
Lasp1 Expression Is Implicated in Embryonic Development of Zebrafish
 ,
,  , , , , and
, , , , and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Zebrafish Maintenance
2.2. Protein Extraction and Western blotting
2.3. Whole-Mount Immunofluorescence
2.4. Fluorescent Immunohistochemistry and Hematoxylin and Eosin Staining
2.5. Morpholino Injection
2.6. RNA Isolation and Droplet Digital PCR (ddPCR)
2.7. In Vitro mRNA Synthesis for the Rescue Experiments
2.8. Acridine Orange Assay for Apoptosis Evaluation
2.9. Statistical Analysis
3. Results
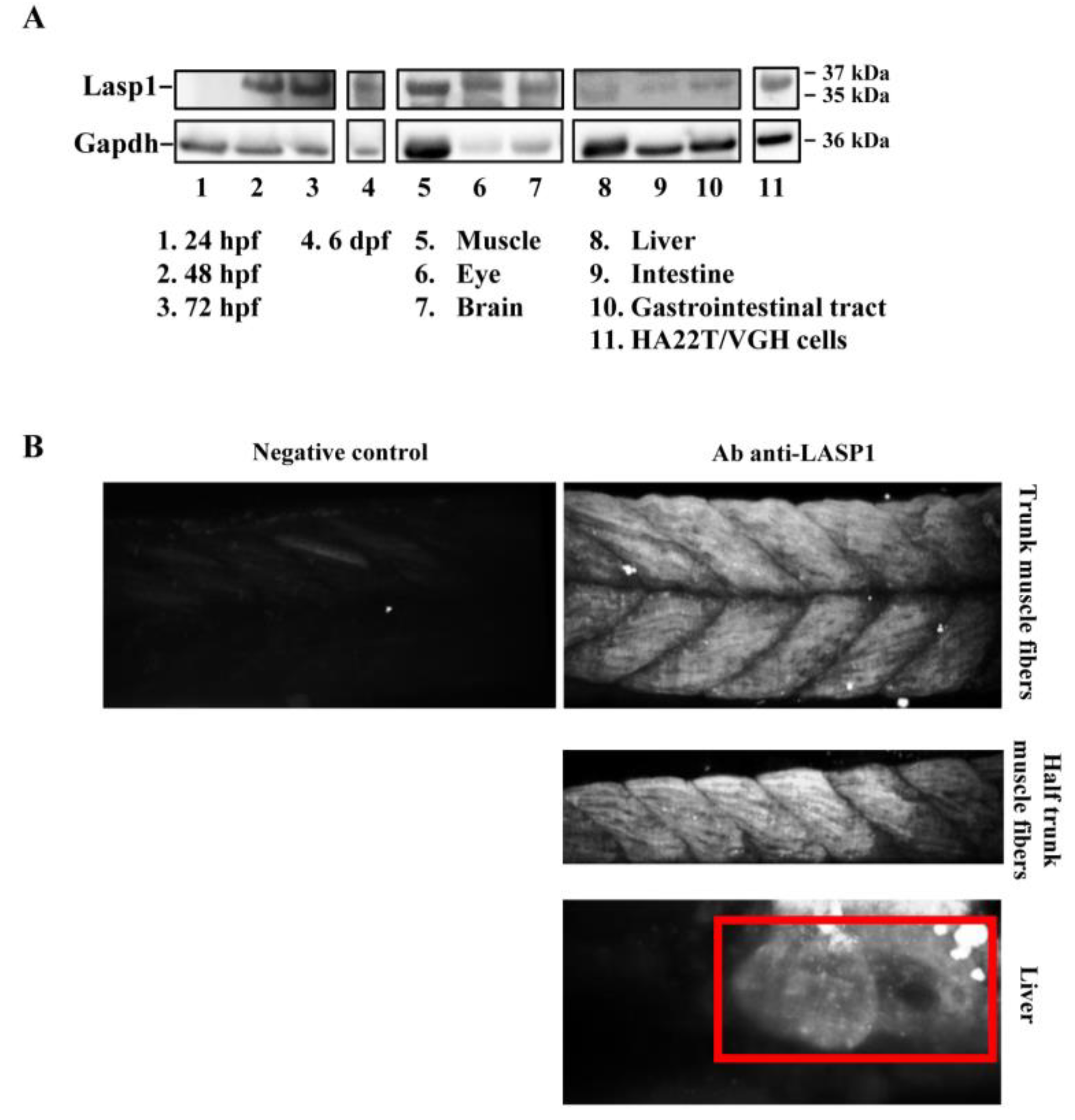
3.1. Lasp1 Protein Is Expressed at Various Stages of Zebrafish Embryonic Development and in Several of Its Adult Organs
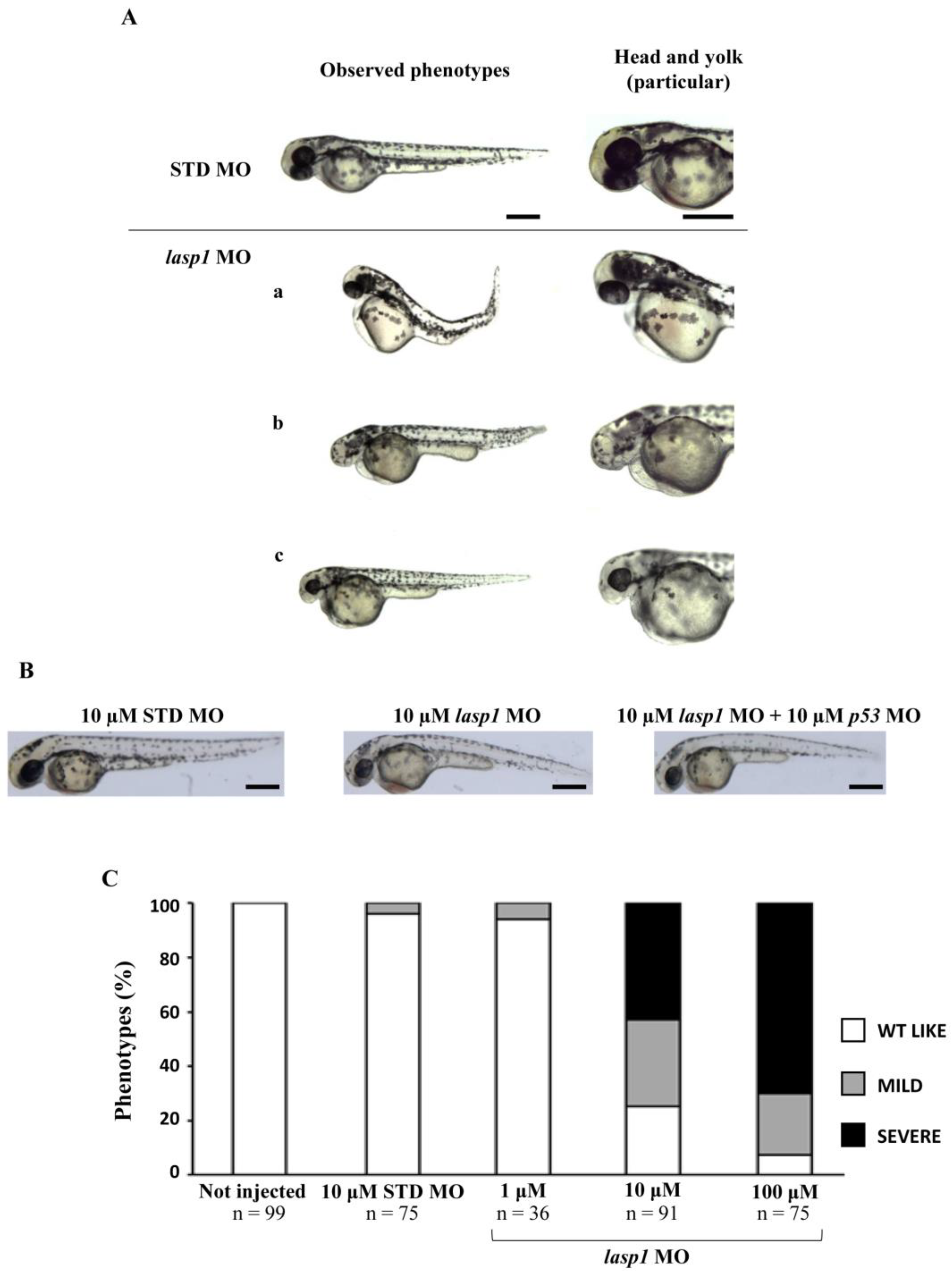
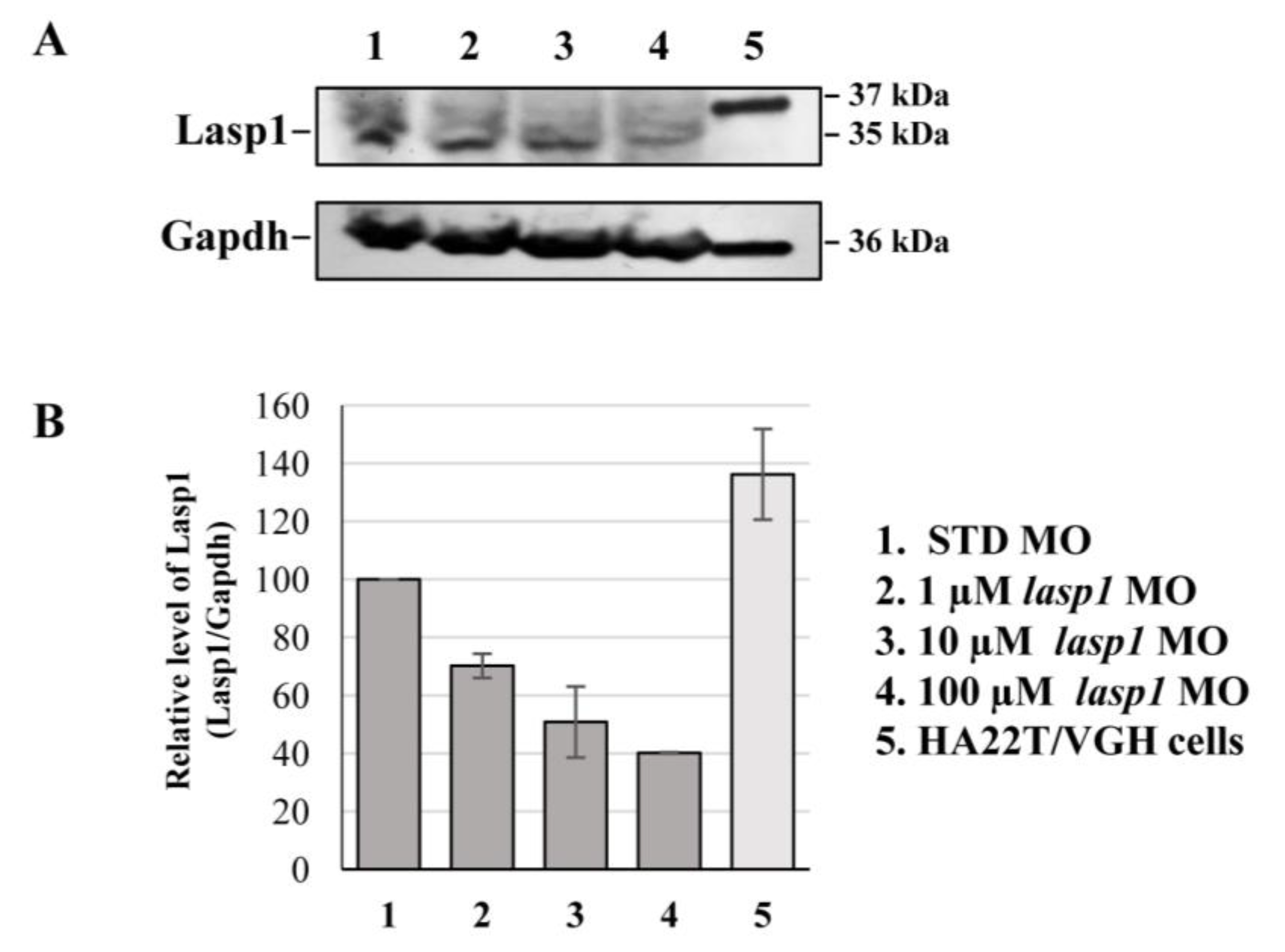
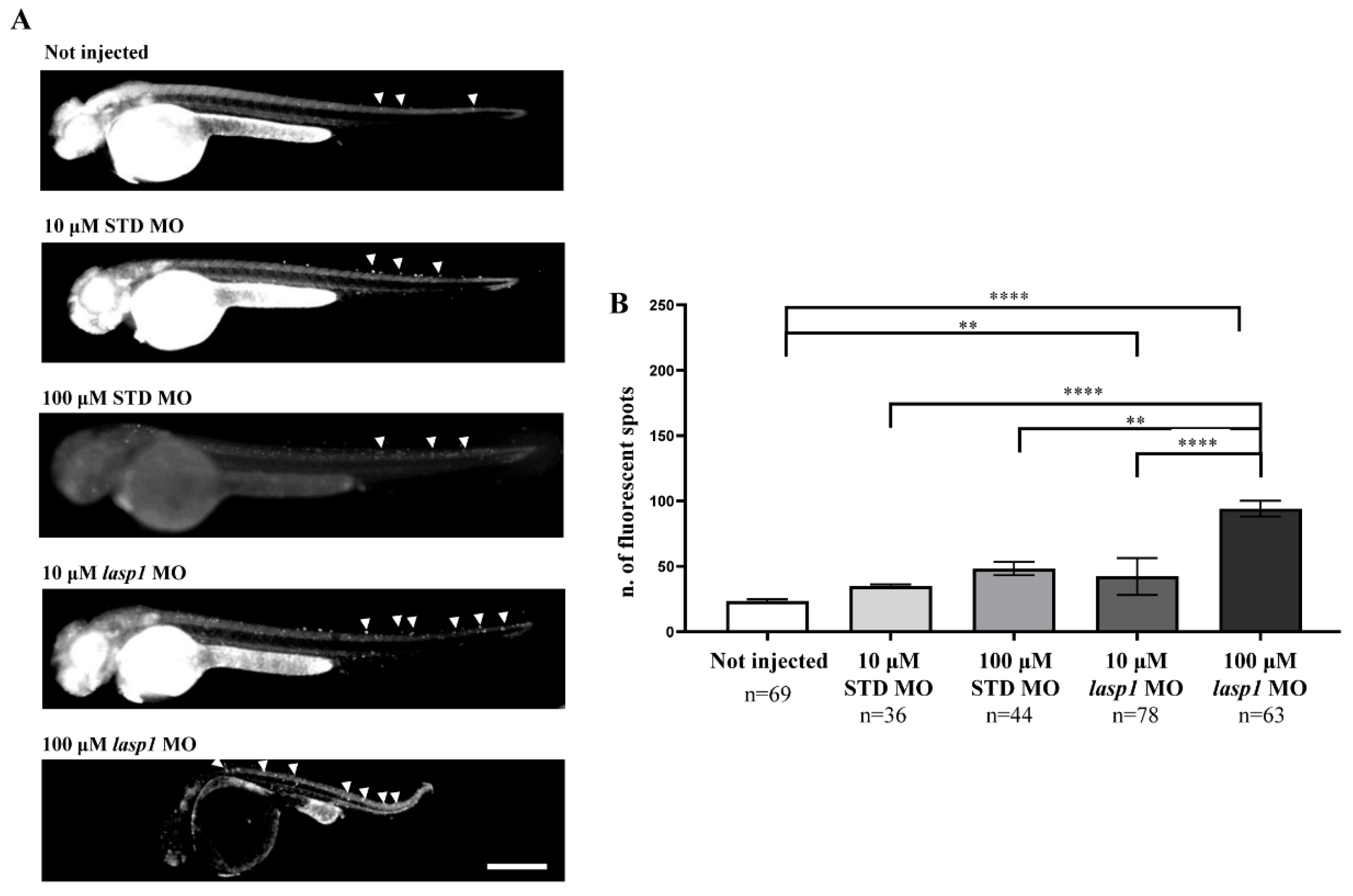
3.2. Silencing of Lasp1 by Morpholino Leads to Morphology Alterations in Zebrafish Morphants
3.3. Rescue of Zebrafish Phenotypes and Lasp1 Expression by Injection of LASP1 mRNA into the Zebrafish Embryos
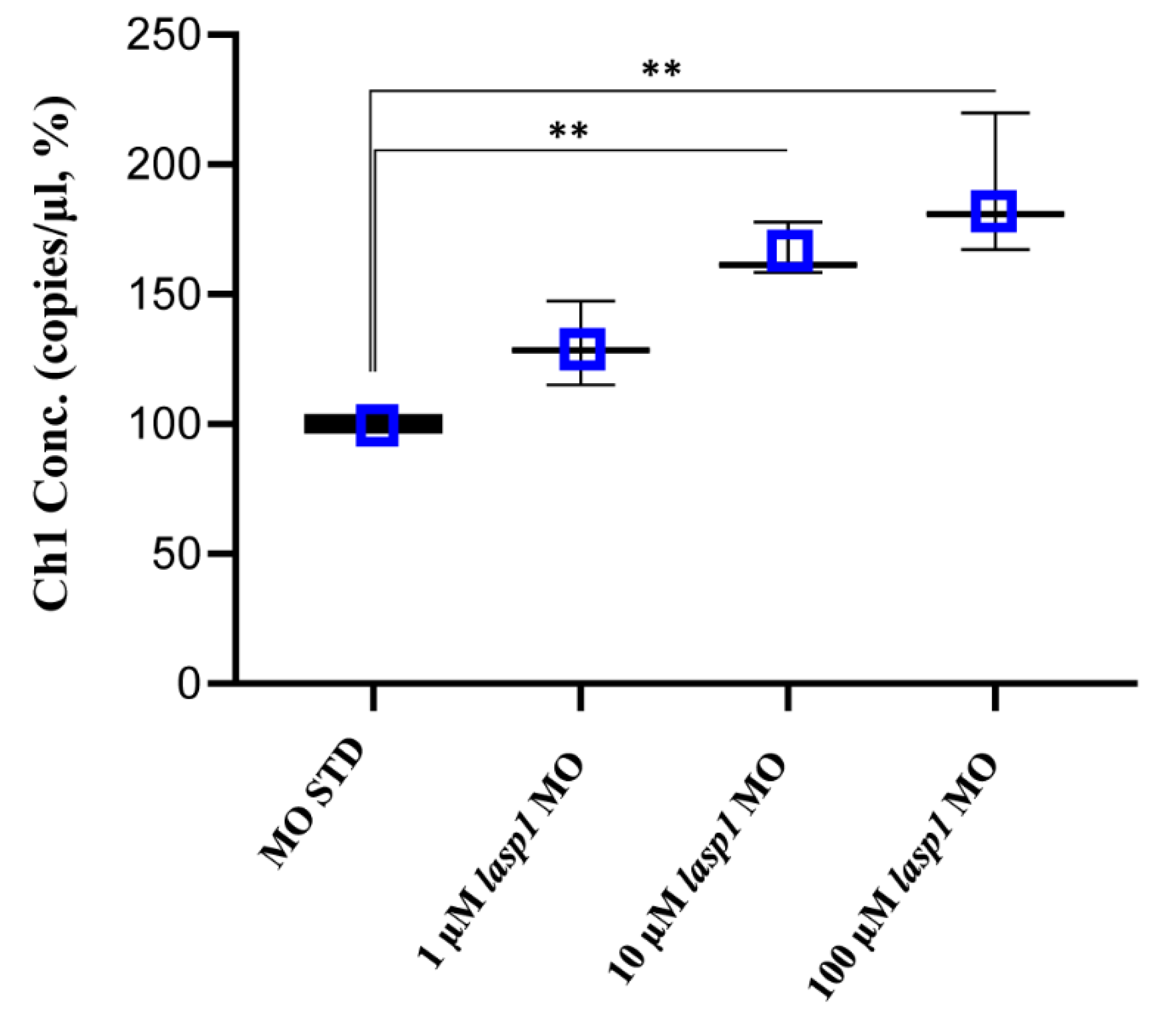
3.4. Lasp1 Silencing by MO Leads to an Increase in Apoptosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tomasetto, C.; Moog-Lutz, C.; Régnier, C.H.; Schreiber, V.; Basset, P.; Rio, M.C. Lasp-1 (MLN 50) Defines a New LIM Protein Subfamily Characterized by the Association of LIM and SH3 Domains. FEBS Lett. 1995, 373, 245–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, T.G.P.; Kammerer, U.; Winkler, C.; Schindler, D.; Sickmann, A.; Honig, A.; Butt, E. Overexpression of LASP-1 Mediates Migration and Proliferation of Human Ovarian Cancer Cells and Influences Zyxin Localisation. Br. J. Cancer 2007, 96, 296–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traenka, C.; Remke, M.; Korshunov, A.; Bender, S.; Hielscher, T.; Northcott, P.A.; Witt, H.; Ryzhova, M.; Felsberg, J.; Benner, A.; et al. Role of LIM and SH3 Protein 1 (LASP1) in the Metastatic Dissemination of Medulloblastoma. Cancer Res. 2010, 70, 8003–8014. [Google Scholar] [CrossRef] [Green Version]
- Payton, S. Bladder Cancer: LASP-1-a Promising Urine Marker for Detection of Bladder Cancer. Nat. Rev. Urol. 2012, 9, 240. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, W.; Jin, X.; Cui, S.; Zhao, L. LIM and SH3 Protein 1, a Promoter of Cell Proliferation and Migration, Is a Novel Independent Prognostic Indicator in Hepatocellular Carcinoma. Eur. J. Cancer 2013, 49, 974–983. [Google Scholar] [CrossRef]
- Shimizu, F.; Shiiba, M.; Ogawara, K.; Kimura, R.; Minakawa, Y.; Baba, T.; Yokota, S.; Nakashima, D.; Higo, M.; Kasamatsu, A.; et al. Overexpression of LIM and SH3 Protein 1 Leading to Accelerated G2/M Phase Transition Contributes to Enhanced Tumourigenesis in Oral Cancer. PLoS ONE 2013, 8, e83187. [Google Scholar] [CrossRef] [Green Version]
- Hailer, A.; Grunewald, T.G.P.; Orth, M.; Reiss, C.; Kneitz, B.; Spahn, M.; Butt, E. Loss of Tumor Suppressor Mir-203 Mediates Overexpression of LIM and SH3 Protein 1 (LASP1) in High-Risk Prostate Cancer Thereby Increasing Cell Proliferation and Migration. Oncotarget 2014, 5, 4144–4153. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, T.G.P.; Kammerer, U.; Schulze, E.; Schindler, D.; Honig, A.; Zimmer, M.; Butt, E. Silencing of LASP-1 Influences Zyxin Localization, Inhibits Proliferation and Reduces Migration in Breast Cancer Cells. Exp. Cell Res. 2006, 312, 974–982. [Google Scholar] [CrossRef]
- Salvi, A.; Bongarzone, I.; Ferrari, L.; Abeni, E.; Arici, B.; De Bortoli, M.; Scuri, S.; Bonini, D.; Grossi, I.; Benetti, A.A.A.; et al. Molecular Characterization of LASP-1 Expression Reveals Vimentin as Its New Partner in Human Hepatocellular Carcinoma Cells. Int. J. Oncol. 2015, 46, 1901–1912. [Google Scholar] [CrossRef] [Green Version]
- Butt, E.; Raman, D. New Frontiers for the Cytoskeletal Protein LASP1. Front. Oncol. 2018, 8, 391. [Google Scholar] [CrossRef]
- Orth, M.F.; Cazes, A.; Butt, E.; Grunewald, T.G.P. An Update on the LIM and SH3 Domain Protein 1 (LASP1): A Versatile Structural, Signaling, and Biomarker Protein. Oncotarget 2015, 6, 26–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollitt, S.L.; Myers, K.R.; Yoo, J.; Zheng, J.Q. LIM and SH3 Protein 1 Localizes to the Leading Edge of Protruding Lamellipodia and Regulates Axon Development. Mol. Biol. Cell 2020, 31, 2718–2732. [Google Scholar] [CrossRef] [PubMed]
- Casari, A.; Schiavone, M.; Facchinello, N.; Vettori, A.; Meyer, D.; Tiso, N.; Moro, E.; Argenton, F. A Smad3 Transgenic Reporter Reveals TGF-β Control of Zebrafish Spinal Cord Development. Dev. Biol. 2014, 396, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Grossi, I.; Arici, B.; Portolani, N.; De Petro, G.; Salvi, A. Clinical and Biological Significance of MiR-23b and MiR-193a in Human Hepatocellular Carcinoma. Oncotarget 2017, 8, 6955–6969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgio, V.; Burchell, V.; Schiavone, M.; Bassot, C.; Minervini, G.; Petronilli, V.; Argenton, F.; Forte, M.; Tosatto, S.; Lippe, G.; et al. Ca2+ Binding to F-ATP Synthase β Subunit Triggers the Mitochondrial Permeability Transition. EMBO Rep. 2017, 18, 1065–1076. [Google Scholar] [CrossRef]
- Frohlich, J.; Kovacovicova, K.; Raffaele, M.; Virglova, T.; Cizkova, E.; Kucera, J.; Bienertova-Vasku, J.; Wabitsch, M.; Peyrou, M.; Bonomini, F.; et al. GDF11 Inhibits Adipogenesis and Improves Mature Adipocytes Metabolic Function via WNT/β-Catenin and ALK5/SMAD2/3 Pathways. Cell Prolif. 2022, 55, e13310. [Google Scholar] [CrossRef]
- Schiavone, M.; Rampazzo, E.; Casari, A.; Battilana, G.; Persano, L.; Moro, E.; Liu, S.; Leach, S.D.; Tiso, N.; Argenton, F. Zebrafish Reporter Lines Reveal in Vivo Signaling Pathway Activities Involved in Pancreatic Cancer. DMM Dis. Model. Mech. 2014, 7, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Manganelli, M.; Grossi, I.; Ferracin, M.; Guerriero, P.; Negrini, M.; Ghidini, M.; Senti, C.; Ratti, M.; Pizzo, C.; Passalacqua, R.; et al. Longitudinal Circulating Levels of Mir-23b-3p, Mir-126-3p and Lncrna Gas5 in Hcc Patients Treated with Sorafenib. Biomedicines 2021, 9, 813. [Google Scholar] [CrossRef]
- Manganelli, M.; Grossi, I.; Corsi, J.; D’Agostino, V.G.; Jurikova, K.; Cusanelli, E.; Molfino, S.; Portolani, N.; Salvi, A.; De Petro, G. Expression of Cellular and Extracellular TERRA, TERC and TERT in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 6183. [Google Scholar] [CrossRef]
- Robu, M.E.; Larson, J.D.; Nasevicius, A.; Beiraghi, S.; Brenner, C.; Farber, S.A.; Ekker, S.C. P53 Activation by Knockdown Technologies. PLoS Genet. 2007, 3, 787–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerety, S.S.; Wilkinson, D.G. Morpholino Artifacts Provide Pitfalls and Reveal a Novel Role for Pro-Apoptotic Genes in Hindbrain Boundary Development. Dev. Biol. 2011, 350, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, J.S.; Smith, J.C. Controlling Morpholino Experiments: Don’t Stop Making Antisense. Development 2008, 135, 1735–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, C.; Chen, Y.; Tao, B.; Peng, L.; Peng, T.; Yang, X.; Xia, X.; Chen, L. LIM and SH3 Protein 1 Regulates Cell Growth and Chemosensitivity of Human Glioblastoma via the PI3K/AKT Pathway. BMC Cancer 2018, 18, 722. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Tang, L.; Wu, L.; Li, K.; Wang, H.; Li, W.; Wu, J.; Li, M.; Wang, S.; Zhao, L. LASP1 Promotes Nasopharyngeal Carcinoma Progression through Negatively Regulation of the Tumor Suppressor PTEN. Cell Death Dis. 2018, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Guo, J.; Li, X. Role of LASP-1, a Novel SOX9 Transcriptional Target, in the Progression of Lung Cancer. Int. J. Oncol. 2018, 52, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.L.; Wang, L.; Wang, X.Y.; Shang, D.; Yin, S.J.; Sun, L.L.; Ji, H.B. MicroRNA-218 Inhibits the Proliferation, Migration, and Invasion and Promotes Apoptosis of Gastric Cancer Cells by Targeting LASP1. Tumour Biol. 2016, 37, 15241–15252. [Google Scholar] [CrossRef]
- Huang, Z.; Pang, G.; Huang, Y.G.; Li, C. MiR-133 Inhibits Proliferation and Promotes Apoptosis by Targeting LASP1 in Lupus Nephritis. Exp. Mol. Pathol. 2020, 114, 104384. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Fulton, R.S.; McLellan, M.D.; Schmidt, H.; Kalicki-Veizer, J.; McMichael, J.F.; Fulton, L.L.; Dooling, D.J.; Ding, L.; Mardis, E.R.; et al. Comprehensive Molecular Portraits of Human Breast Tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Nasevicius, A.; Ekker, S.C. Effective Targeted Gene “knockdown” in Zebrafish. Nat. Genet. 2000, 26, 216–220. [Google Scholar] [CrossRef]
- Ekker, S.C.; Larson, J.D. Morphant Technology in Model Developmental Systems. Genesis 2001, 30, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stainier, D.Y.R.; Raz, E.; Lawson, N.D.; Ekker, S.C.; Burdine, R.D.; Eisen, J.S.; Ingham, P.W.; Schulte-Merker, S.; Yelon, D.; Weinstein, B.M.; et al. Guidelines for Morpholino Use in Zebrafish. PLOS Genet. 2017, 13, e1007000. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grossi, I.; Schiavone, M.; Cannone, E.; Grejdan, O.A.; Tobia, C.; Bonomini, F.; Rezzani, R.; Salvi, A.; De Petro, G. Lasp1 Expression Is Implicated in Embryonic Development of Zebrafish. Genes 2023, 14, 35. https://doi.org/10.3390/genes14010035
Grossi I, Schiavone M, Cannone E, Grejdan OA, Tobia C, Bonomini F, Rezzani R, Salvi A, De Petro G. Lasp1 Expression Is Implicated in Embryonic Development of Zebrafish. Genes. 2023; 14(1):35. https://doi.org/10.3390/genes14010035
Chicago/Turabian StyleGrossi, Ilaria, Marco Schiavone, Elena Cannone, Oana Andreea Grejdan, Chiara Tobia, Francesca Bonomini, Rita Rezzani, Alessandro Salvi, and Giuseppina De Petro. 2023. "Lasp1 Expression Is Implicated in Embryonic Development of Zebrafish" Genes 14, no. 1: 35. https://doi.org/10.3390/genes14010035