Abstract
Ewing’s sarcoma is a rare type of cancer that forms in bones and soft tissues in the body, affecting mostly children and young adults. Current treatments for ES are limited to chemotherapy and/or radiation, followed by surgery. Recently, microRNAs have shown favourable results as latent diagnostic and prognostic biomarkers in various cancers. Furthermore, microRNAs have shown to be a good therapeutic agent due to their involvement in the dysregulation of various molecular pathways linked to tumour progression, invasion, angiogenesis, and metastasis. In this review, comprehensive data mining was employed to explore various microRNAs that might have therapeutic potential as target molecules in the treatment of ES.
1. Introduction
Ewing’s sarcoma (ES) is a rare, aggressive cancer, which particularly affects bones and soft tissues surrounding the bones [1,2,3]. ES affects any bone in the body, however, about 45% of cases have been located in the lower extremities, with another 45% in the pelvis, and 13% in the upper extremities, as well as the axial skeleton and ribs, and a further 2% in the face [4]. ES is an uncommon bone tumor that has a poor prognosis and a significant chance of spreading to the bone marrow and lung as its main sites of metastasis. ES tumors can be classified into askin tumors and peripheral primitive neuroectodermal tumors (pPNETs) based on the location of tumors in the human body. ES is considered the second death-causing bone tumour after osteosarcoma that affects children, adults, and young (less than 16 years old), respectively [5]. ES is controlled by the chromosomal translocation t(11;22) (q24;q12), which produces the fused EWSR1-FLI1 protein, a well-known transcription factor that regulates the activity of oncogenes.
The pathogenesis of ES remains unclear and there are cellular environmental factors that may possibly have an influence on the overexpression of the EWS-FLI1 complex. However, data show that this overexpression of EWS-FLI1 in primary cells causes an arrest in growth or even cell death. The differentiation defects in ES result in the enhanced expression of EWS/FLI in primitive cells that lack definitive lineage markers [6]. Another study reported that the formation of the EWS-FLI1 protein is responsible for biological alterations that cause neoplastic transformation in mesenchymal stem cells (MSCs) leading to the existence of ES [7]. However, further investigation is required to better understand the formation of the EWS-FLI1 gene and its involvement in ES. However, multiple published research reports showed that the EWS-FLI1 gene works as a transcriptional repressor and controls cellular proliferation and survival activities [6,8]. EWS-FLI1 also works as a transcriptional activator and promotes the expression of SRY-Box Transcription Factor 2 (SOX2), Insuline-like Growth Factor (IGF1), Enhancer of Zeste Homolog 2 (EZH2) genes respectively [9,10].
Around 22 percent of ES patients have metastases, and 14% have free survival rates. Additionally, the survival percentage for 5-year-old patients with no apparent metastases at diagnosis is 61% [11]. In the last few years, researchers have been trying to investigate some novel targets including genes, proteins, and microRNAs (miRs) to better understand the basic mechanism of ES [12,13,14,15]. Therefore, by knowing the significance of miRs in ES, the present review explores the miRs that could be used as therapeutic targets for ES and could be used in the designing of new chemical scaffolds and vaccines for the medication of ES.
2. Biosynthesis of miRs and Their Involvement in Cancers
miRs are small, composed of approximately 22 nucleotides, non-coding RNAs involved in switching on and off specific genes through transcriptional gene silencing (TGS) and changing their expression by post-transcriptional gene silencing (PTGS). miRs are accountable for the physiological and pathological processes related to cancer [16]. miRs are highly conserved and are transcribed by RNA polymerases II and III, respectively, which are responsible for the generation of the precursors that are then further subjected to a series of cleavages, which are nuclear and cytoplasmic in their nature. Additionally, miRs regulate at least 30% of the protein-coding genes and makeup roughly 1% to 5% of the human genome. The biogenesis of miR is a multi-step process that includes a nuclear and cytoplasmic phase as well as the participation of the ribonuclease III enzymes, dicer protein in the cytoplasm, and drosha protein in the nucleus.
The miR transcripts are first formed by RNA polymerase II and then go through a processing step, allowing miR synthesis according to specific conditions and cell types. Multiple research studies have reported that miRs are significantly implicated in stem cell pluripotency and somatic cell reprogramming [17,18,19,20]. Genes for miRs are transcribed into primary miRs, and handed to the drosha enzyme in the nucleus, where they are processed. In this processing step catalysed by Drosha, the hairpin loop of the pre-miR is cleaved at the stem and removed [21] (Figure 1).
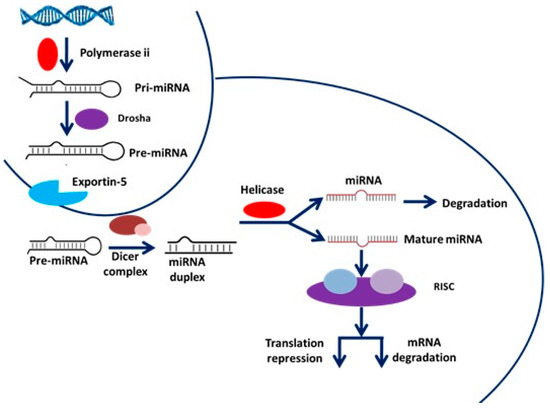
Figure 1.
The basic mechanism of miR biogenesis [22].
Numerous published data have shown the significant role of miRs in various diseases such as cancer, Alzheimer‘s, cardiovascular, and skeletal muscle hypertrophy [23,24,25,26]. The prior reports showed that miRs control the expression of various genes by binding with messenger RNA (mRNA) in the cell [27]. Such bound mRNA stops coding for a specific protein and is destroyed [28]. A miR associated with cancer is called oncomir (or oncomiR). Additionally, many studies described the functions of miRs in cancer biology as biomarkers and new therapeutic targets against cancer [29,30,31].
The deletion of the miR-15/-16 cluster is associated with increased expression of a leukemia-related anti-apoptotic target gene [32]. Moreover, different studies reported that miRs are linked with tumour biologies such as angiogenesis, metastasis, and cell death [33,34]. There are multiple reports which explore the functionality of miRs in promoting cell invasion and metastasis [35,36] along with tumour suppressors by inhibiting Epithelial Mesenchymal Transition (EMT) and metastasis [37,38]. Furthermore, it has also been found that miRs are involved in Cancer Stem Cells (CSCs) biology by targeting various transcription factors such as Oct4, Sox2, Lin28A, and KLF4, respectively [39,40,41,42]. Recent studies have shown that miRs are associated with chemoresistance and chemosensitivity in various cancers. Another study found that the overexpression of miR-105/93-3p activates the Wnt/β-catenin signalling pathway by activating SFRP1, leading to a promotion of chemoresistance in breast cancer [43].
3. miRs in ES Development and Progression
There are multiple studies suggesting a possible association of miRs with ES. The functional study of miR-30a showed a functional correlation between EWS-FLI1 and CD99 membrane glycoprotein which are two significant biomarkers and therapeutic targets for ES [44]. Moreover, there is also a report that miR-20b, miR-130b, and miR-34b act as oncomiRs (miRs associated with cancer) in ES progression [9]. According to Kawano et al. (2017), the modulation of Transforming Growth Factor β Receptor II (TGFBR2) by miRs contributes significantly to the advancement of ES. They observed an antagonistic association between the downregulation of TGFBR2 and the overexpression of miR-20b in humans. This clearly indicates the oncomiRic character of miR-20b and its potential to regulate the TGFRB2 pathway [45].
miR-130b has a direct association with ES metastasis and found that its overexpression leads to enhanced cellular invasion and migration ability [46]. Additionally, the molecular evidence demonstrates that miR-130b promotes the Cell Division Cycle 42 (CDC42) pathway by suppressing Rho GTPase Activating Protein 1 (ARHGAP1), which results in the constitutive activation of CDC42. The activation of P21 (RAC1) Activated Kinase 1 (PAK1), which positively targets gene expression and is responsible for sarcoma invasion and metastasis and creates a positive feedback loop by promoting miR-130b expression, is furthermore caused by the knockdown of ARHGAP1 gene expression and prolonged activation of CDC42 [46]. miR-34b is also involved in promoting proliferation, migration, and invasion of Ewing’s sarcoma cells by decreasing the levels of mRNA and Notch homolog protein 1 (Notch 1), respectively [47]. The prior data indicated that the Notch signalling pathway can act as an oncogene in human cancers [48]. Another miR linked with ES is miR-708-5p through a differential expression pattern. miR-708-5p is downregulated in ES cells and its overexpression is correlated with reduced proliferation of various tumours [49,50]. The overexpression of miR-708-5p suppresses the tumor invasiveness and migration, thanks to positively targeting Rho-GTPase Rho C effectors (ROCK1 and ROCK2) and Matrix Metallopeptidase 12 (MMP-12), respectively [51]. On the contrary, another oncomiR miR-181c acts as an ES tumor promotor by negatively targeting the Tumour Necrosis Factor Receptor Super Family Member 6 (TNFRSF6) in ES cells [52].
One prior report showed that both miR-193b and miR-124 can act as tumour suppressors and specifically, miR-193b has been shown its associated with tumorigenesis and inhibit tumour growth and metastasis [53]. The functional data reported that miR-124 target Zinc Finger Protein SNAI2 (SLUG), which regulate cellular motility, inducing epithelial to mesenchymal transition (EMT) and overexpression of the cell cycle Cyclin D2 regulator (encoded by the CCND2 gene). Moreover, miR-124 overexpression inhibits cell motility and block the cell cycle progression [54]. The miR-193 negatively correlated with receptor tyrosine-protein kinase erbB-4 (ErbB4 gene), which has been identified as a metastasis promoter in ES [55,56]. Similarly, miR-30d is associated with growth, malignant progression, and apoptosis in ES. The miR-30d is also considered a tumor suppressor and its overexpression is linked with poor prognosis in prostate cancer [57,58]. The mechanistic study also investigated how downregulating the production of matrix metalloproteinases-9 and -2 (MMP-9/-2) in ES cell lines led to an increase in miR-30d in those cells.
Additionally, miR-30d overexpression results in the accumulation of apoptotic cells, which increases the expression of the Bcl-2-like protein 4 (Bax) and concurrently decreases the levels of the B-cell lymphoma 2 (Bcl-2) proteins. Furthermore, the MEK/ERK and PI3K/Akt signalling pathways, which are involved in cell survival, proliferation, cell cycle progression, metabolism, and protein expression in ES cells, have been connected to miR-30d and other key signalling pathways in the evolution of tumours [59,60]. The functional study reported that miR-107 overexpression is correlated with cell cycle progression, apoptosis, and restrained cell cycle arrest. The molecular mechanism behind these effects is probably mediated by the negative target of miR-107 on the Hypoxia-Inducible factor-1β (HIF-1β) gene [61]. Another miR-185 is also known as oncomiR due to its low expression in ES tissues and cells and tumour suppressor activity in various cancers [62,63]. The prior study explores the role of miR-185 in ES progression by its overexpression in ES cell lines which reduces cell proliferation and colony formation ability through Bcl-2 and the E2F Transcription Factor 6 (E2F6) targets [64].
In ES, miRs such as miR-683 and miR-107 prevent tumor progression and cell proliferation. The ES cells demonstrated that miR-638 is downregulated, and its overexpression inhibits vascular endothelial growth factor A (VEGFA) function by suppressing mRNA and VEGFA protein expression. It also induces cell death and inhibits cell proliferation. While miR-107 appears to suppress angiogenesis by inhibiting the development of new blood vessels [65]. In another bioinformatic study, it has been reported that miR-21, upon expression in ES cells, works as a tumour suppressor by targeting and downregulating the expression of Activated Leucocyte Cell Adhesion Molecule (ALCAM) [66]. Whereas the downregulation of miR-199b-5p has been reported in ES cell lines in comparison with human MSCs (Table 1) [67].

Table 1.
miRs and their possible target genes.
The miR-181c is linked with cancer development and manifested as an oncogene in ES [72]. The miR-214-3p is exhibited oncomiR behaviour and is constitutively suppressed in cell lines. However, when the EWS-FLI1 and/or CD99 genes are silenced, it is once again expressed, and miR-214-3p function as a marker of tumour aggressiveness by restricting ES cell proliferation and migration and suppressing the expression of its target HMGA1 [73].
miRs have altered the expression levels in cancer and this changed miRs expression profiles have been identified in several malignancies, particularly in osteosarcoma and EWS, respectively. It has been studied that EWS/FLI1 makes changes in miR-145 and let-7a expression profiles in EWS. When EWS-FLI1 is knocked down, the miR-145 expression increases rapidly. It is markedly decreased in EWS cell lines [74]. In contrast, miR-145 transfection raised EWS-FLI1 levels while ectopic expression of miR-145 in Ewing’s sarcoma cell lines significantly decreased EWS-FLI1 protein. Like let-7a, HMGA2 is a direct EWS-FLI-1 target implicated in the tumorigenicity of EFT cells because of its impact on the target oncogene’s expression. A signature of five miRs (miR-34a, miR-23a, miR-92a, miR-490-3p, and miR-130b) was found to be an independent predictor of risk for disease progression and survival after microarray analysis of data from 49 individuals with primary ES [75].
4. The Functional Role of Let-7 Family in ES
The members of the let-7 family, commonly known as tumour-suppressing miRs, have been involved in the downregulation of various kinds of cancers in comparison to normal, healthy tissues. There are a total of ten members of the let-7 family of miRs discovered in the human species, namely: let-7a, let-7b, let-7c, let-7d, let-7e, let-7f, let-7g, let-7i, miR-98, and miR-202, produced from approximately thirteen precursor sequences. Boyerinas et al. showed that a decrease in the let-7 family may lead to lower and poor survival rates of cancer patients [76]. The size of the family members of miRs varies, there are three basic separate precursor sequences of miRs such as let-7a-1, let-7a-2, and let-7a-3 for the production of let-7a. Similarly, let-7f is produced from two precursor miRs let-7f-1 and let-7f-2, respectively. It has been reported that five different miRs: let-7a, let-7b, let-7e, let-7f, and let-7g are directly associated with ES. Additionally, these miRs are linked with ES through the deregulation of miRs in 90% of ES patients. Furthermore, these observations were made through target gene prediction conducted of the deregulated miRNAs. A major correlation has been found through the observation of expression of let-7a and let-7b, especially a low expression of let-7b correlates with a higher risk of relapse of the disease [76]. Let-7 is known to have a crucial role in the negative regulation of oncogenic mutations of rat sarcoma virus (RAS), as well as in down or negative regulation of HIF-1α and EWS/FLI1 that are known to have an important role in the prognosis of ES. Therefore, let-7a has been reported as a direct target of EWS/FLI1 activity. The let-7a precursor sequence itself contains two mature miR sequences, 3p (containing 21 nucleotides) and 5p (containing 22 nucleotides), respectively. Another study showed that there is a link between the let-7a by EWS/FLI-1 [77]. The general mechanism through which let-7a expression controls the growth of ES is through the High Mobility Group AT-Hook 2 (HMGA2) protein-coding gene. As let-7a is overexpressed and HMGA2 in turn is repressed, both may work to block ES tumor cell tumorigenicity. Hence, there can be therapeutic use of overexpression and use of let-7a against the aggressive malignant tumour-forming mRNA and proteins, such as those in ES. The mechanistic study showed that let-7a is a direct EWS/FLI-1 target that has been evidenced to be involved in ES family tumors, this mechanistic pathway may be suggested as a possible (Figure 2). Firstly, the primary miRNA interacts with the Drosha enzyme processing complex and is converted into the precursor let-7a sequence. The Dicer enzyme further processes this precursor sequence into the mature sequence of let-7a. This mature sequence in turn may form a complex with the EWSR1 mRNA to silence this gene and disrupt the formation of the EWS protein, thus also impairing the formation of the EWS/FLI1 complex that occurs in ES [69,77]. The let-7 miRs having associations with carcinogenesis have been tabulated in Table 2.
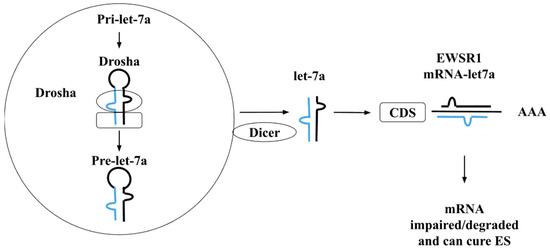
Figure 2.
Mechanistic pathway of let-7a in ES. The blue and black color represents the 5’cap and poly A tail of miRNAs and after unwinding it binds with mRNA.
Table 2.
Let-7 family in association with carcinogenesis genes.
5. miRs as Therapeutic Agents and Applications
Due to their ability to target multiple genes and cellular pathways altered in pathological conditions, miRs could be good candidates for novel therapeutic agents. miRs are involved in various diseases and play important roles due to their involvement in mutations and downregulations of key enzymes. The dysregulation of the miR biogenesis pathway has been linked in several studies to the emergence of cancer. Additionally, it has been noted that a poor prognosis is associated with the downregulation of the drosha and dicer enzymes, which has been found in numerous cancers [87,88].
ES is a paediatric sarcoma that usually originates in bones. Genetically, it arises from chromosomal translocations that fuse the EWS protein to different transcription factor members of the ETS family, with EWS-ETS fusion genes acting as the pathogenic catalyst for ES [12,89]. Additionally, some miRs could potentially target insulin-like growth factor (IGF) which is a known player for ES [12]. Another study identified miR-145 as a top miR candidate that is repressed in ES [89]. The EWS/FLI1 fusion transcript may be directly targeted by miR-145, which suggests that this regulatory network should be further investigated as a possible target for miR-mediated therapeutics in this kind of cancer. Let-7a is also identified as a direct target for the EWS/FLI1 fusion transcript [77]. The role of miRs as biomarkers in ES has been investigated [90,91]. It has been shown that patients with high levels of miR-34a have better 5-year survival outcomes than patients with low levels of miR-34a [90]. An integrated analysis study of miRs and copy number changes revealed around 20 differentially expressed miRs in chromosomal regions with altered copy numbers. There are around 35 miRs have been shown to have distinct expression patterns in ES tissues and cell lines in comparison with MSCs [92]. Furthermore, the generated results showed that miR-31 act as a tumor suppressor that affected proliferation and invasion in Ewing cell lines.
The expression levels of miR-100, miR-125b, miR-22, miR-221/222, miR-27a, and miR-29a are also suppressed upon suppression of EWS/FLI1 in ES cells. These miRs regulate the insulin-like growth factor (IGF) pathway and promote ES cell growth [12]. Moreover, miR-145 also controls the expression level of EWS/FLI1 and suppresses the ES cell growth [89]. The EWS/FLI1 fusion protein upregulates the EYA3 by suppressing miR-708 and as a result it may be considered as a drug target for the designing of novel chemical scaffolds against ES [93]. Another cell surface glycoprotein CD99 is present in ES tissue, and strongly related to ES malignancy and expression of EWS/FLI1 [94]. Expression of CD99 is regulated by miR-34a via the Notch signalling pathway and prevents ES differentiation. In addition, CD99 is also expressed in the membrane-bound extracellular vesicles, called exosomes derived from ES cells [94]. The analysis of 49 primary ES tumours revealed that miR-34a is related to ES progression and its low expression predicts poorer outcomes and overall worse survival. The IGF-1/AKT/mTOR pathway is also one of the key targets for ES treatment and IGF receptor 1 (IGF-1R) and mTOR inhibitors prevent tumour growth and improve ES survival rate in a mouse xenograft model [95].
However, besides these miRs, there are some other long non-coding RNAs (lncRNAs) that have been shown to affect various aspects of carcinogenesis mostly through modulation of the function of cancer-associated miRs [96]. Deregulation of the miRs is a frequent feature of several illnesses. The development of miR therapies for the treatment of a wide range of human illnesses is therefore the focus of many different sorts of study. The first-ever miRNA therapy is undergoing phase II clinical trials for the treatment of hepatitis C virus (HCV) infection. Miravirsen is a short-locked nucleic acid (LNA) medication targeting miR-122 that is quickly making its way from the lab to the clinic. Novel nucleic acid analogues called LNAs have some therapeutic potential. They are essentially modified or customized RNA nucleotides [97].
miRs have shown potential implications in the treatment of ES, a rare and aggressive form of bone and soft tissue cancer however, current research has explored different promising strategies and applications of miRs in ES therapy [98]. In tumor suppression, multiple miRs function as tumor suppressors by inhibiting the expression of genes involved in cancer growth and metastasis [99]. Researchers have identified miRs that are downregulated in ES and have explored the use of miR mimics to restore their levels. These miRNA mimics can potentially inhibit ES cell growth and induce apoptosis (cell death). Secondly, miRs target and downregulate oncogenes (genes that promote cancer), therefore, modulating the activity of miRs helps to suppress the expression of oncogenes that drive ES development and progression along with metastatic inhibition. Another significant application of miRs is chemotherapy sensitization; miRs modulate the sensitivity of cancer cells to chemotherapy drugs. Researchers have investigated the potential of miRs to sensitize ES cells to chemotherapy, making them more responsive to standard treatments. Most importantly, miR-based therapies also support traditional treatments such as chemotherapy and radiation therapy to achieve synergistic effects. Therefore, combinations of miR inhibitors with other drugs enhance the therapeutic response [100]. Moreover, miR profiling of ES tumors supports theidentification of specific miR expression patterns associated with disease aggressiveness and prognosis. This information can guide treatment decisions and the development of personalized therapeutic strategies against ES. It’s noteworthy that miR-based therapies hold promise for ES treatment, further research is needed to fully understand the mechanisms and optimize their use in clinical settings for ES therapy in the future.
6. Conclusions and Future Prospectus
ES causes significant mortality due to the low effectiveness of currently available treatments [101]. Despite the fact that surgical interventions have improved the prognosis, the substantial risk of local relapse makes ES treatment ineffective. Due to the lack of precise information on the molecular pathways involved in ES pathogenesis, the early identification of ES is a significant problem. Therefore, developing new therapeutic techniques for therapy necessitates a deeper understanding of ES and its pathogenic process. There has been a rapid accumulation of evidence over the past ten years supporting the crucial role that unregulated miRs play in the development of human cancers [102,103]. This creates enormous potential for innovative developments in diagnostic and therapeutic strategies in the management of malignant diseases, including ES.
Our knowledge of cancer biology has been significantly improved by the discovery of miRs as post-transcriptional regulators of gene expression, particularly for ES. Although the results are intriguing, more research is required to fully understand the biological functions of miRs and to investigate any potential clinical applications. It is challenging to find consistent miR signatures for the prognosis, diagnosis, and treatment of sarcomas since miR expression is tissue-specific and can be disrupted by a variety of other variables such as infection, hypoxia, pathology, and cytotoxic treatment. miR-directed therapies may have unfavorable side effects because miRs also function in cell signaling networks and affect the expression of several genes. There are multiple miR-based drugs under clinical trials against different diseases which open the new gateways for ES. The miR data could be used as new biomarkers and therapeutic targets to design novel chemical scaffolds for the treatment of ES. Nonetheless, the potential therapeutic utilization of miRs may reshape our understanding of tumorigenesis and will improve the management of ES in the future, potentially giving rise to miR-based medicines and vaccines that could be innovative for paediatric sarcoma therapies, particularly for ES.
Author Contributions
Conceptualization, M.H. and A.K.; data collection, S.S., A.M., S.u.D. and M.Y.; writing, original draft preparation, W.C. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NSF grant DBI 1661391, and NIH grants R01GM127701, and R01HG012117.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
A.K. acknowledges the financial support from NSF grant DBI 1661391, and NIH grants R01GM127701, and R01HG012117. M.H. acknowledges the Ohio State University for providing the “President’s Postdoctoral Scholars Program (PPSP)” award for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roundhill, E.A.; Pantziarka, P.; Liddle, D.E.; Shaw, L.A.; Albadrani, G.; Burchill, S.A. Exploiting the stemness and chemoresistance transcriptome of Ewing sarcoma to identify candidate therapeutic targets and drug-repurposing candidates. Cancers 2023, 15, 769. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Park, J.; Han, E.-T.; Park, W.S.; Han, J.-H.; Kwon, Y.-S.; Lee, H.-J.; Hassan, M.; Kloczkowski, A.; Chun, W. Exploration of Flavonoids as Lead Compounds against Ewing Sarcoma through Molecular Docking, Pharmacogenomics Analysis, and Molecular Dynamics Simulations. Molecules 2023, 28, 414. [Google Scholar] [CrossRef] [PubMed]
- Caltavituro, A.; Buonaiuto, R.; Salomone, F.; Morra, R.; Pietroluongo, E.; De Placido, P.; Tortora, M.; Peddio, A.; Picozzi, F.; Ottaviano, M. Extraskeletal Ewing’s sarcoma of the mediastinum: Case report. Front. Oncol. 2023, 13, 1074378. [Google Scholar] [CrossRef]
- Burchill, S. Ewing’s sarcoma: Diagnostic, prognostic, and therapeutic implications of molecular abnormalities. J. Clin. Pathol. 2003, 56, 96–102. [Google Scholar] [CrossRef]
- Chen, F.; Wang, S.; Wei, Y.; Wu, J.; Huang, G.; Chen, J.; Shi, J.; Xia, J. Norcantharidin modulates the miR-30a/Metadherin/AKT signaling axis to suppress proliferation and metastasis of stromal tumor cells in giant cell tumor of bone. Biomed. Pharmacother. 2018, 103, 1092–1100. [Google Scholar] [CrossRef]
- Herrero-Martin, D.; Fourtouna, A.; Niedan, S.; Riedmann, L.T.; Schwentner, R.; Aryee, D.N. Factors affecting EWS-FLI1 activity in Ewing’s sarcoma. Sarcoma 2011, 2011, 352580. [Google Scholar] [CrossRef]
- Garcia-Echeverria, C.; Hoffmann, F.; Trumpp, A.; Stamenkovic, I. Development of Ewing’s Sarcoma from Primary Bone Marrow–Derived Mesenchymal Progenitor Cells. Cancer Res. 2005, 65, 11459–11468. [Google Scholar]
- Boro, A.; Prêtre, K.; Rechfeld, F.; Thalhammer, V.; Oesch, S.; Wachtel, M.; Schäfer, B.W.; Niggli, F.K. Small-molecule screen identifies modulators of EWS/FLI1 target gene expression and cell survival in Ewing’s sarcoma. Int. J. Cancer 2012, 131, 2153–2164. [Google Scholar] [CrossRef]
- Palmini, G.; Brandi, M.L. microRNAs and bone tumours: Role of tiny molecules in the development and progression of chondrosarcoma, of giant cell tumour of bone and of Ewing’s sarcoma. Bone 2021, 149, 115968. [Google Scholar] [CrossRef]
- Nishimori, H.; Sasaki, Y.; Yoshida, K.; Irifune, H.; Zembutsu, H.; Tanaka, T.; Aoyama, T.; Hosaka, T.; Kawaguchi, S.; Wada, T. The Id2 gene is a novel target of transcriptional activation by EWS-ETS fusion proteins in Ewing family tumors. Oncogene 2002, 21, 8302–8309. [Google Scholar] [CrossRef]
- Cotterill, S.; Ahrens, S.; Paulussen, M.; Jurgens, H.; Voute, P.; Gadner, H.; Craft, A. Prognostic factors in Ewing’s tumor of bone: Analysis of 975 patients from the European Intergroup Cooperative Ewing’s Sarcoma Study Group. J. Clin. Oncol. 2000, 18, 3108–3114. [Google Scholar] [CrossRef] [PubMed]
- McKinsey, E.; Parrish, J.; Irwin, A.; Niemeyer, B.; Kern, H.; Birks, D.; Jedlicka, P. A novel oncogenic mechanism in Ewing sarcoma involving IGF pathway targeting by EWS/Fli1-regulated microRNAs. Oncogene 2011, 30, 4910–4920. [Google Scholar] [CrossRef] [PubMed]
- Dylla, L.; Moore, C.; Jedlicka, P. MicroRNAs in Ewing sarcoma. Front. Oncol. 2013, 3, 46128. [Google Scholar] [CrossRef] [PubMed]
- Parafioriti, A.; Bason, C.; Armiraglio, E.; Calciano, L.; Daolio, P.A.; Berardocco, M.; Di Bernardo, A.; Colosimo, A.; Luksch, R.; Berardi, A.C. Ewing’s sarcoma: An analysis of miRNA expression profiles and target genes in paraffin-embedded primary tumor tissue. Int. J. Mol. Sci. 2016, 17, 656. [Google Scholar] [CrossRef]
- Li, Z.; Yu, X.; Shen, J.; Wu, W.K.K.; Chan, M.T. MicroRNA expression and its clinical implications in Ewing’s sarcoma. Cell Prolif. 2015, 48, 1–6. [Google Scholar] [CrossRef]
- Sassen, S.; Miska, E.A.; Caldas, C. MicroRNA—Implications for cancer. Virchows Arch. 2008, 452, 1–10. [Google Scholar] [CrossRef]
- Li, M.A.; He, L. microRNAs as novel regulators of stem cell pluripotency and somatic cell reprogramming. Bioessays 2012, 34, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Crist, C.G.; Montarras, D.; Pallafacchina, G.; Rocancourt, D.; Cumano, A.; Conway, S.J.; Buckingham, M. Muscle stem cell behavior is modified by microRNA-27 regulation of Pax3 expression. Proc. Natl. Acad. Sci. USA 2009, 106, 13383–13387. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and-503 are induced during muscle differentiation and promote cell cycle quiescence and differentiation by down-regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138–2149. [Google Scholar] [CrossRef]
- Khach Lai, V.; Ashraf, M.; Jiang, S.; Haider, K. MicroRNA-143 is a critical regulator of cell cycle activity in stem cells with co-overexpression of Akt and angiopoietin-1 via transcriptional regulation of Erk5/cyclin D1 signaling. Cell Cycle 2012, 11, 767–777. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Hassan, M.; Iqbal, M.S.; Naqvi, S.; Alashwal, H.; Moustafa, A.A.; Kloczkowski, A. Prediction of Site Directed miRNAs as Key Players of Transcriptional Regulators Against Influenza C Virus Infection Through Computational Approaches. Front. Mol. Biosci. 2022, 9, 866072. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2017, 26, 865–874. [Google Scholar] [CrossRef]
- Packer, A.N.; Xing, Y.; Harper, S.Q.; Jones, L.; Davidson, B.L. The bifunctional microRNA miR-9/miR-9* regulates REST and CoREST and is downregulated in Huntington’s disease. J. Neurosci. 2008, 28, 14341–14346. [Google Scholar] [CrossRef] [PubMed]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer’s disease correlates with increased BACE1/β-secretase expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6415–6420. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Esser, K.A. MicroRNA-1 and microRNA-133a expression are decreased during skeletal muscle hypertrophy. J. Appl. Physiol. 2007, 102, 306–313. [Google Scholar] [CrossRef]
- Maroney, P.A.; Yu, Y.; Fisher, J.; Nilsen, T.W. Evidence that microRNAs are associated with translating messenger RNAs in human cells. Nat. Struct. Mol. Biol. 2006, 13, 1102–1107. [Google Scholar] [CrossRef]
- Jackson, R.J.; Standart, N. How do microRNAs regulate gene expression? Sci. STKE 2007, 2007, re1. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, T.; Doss C, G.P. Non-coding RNAs in human health and disease: Potential function as biomarkers and therapeutic targets. Funct. Integr. Genom. 2023, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, N.; Al Samarrai, O.R.; Al-Zaidi, H.M.H.; Mirzaei, A.R.; Heidari, M.J. Comprehensive analysis of microRNA (miRNA) in cancer cells. Cell. Mol. Biomed. Rep. 2023, 3, 89–97. [Google Scholar] [CrossRef]
- Baran, Y. MicroRNAs and Long Non-Coding RNAs as Novel Targets in Anti-Cancer Drug Development. Curr. Pharm. Biotechnol. 2023, 24, 913–925. [Google Scholar]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Dalmay, T.; Edwards, D. MicroRNAs and the hallmarks of cancer. Oncogene 2006, 25, 6170–6175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Hou, W. Regulation of angiogenesis by microRNAs in cancer. Mol. Med. Rep. 2021, 24, 583. [Google Scholar] [CrossRef]
- Huang, Q.; Gumireddy, K.; Schrier, M.; Le Sage, C.; Nagel, R.; Nair, S.; Egan, D.A.; Li, A.; Huang, G.; Klein-Szanto, A.J. The microRNAs miR-373 and miR-520c promote tumour invasion and metastasis. Nat. Cell Biol. 2008, 10, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Soheilyfar, S.; Velashjerdi, Z.; Hajizadeh, Y.S.; Maroufi, N.F.; Amini, Z.; Khorrami, A.; Azimian, S.H.; Isazadeh, A.; Taefehshokr, S.; Taefehshokr, N. In vivo and in vitro impact of miR-31 and miR-143 on the suppression of metastasis and invasion in breast cancer. J. BUON 2018, 23, 1290–1296. [Google Scholar] [PubMed]
- Chou, J.; Lin, J.H.; Brenot, A.; Kim, J.-w.; Provot, S.; Werb, Z. GATA3 suppresses metastasis and modulates the tumour microenvironment by regulating microRNA-29b expression. Nat. Cell Biol. 2013, 15, 201–213. [Google Scholar] [CrossRef]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef]
- Zoni, E.; Van Der Horst, G.; Van de Merbel, A.; Chen, L.; Rane, J.; Pelger, R.; Collins, A.; Visakorpi, T.; Snaar-Jagalska, B.; Maitland, N. miR-25 modulates invasiveness and dissemination of human prostate cancer cells via regulation of αv-and α6-integrin expression. Cancer Res. 2015, 75, 2326–2336. [Google Scholar] [CrossRef]
- Hwang, W.-L.; Jiang, J.-K.; Yang, S.-H.; Huang, T.-S.; Lan, H.-Y.; Teng, H.-W.; Yang, C.-Y.; Tsai, Y.-P.; Lin, C.-H.; Wang, H.-W. MicroRNA-146a directs the symmetric division of Snail-dominant colorectal cancer stem cells. Nat. Cell Biol. 2014, 16, 268–280. [Google Scholar] [CrossRef]
- Li, H.-Y.; Liang, J.-L.; Kuo, Y.-L.; Lee, H.-H.; Calkins, M.J.; Chang, H.-T.; Lin, F.-C.; Chen, Y.-C.; Hsu, T.-I.; Hsiao, M. miR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. 2017, 19, 133. [Google Scholar] [CrossRef]
- Franzetti, G.; Laud-Duval, K.; Bellanger, D.; Stern, M.; Sastre-Garau, X.; Delattre, O. MiR-30a-5p connects EWS-FLI1 and CD99, two major therapeutic targets in Ewing tumor. Oncogene 2013, 32, 3915–3921. [Google Scholar] [CrossRef]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. MicroRNA-20b promotes cell proliferation via targeting of TGF-β receptor II and upregulates MYC expression in Ewing’s sarcoma cells. Int. J. Oncol. 2017, 51, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, L.; Shuck, R.; Kurenbekova, L.; Allen-Rhoades, W.; Edwards, D.; Huang, S.; Rajapakshe, K.; Coarfa, C.; Donehower, L.A.; Yustein, J.T. miR-130b directly targets Arhgap1 to drive activation of a metastatic CDC42-PAK1-AP1 positive feedback loop in Ewing sarcoma. Int. J. Cancer 2017, 141, 2062–2075. [Google Scholar] [CrossRef]
- Lu, Q.; Lu, M.; Li, D.; Zhang, S. MicroRNA-34b promotes proliferation, migration and invasion of Ewing’s sarcoma cells by downregulating Notch1. Mol. Med. Rep. 2018, 18, 3577–3588. [Google Scholar] [CrossRef]
- Lobry, C.; Oh, P.; Aifantis, I. Oncogenic and tumor suppressor functions of Notch in cancer: It’s NOTCH what you think. J. Exp. Med. 2011, 208, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yang, F.; Xu, H.; Yue, Z.; Fang, X.; Liu, J. MicroRNA-708 is downregulated in hepatocellular carcinoma and suppresses tumor invasion and migration. Biomed. Pharmacother. 2015, 73, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.-L.; Zhao, H.; Yao, H.-L.; Chen, Y.; Lei, Z.-D.; Liu, K.-J.; Yang, Q. Regulatory roles of microRNA-708 and microRNA-31 in proliferation, apoptosis and invasion of colorectal cancer cells. Oncol. Lett. 2014, 8, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Roberto, G.M.; Vieira, G.M.; Delsin, L.E.A.; de Oliveira Silva, M.; Hakime, R.G.; Engel, E.E.; Scrideli, C.A.; Tone, L.G.; Brassesco, M.S. MiR-708-5p is inversely associated with EWS/FLI1 Ewing sarcoma but does not represent a prognostic predictor. Cancer Genet. 2019, 230, 21–27. [Google Scholar] [CrossRef]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. MicroRNA-181c prevents apoptosis by targeting of FAS receptor in Ewing’s sarcoma cells. Cancer Cell Int. 2018, 18, 37. [Google Scholar] [CrossRef]
- Gao, X.-N.; Lin, J.; Gao, L.; Li, Y.-H.; Wang, L.-L.; Yu, L. MicroRNA-193b regulates c-Kit proto-oncogene and represses cell proliferation in acute myeloid leukemia. Leuk. Res. 2011, 35, 1226–1232. [Google Scholar] [CrossRef]
- Li, Y.; Shao, G.; Zhang, M.; Zhu, F.; Zhao, B.; He, C.; Zhang, Z. miR-124 represses the mesenchymal features and suppresses metastasis in Ewing sarcoma. Oncotarget 2017, 8, 10274. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Parrish, J.K.; Jedlicka, P. MiR-193b, downregulated in Ewing Sarcoma, targets the ErbB4 oncogene to inhibit anchorage-independent growth. PLoS ONE 2017, 12, e0178028. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Naranjo, A.; El-Naggar, A.; Wai, D.H.; Mistry, P.; Lazic, N.; Ayala, F.R.R.; da Cunha, I.W.; Rodriguez-Viciana, P.; Cheng, H.; Tavares Guerreiro Fregnani, J.H. ERBB 4 confers metastatic capacity in ewing sarcoma. EMBO Mol. Med. 2013, 5, 1087–1102. [Google Scholar] [CrossRef] [PubMed]
- Esposito, F.; Tornincasa, M.; Pallante, P.; Federico, A.; Borbone, E.; Pierantoni, G.M.; Fusco, A. Down-regulation of the miR-25 and miR-30d contributes to the development of anaplastic thyroid carcinoma targeting the polycomb protein EZH2. J. Clin. Endocrinol. 2012, 97, E710–E718. [Google Scholar] [CrossRef]
- Chen, D.; Guo, W.; Qiu, Z.; Wang, Q.; Li, Y.; Liang, L.; Liu, L.; Huang, S.; Zhao, Y.; He, X. MicroRNA-30d-5p inhibits tumour cell proliferation and motility by directly targeting CCNE2 in non-small cell lung cancer. Cancer Lett. 2015, 362, 208–217. [Google Scholar] [CrossRef]
- Zenali, M.J.; Zhang, P.L.; Bendel, A.E.; Brown, R.E. Morphoproteomic confirmation of constitutively activated mTOR, ERK, and NF-kappaB pathways in Ewing family of tumors. Ann. Clin. Lab. Sci. 2009, 39, 160–166. [Google Scholar]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, X.; Xiao, Q.; Wang, T.; Shao, G.; Li, Y.; Zhang, Z. MiR-107 suppresses cell proliferation and tube formation of Ewing sarcoma cells partly by targeting HIF-1β. Hum. Cell 2018, 31, 42–49. [Google Scholar] [CrossRef]
- Liu, C.; Li, G.; Ren, S.; Su, Z.; Wang, Y.; Tian, Y.; Liu, Y.; Qiu, Y. miR-185-3p regulates the invasion and metastasis of nasopharyngeal carcinoma by targeting WNT2B in vitro. Oncol. Lett. 2017, 13, 2631–2636. [Google Scholar] [CrossRef]
- Li, S.; Ma, Y.; Hou, X.; Liu, Y.; Li, K.; Xu, S.; Wang, J. MiR-185 acts as a tumor suppressor by targeting AKT1 in non-small cell lung cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 11854. [Google Scholar]
- Ordóñez, J.L.; Osuna, D.; Herrero, D.; de Alava, E.; Madoz-Gúrpide, J. Advances in Ewing’s sarcoma research: Where are we now and what lies ahead? Cancer Res. 2009, 69, 7140–7150. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, J.; Xiao, Q.; Wang, T.; Yu, Y.; Li, B.; Shao, G.; Li, Y.; Zhang, Z. MicroRNA-638 inhibits cell growth and tubule formation by suppressing VEGFA expression in human Ewing sarcoma cells. Biosci. Rep. 2018, 38, BSR20171017. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, G.; Liu, H.; Li, Z.; Yang, Q.; Gu, X.; Du, Z.; Zhang, G.; Wang, J. Integrated bioinformatics analysis of miRNA expression in Ewing sarcoma and potential regulatory effects of miR-21 via targeting ALCAM/CD166. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2114–2122. [Google Scholar] [CrossRef]
- Rodrigues, D.S.; Monteiro, V.S.; Navegantes-Lima, K.C.; Oliveira, A.; Gaspar, S.; Quadros, L.G.; Monteiro, M.C. MicroRNAs in cell cycle progression and proliferation: Molecular mechanisms and pathways. Non-Coding RNA Investig. 2018, 2, 28. [Google Scholar] [CrossRef]
- Sun, D.; Wu, Y.; Zhang, S.; Han, Y.; Shen, J.; Zheng, W.; Wei, L.; Liu, Y.; Ren, L.; Gu, Z. Distinct roles of miR-34 family members on suppression of lung squamous cell carcinoma. Biomed. Pharmacother. 2021, 142, 111967. [Google Scholar] [CrossRef]
- Barrett, C.; Budhiraja, A.; Parashar, V.; Batish, M. The landscape of regulatory noncoding rnas in ewing’s sarcoma. Biomedicines 2021, 9, 933. [Google Scholar] [CrossRef]
- Ye, C.; Yu, X.; Liu, X.; Dai, M.; Zhang, B. miR-30d inhibits cell biological progression of Ewing’s sarcoma by suppressing the MEK/ERK and PI3K/Akt pathways in vitro Corrigendum in/10.3892/ol. 2020.12394. Oncol. Lett. 2018, 15, 4390–4396. [Google Scholar]
- Zhang, S.; Li, D.; Jiao, G.-J.; Wang, H.-L.; Yan, T.-B. miR-185 suppresses progression of Ewing’s sarcoma via inhibiting the PI3K/AKT and Wnt/β-catenin pathways. OncoTargets Ther. 2018, 11, 7967. [Google Scholar] [CrossRef]
- Huang, W.-K.; Yeh, C.-N. The emerging role of MicroRNAs in regulating the drug response of cholangiocarcinoma. Biomolecules 2020, 10, 1396. [Google Scholar] [CrossRef]
- De Feo, A.; Pazzaglia, L.; Ciuffarin, L.; Mangiagli, F.; Pasello, M.; Simonetti, E.; Pellegrini, E.; Ferrari, C.; Bianchi, G.; Spazzoli, B. miR-214-3p Is Commonly Downregulated by EWS-FLI1 and by CD99 and Its Restoration Limits Ewing Sarcoma Aggressiveness. Cancers 2022, 14, 1762. [Google Scholar] [CrossRef]
- Nugent, M. microRNA and bone cancer. Microrna Cancer 2015, 889, 201–230. [Google Scholar]
- Nugent, M. MicroRNA function and dysregulation in bone tumors: The evidence to date. Cancer Manag. Res. 2014, 6, 15. [Google Scholar] [CrossRef]
- Boyerinas, B.; Park, S.-M.; Hau, A.; Murmann, A.E.; Peter, M.E. The role of let-7 in cell differentiation and cancer. Endocr. -Relat. Cancer 2010, 17, F19–F36. [Google Scholar] [CrossRef]
- De Vito, C.; Riggi, N.; Suva, M.-L.; Janiszewska, M.; Horlbeck, J.; Baumer, K.; Provero, P.; Stamenkovic, I. Let-7a is a direct EWS-FLI-1 target implicated in Ewing’s sarcoma development. PLoS ONE 2011, 6, e23592. [Google Scholar] [CrossRef]
- Dong, Q.; Meng, P.; Wang, T.; Qin, W.; Qin, W.; Wang, F.; Yuan, J.; Chen, Z.; Yang, A.; Wang, H. MicroRNA let-7a inhibits proliferation of human prostate cancer cells in vitro and in vivo by targeting E2F2 and CCND2. PLoS ONE 2010, 5, e10147. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Huang, L.; Xiao, Q.; Chen, X.; Zhong, J.; Chen, Y.; Yang, D.; Han, Z.; Shu, Y. Let-7a suppresses macrophage infiltrations and malignant phenotype of Ewing sarcoma via STAT3/NF-κB positive regulatory circuit. Cancer Lett. 2016, 374, 192–201. [Google Scholar] [CrossRef]
- Sand, L.G.; Berghuis, D.; Szuhai, K.; Hogendoorn, P.C. Expression of CCL21 in Ewing sarcoma shows an inverse correlation with metastases and is a candidate target for immunotherapy. Cancer Immunol. Immunother. 2016, 65, 995–1002. [Google Scholar] [CrossRef]
- Bill, C.A.; Allen, C.M.; Vines, C.M. CC chemokine receptor 7 in cancer. Cells 2022, 11, 656. [Google Scholar] [CrossRef]
- Soini, Y. Epigenetic and genetic changes in soft tissue sarcomas: A review. Apmis 2016, 124, 925–934. [Google Scholar] [CrossRef]
- Fu, T.-Y.; Chang, C.-C.; Lin, C.-T.; Lai, C.-H.; Peng, S.-Y.; Ko, Y.-J.; Tang, P.-C. Let-7b-mediated suppression of basigin expression and metastasis in mouse melanoma cells. Exp. Cell Res. 2011, 317, 445–451. [Google Scholar] [CrossRef]
- Hameiri-Grossman, M.; Porat-Klein, A.; Yaniv, I.; Ash, S.; Cohen, I.J.; Kodman, Y.; Haklai, R.; Elad-Sfadia, G.; Kloog, Y.; Chepurko, E. The association between let-7, RAS and HIF-1α in Ewing Sarcoma tumor growth. Oncotarget 2015, 6, 33834. [Google Scholar] [CrossRef]
- Sohn, E.J.; Park, J.; Wu, Y.-P. Accumulation of pre-let-7g and downregulation of mature let-7g with the depletion of EWS. Biochem. Biophys. Res. Commun. 2012, 426, 89–93. [Google Scholar] [CrossRef] [PubMed]
- D’souza, N.; Burns, J.S.; Grisendi, G.; Candini, O.; Veronesi, E.; Piccinno, S.; Horwitz, E.M.; Paolucci, P.; Conte, P.; Dominici, M. MSC and tumors: Homing, differentiation, and secretion influence therapeutic potential. Mesenchymal Stem Cells-Basics Clin. Appl. II 2013, 130, 209–266. [Google Scholar]
- Lin, R.-J.; Lin, Y.-C.; Chen, J.; Kuo, H.-H.; Chen, Y.-Y.; Diccianni, M.B.; London, W.B.; Chang, C.-H.; Yu, A.L. microRNA Signature and Expression of Dicer and Drosha Can Predict Prognosis and Delineate Risk Groups in NeuroblastomamicroRNA, Dicer, and Drosha Expression in Neuroblastoma. Cancer Res. 2010, 70, 7841–7850. [Google Scholar] [CrossRef] [PubMed]
- Merritt, W.M.; Lin, Y.G.; Han, L.Y.; Kamat, A.A.; Spannuth, W.A.; Schmandt, R.; Urbauer, D.; Pennacchio, L.A.; Cheng, J.-F.; Nick, A.M. Dicer, Drosha, and outcomes in patients with ovarian cancer. N. Engl. J. Med. 2008, 359, 2641–2650. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.; Jug, G.; Mestdagh, P.; Schwentner, R.; Kauer, M.; Aryee, D.N.; Schaefer, K.-L.; Nakatani, F.; Scotlandi, K.; Reiter, M. Hsa-mir-145 is the top EWS-FLI1-repressed microRNA involved in a positive feedback loop in Ewing’s sarcoma. Oncogene 2011, 30, 2173–2180. [Google Scholar] [CrossRef]
- Nakatani, F.; Ferracin, M.; Manara, M.C.; Ventura, S.; Del Monaco, V.; Ferrari, S.; Alberghini, M.; Grilli, A.; Knuutila, S.; Schaefer, K.L. miR-34a predicts survival of Ewing’s sarcoma patients and directly influences cell chemo-sensitivity and malignancy. J. Pathol. 2012, 226, 796–805. [Google Scholar] [CrossRef]
- Mosakhani, N.; Guled, M.; Leen, G.; Calabuig-Fariñas, S.; Niini, T.; Machado, I.; Savola, S.; Scotlandi, K.; López-Guerrero, J.A.; Llombart-Bosch, A. An integrated analysis of miRNA and gene copy numbers in xenografts of Ewing’s sarcoma. J. Exp. Clin. Cancer Res. 2012, 31, 24. [Google Scholar] [CrossRef]
- Karnuth, B.; Dedy, N.; Spieker, T.; Lawlor, E.R.; Gattenlöhner, S.; Ranft, A.; Dirksen, U.; Jürgens, H.; Bräuninger, A. Differentially expressed miRNAs in Ewing sarcoma compared to mesenchymal stem cells: Low miR-31 expression with effects on proliferation and invasion. PLoS ONE 2014, 9, e93067. [Google Scholar] [CrossRef]
- Robin, T.P.; Smith, A.; McKinsey, E.; Reaves, L.; Jedlicka, P.; Ford, H.L. EWS/FLI1 Regulates EYA3 in Ewing Sarcoma via Modulation of miRNA-708, Resulting in Increased Cell Survival and ChemoresistanceNovel Mechanisms of Ewing Sarcoma Chemoresistance. Mol. Cancer Res. 2012, 10, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, A.; Manara, M.C.; Sciandra, M.; Zambelli, D.; Nardi, F.; Nicoletti, G.; Garofalo, C.; Meschini, S.; Astolfi, A.; Colombo, M.P. CD99 inhibits neural differentiation of human Ewing sarcoma cells and thereby contributes to oncogenesis. J. Clin. Investig. 2010, 120, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Lamhamedi-Cherradi, S.-E.; Menegaz, B.A.; Ramamoorthy, V.; Vishwamitra, D.; Wang, Y.; Maywald, R.L.; Buford, A.S.; Fokt, I.; Skora, S.; Wang, J. IGF-1R and mTOR blockade: Novel resistance mechanisms and synergistic drug combinations for Ewing sarcoma. JNCI J. Natl. Cancer Inst. 2016, 108, djw182. [Google Scholar] [CrossRef]
- Aryee, D.N.; Fock, V.; Kapoor, U.; Radic-Sarikas, B.; Kovar, H. Zooming in on long non-coding RNAs in ewing sarcoma pathogenesis. Cells 2022, 11, 1267. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Kalariya, N.; Brassil, K.; Calin, G. MicroRNAs: Clinical trials and potential applications. Clin. J. Oncol. Nurs. 2017, 21, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Sun, X.; Zhang, Y.; Liu, L.; Yuan, Q. MiR-598: A tumor suppressor with biomarker significance in osteosarcoma. Life Sci. 2017, 188, 141–148. [Google Scholar] [CrossRef]
- Zhao, Y.; Tu, M.-J.; Yu, Y.-F.; Wang, W.-P.; Chen, Q.-X.; Qiu, J.-X.; Yu, A.-X.; Yu, A.-M. Combination therapy with bioengineered miR-34a prodrug and doxorubicin synergistically suppresses osteosarcoma growth. Biochem. Pharmacol. 2015, 98, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Edwards, W.; Dore, S.; van Schalkwyk, J.; Armson, B.A. Prioritizing maternal sepsis: National adoption of an obstetric early warning system to prevent morbidity and mortality. J. Obstet. Gynaecol. Can. 2020, 42, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Q.; Liu, H.; Hu, B.; Zhou, W.; Cheng, Y. MicroRNA expression and its implication for the diagnosis and therapeutic strategies of gastric cancer. Cancer Lett. 2010, 297, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Farazi, T.A.; Spitzer, J.I.; Morozov, P.; Tuschl, T. miRNAs in human cancer. J. Pathol. 2011, 223, 102–115. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).