19. 2020. Available online: https://www.foodsafetynews.com/2020/12/high-risk-firms-may-miss-an-inspection-due-to-covid-19/ (accessed on 15 October 2022).
- Rehbein, H.; Kundiger, R.; Yman, I.M.; Ferm, M.; Etienne, M.; Jerome, M.; Craig, A.; Mackie, I.; Jessen, F.; Martinez, I.; et al. Species identification of cooked fish by urea isoelec-tric focusing and sodium dodecylsulfate polyacryla-mide gel electrophoresis: A collaborative study. Food Chem. 1999, 67, 333–339. [Google Scholar] [CrossRef]
- Piñeiro, C.; Sotelo, C.G.; Medina, I.; Gallardo, J.M.; Pérez-Martín, R.I. Reversed-phase HPLC as a method for the identification of gadoid fish species. Z. Lebensm. Unters. Forsch. 1997, 204, 411–416. [Google Scholar] [CrossRef]
- Céspedes, A.; García, T.; Carrera, E.; González, I.; Fernández, A.; Asensio, L.; Hernández, P.E.; Martín, R. Indirect enzyme-linked immunosorbent assay for the identification of sole (Solea solea), European plaice (Pleuronectes platessa), floun-der (Platichthys flesus), and Greenland hali-but (Reinhardtius hippoglossoides). J. Food Prot. 1999, 62, 1178–1182. [Google Scholar] [CrossRef]
- Hubalkova, Z.; Kralik, P.; Kasalova, J.; Rencova, E. Identification of gadoid species in fish meat by polymerase chain reaction (PCR) on genomic DNA. J. Agric. Food Chem. 2008, 56, 3454–3459. [Google Scholar] [CrossRef]
- Kochzius, M.; Seidel, C.; Antoniou, A.; Botla, S.K.; Campo, D.; Cariani, A.; Vazquez, E.G.; Hauschild, J.; Hervet, C.; Hjörleifsdottir, S.; et al. Identifying fishes through DNA barcodes and microarrays. PLoS One 2010, 5, e12620. [Google Scholar] [CrossRef]
- Hulley, E.N.; Tharmalingam, S.; Zarnke, A.; Boreham, D.R. Development and validation of probe-based multiplex real-time PCR assays for the rapid and accurate detection of freshwater fish species. PLoS One 2019, 14, e0210165. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, H. Differentiation of fish species by PCR-based DNA analysis of nuclear genes. Eur. Food Res. Technol. 2013, 236, 979–990. [Google Scholar] [CrossRef]
- Jiang, D.; Jiang, H.; Ji, J.; Sun, X.; Qian, H.; Zhang, G.; Tang, L. Mast-cell-based fluorescence biosensor for rapid detection of major fish allergen parvalbumin. J. Agric. Food Chem. 2014, 62, 6473–6480. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Yu, T.; Liu, D.; Xianyu, Y. Recent advances in gold nanoparticles-based biosensors for food safety detection. Biosens. Bioelectron. 2021, 179, 113076. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, H.; Cheng, J.H. DNA, protein and aptamer-based methods for seafood allergens detection: Principles, comparisons and updated applications. Crit. Rev. Food Sci. Nutr. 2023, 63, 178–191. [Google Scholar] [CrossRef]
- Piñeiro, C.; Vázquez, J.; Marina, A.I.; Barros-Velázquez, J.; Gallardo, J.M. Characterization and partial sequencing of species-specific sarcoplasmic polypeptides from commercial hake species by mass spectrometry following two-dimensional electrophoresis. Electrophoresis 2001, 22, 1545–1552. [Google Scholar] [CrossRef]
- Elsayed, S.; Bennich, H. The primary structure of allergen M from cod. Scand. J. Immunol. 1975, 4, 203–208. [Google Scholar] [CrossRef]
- Carrera, M.; Canas, B.; Vázquez, J.; Gallardo, J.M. Extensive de novo sequencing of new parvalbumin isoforms using a novel combination of bottom-up proteomics, accurate molecular mass measurement by FTICR− MS, and selected MS/MS Ion monitoring. J. Proteome Res. 2010, 9, 4393–4406. [Google Scholar] [CrossRef]
- Rehbein, H.; Kundiger, R.; Pineiro, C.; Perez-Martin, R.I. Fish muscle parvalbumins as marker proteins for native and urea isoelectric focusing. Electrophoresis 2000, 21, 1458–1463. [Google Scholar] [CrossRef]
- Etienne, M.; Jerome, M.; Fleurence, J.; Rehbein, H.; Kundiger, R.; Malmheden Yman, I.; Ferm, M.; Craig, A.; Mackie, I.; Jessen, F.; et al. A standardised method of identification of raw and heat-processed fish by urea isoelectric focusing: A collaborative study. Electrophoresis 1999, 20, 1923–1933. [Google Scholar] [CrossRef]
- Dobrovolov, I.; Ivanova, P.; Tsekov, A. Genetic-biochemical identification of some sturgeons and their hybrids (Pisces, Acipenseridae). Verh. Int. Ver. Theor. Angew. Limnol. 2005, 29, 917–921. [Google Scholar] [CrossRef]
- Rehbein, H.; Lopata, A.L. Presence of parvalbumin in different tissues of three sturgeon species (Acipenser baeri, A. gueldenstaedtii, A. ruthenus). J. Appl. Ichthyol. 2011, 27, 219–225. [Google Scholar] [CrossRef]
- Rehbein, H.; Oliveira, A. Alaskan flatfishes on the German market: Part 1: Identification by DNA and protein analytical methods. Eur. Food Res. Technol. 2012, 234, 245–251. [Google Scholar] [CrossRef]
- Mazzeo, M.F.; Giulio, B.D.; Guerriero, G.; Ciarcia, G.; Malorni, A.; Russo, G.L.; Siciliano, R.A. Fish authentication by MALDI-TOF mass spectrometry. J. Agric. Food Chem. 2008, 56, 11071–11076. [Google Scholar] [CrossRef] [PubMed]
- Carrera, M.; Cañas, B.; Gallardo, J.M. Rapid direct detection of the major fish allergen, parvalbumin, by selected MS/MS ion monitoring mass spectrometry. J. Proteom. 2012, 75, 3211–3220. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Pawar, R.; Wang, G.; Lin, H. Development of an optimized protein chip for the detection of fish parvalbumin allergen. Curr. Anal. Chem. 2011, 7, 349–356. [Google Scholar] [CrossRef]
- Mazzeo, M.F.; Siciliano, R.A. Proteomics for the authentication of fish species. J. Proteom. 2016, 147, 119–124. [Google Scholar] [CrossRef]
- Carrera, M.; Cañas, B.; Gallardo, J.M. Proteomics for the assessment of quality and safety of fishery products. Food Res. Int. 2013, 54, 972–979. [Google Scholar] [CrossRef]
- Carrera, M.; Cañas, B.; Piñeiro, C.; Vázquez, J.; Gallardo, J.M. Identification of commercial hake and grenadier species by proteomic analysis of the parvalbumin fraction. Proteomics 2006, 6, 5278–5287. [Google Scholar] [CrossRef]
- Siciliano, R.A.; d’Esposito, D.; Mazzeo, M.F. Food authentication by MALDI MS: MALDI-TOF MS analysis of fish species. In Advances in MALDI and Laser-Induced Soft Ionization Mass Spectrometry; Springer: Berlin/Heidelberg, Germany, 2016; pp. 263–277. [Google Scholar]
- Dong, X.; Raghavan, V. A comprehensive overview of emerging processing techniques and detection methods for seafood allergens. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3540–3557. [Google Scholar] [CrossRef]
- Xu, J.; Ye, Y.; Ji, J.; Sun, J.; Sun, X. Advances on the rapid and multiplex detection methods of food allergens. Crit. Rev. Food Sci. Nutr. 2022, 62, 6887–6907. [Google Scholar] [CrossRef] [PubMed]
- Rychert, J. Benefits and limitations of MALDI-TOF mass spectrometry for the identification of microorganisms. J. Infect. Epidemiol. 2019, 2. [Google Scholar] [CrossRef]
- Holzhauser, T.; Röder, M. Polymerase chain reaction (PCR) methods for detecting allergens in foods. In Handbook of Food Allergen Detection and Control; Elsevier: Amsterdam, The Netherlands, 2015; pp. 245–263. [Google Scholar]
- Liu, Z.J.; Cordes, J.F. DNA marker technologies and their applications in aquaculture genetics. Aquaculture 2004, 238, 1–37. [Google Scholar] [CrossRef]
- Broeders, S.R.; De Keersmaecker, S.C.; Roosens, N.H. How to deal with the upcoming challenges in GMO detection in food and feed. J. Biotechnol. Biomed. 2012, 2012, 402418. [Google Scholar] [CrossRef] [PubMed]
- Eischeid, A.C.; Kim, B.H.; Kasko, S.M. Two quantitative real-time PCR assays for the detection of penaeid shrimp and blue crab, crustacean shellfish allergens. J. Agric. Food Chem. 2013, 61, 5669–5674. [Google Scholar] [CrossRef] [PubMed]
- Herrero, B.; Vieites, J.M.; Espiñeira, M. Development of an in-house fast real-time PCR method for detection of fish allergen in foods and comparison with a commercial kit. Food Chem. 2014, 151, 415–420. [Google Scholar] [CrossRef]
- Cline, S.D. Mitochondrial DNA damage and its consequences for mitochondrial gene expression. Biochim. Biophys. Acta Gene Regul. Mech. BBA 2012, 1819, 979–991. [Google Scholar] [CrossRef]
- Raupach, M.J.; Barco, A.; Steinke, D.; Beermann, J.; Laakmann, S.; Mohrbeck, I.; Neumann, H.; Kihara, T.C.; Pointner, K.; Radulovici, A.; et al. The application of DNA barcodes for the identification of marine crustaceans from the North Sea and adjacent regions. PLoS One 2015, 10, e0139421. [Google Scholar] [CrossRef]
- Bremer, J.R.A.; Viñas, J.; Mejuto, J.; Ely, B.; Pla, C. Comparative phylogeography of Atlantic bluefin tuna and swordfish: The combined effects of vicariance, secondary contact, introgression, and population expansion on the regional phylogenies of two highly migratory pelagic fishes. Mol. Phylogenet. Evol. 2005, 36, 169–187. [Google Scholar] [CrossRef]
- Buhay, J.E. “COI-Like” sequences are becoming problematic in molecular systematic and DNA barcoding studies. J. Crustac. Biol. 2009, 29, 96–110. [Google Scholar] [CrossRef]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [PubMed]
- Mariani, S.; Bekkevold, D. The nuclear genome: Neutral and adaptive markers in fisheries science. In Stock Identification Methods; Academic Press: Cambridge, MA, USA, 2014; pp. 297–327. [Google Scholar]
- Paracchini, V.; Petrillo, M.; Lievens, A.; Gallardo, A.P.; Martinsohn, J.T.; Hofherr, J.; Maquet, A.; Silva, A.P.; Kagkli, D.M.; Querci, M.; et al. Novel nuclear barcode regions for the identification of flatfish species. Food Control 2017, 79, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, H.; Kress, G. Detection of short mRNA sequences in fishery products. Dtsch. Lebensm. Rundsch. 2005, 101, 333–337. [Google Scholar]
- Rehbein, H. Differentiation of hake species by RFLP-and SSCP-analysis of PCR amplified cytochrome b and parvalbumin sequences. Dtsch. Lebensm. Rundsch. 2007, 103, 511–517. [Google Scholar]
- Rencova, E.; Kostelnikova, D.; Tremlova, B. Detection of allergenic parvalbumin of Atlantic and Pacific herrings in fish products by PCR. Food Addit. Contam. Part A 2013, 30, 1679–1683. [Google Scholar] [CrossRef]
- Ma, Y.; Griesmeier, U.; Susani, M.; Radauer, C.; Briza, P.; Erler, A.; Bublin, M.; Alessandri, S.; Himly, M.; Vàzquez-Cortés, S.; et al. Comparison of natural and recombinant forms of the major fish allergen parvalbumin from cod and carp. Mol. Nutr. Food Res. 2008, 52, S196–S207. [Google Scholar] [CrossRef]
- Mukherjee, S.; Hanak, P.; Akhatova, D.; Musilova, Z.; Horka, P.; Lerch, Z.; Zdenkova, K.; Cermakova, E. Simultaneous detection and quantification of two European anglerfishes by novel genomic primer. J. Food Compos. Anal. 2022, 115, 104992. [Google Scholar] [CrossRef]
- Sun, L.; Xu, L.; Huang, Y.; Lin, H.; Ahmed, I.; Li, Z. Identification and comparison of allergenicity of native and recombinant fish major allergen parvalbumins from Japanese flounder (Paralichthys olivaceus). Food Funct. 2019, 10, 6615–6623. [Google Scholar] [CrossRef]
- Muñoz-Colmenero, M.; Rahman, S.; Martínez, J.L.; Garcia-Vazquez, E. High variability in parvalbumin beta 1 genes offers new molecular options for controlling the mislabeling in commercial Salmonids. Eur. Food Res. Technol. 2019, 245, 1685–1694. [Google Scholar] [CrossRef]
- Choi, K.Y.; Hong, K.W. Genomic DNA sequence of mackerel parvalbumin and a PCR test for rapid detection of allergenic mackerel ingredients in food. Food Sci. Biotechnol. 2007, 16, 67–70. [Google Scholar]
- Prado, M.; Boix, A.; von Holst, C. Development of a real-time PCR method for the simultaneous detection of mackerel and horse mackerel. Food Control 2013, 34, 19–23. [Google Scholar] [CrossRef]
- Heizmann, W.; Hauptle, M.; Eppenberger, H.M. The purification, characterisation and localisation of a parvalbumin like protein from chicken-leg muscle. Eur. J. Biochem. 1977, 80, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Gerday, C.; Joris, B.; Gerardin-Otthiers, N.; Collin, S.; Hamoir, G. Parvalbumins from the lungfish (Protopterus dolloi). Biochemistry 1979, 61, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Hamoir, G.; Gerardin-Otthiers, N. Differentiation of the sarcoplasmic proteins of white, yellowish and cardiac muscles of an antarctic haemoglobin-free fish Champsocephalus gunnari. Comp. Biochem. Physiol. 1980, 65, 199–206. [Google Scholar]
- Hildebrandt, S. Multiplexed identification of different fish species by detection of parvalbumin, a common fish allergen gene: A DNA application of multi-analyte profiling (xMAP™) technology. Anal. Bioanal. Chem. 2010, 397, 1787–1796. [Google Scholar] [CrossRef]
- Lu, Y.; Ohshima, T.; Ushio, H. Rapid detection of fish major allergen parvalbumin by surface plasmon resonance biosensor. J. Food Sci. 2004, 69, C652–C658. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Zhou, J.; Qi, Q.; Fu, L. A colorimetric and fluorescent gold nanoparticle-based dual-mode aptasensor for parvalbumin detection. Microchem. J. 2020, 159, 105413. [Google Scholar] [CrossRef]
- Fu, L.; Qian, Y.; Zhou, J.; Zheng, L.; Wang, Y. Fluorescence-based quantitative platform for ultrasensitive food allergen detection: From immunoassays to DNA sensors. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3343–3364. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukherjee, S.; Horka, P.; Zdenkova, K.; Cermakova, E. Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker. Genes 2023, 14, 223. https://doi.org/10.3390/genes14010223
Mukherjee S, Horka P, Zdenkova K, Cermakova E. Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker. Genes. 2023; 14(1):223. https://doi.org/10.3390/genes14010223
Chicago/Turabian StyleMukherjee, Subham, Petra Horka, Kamila Zdenkova, and Eliska Cermakova. 2023. "Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker" Genes 14, no. 1: 223. https://doi.org/10.3390/genes14010223
APA StyleMukherjee, S., Horka, P., Zdenkova, K., & Cermakova, E. (2023). Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker. Genes, 14(1), 223. https://doi.org/10.3390/genes14010223