
Molecular Mechanisms Involved in MAFLD in Cholecystectomized Patients: A Cohort Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Patients
2.2. RT-qPCR Method
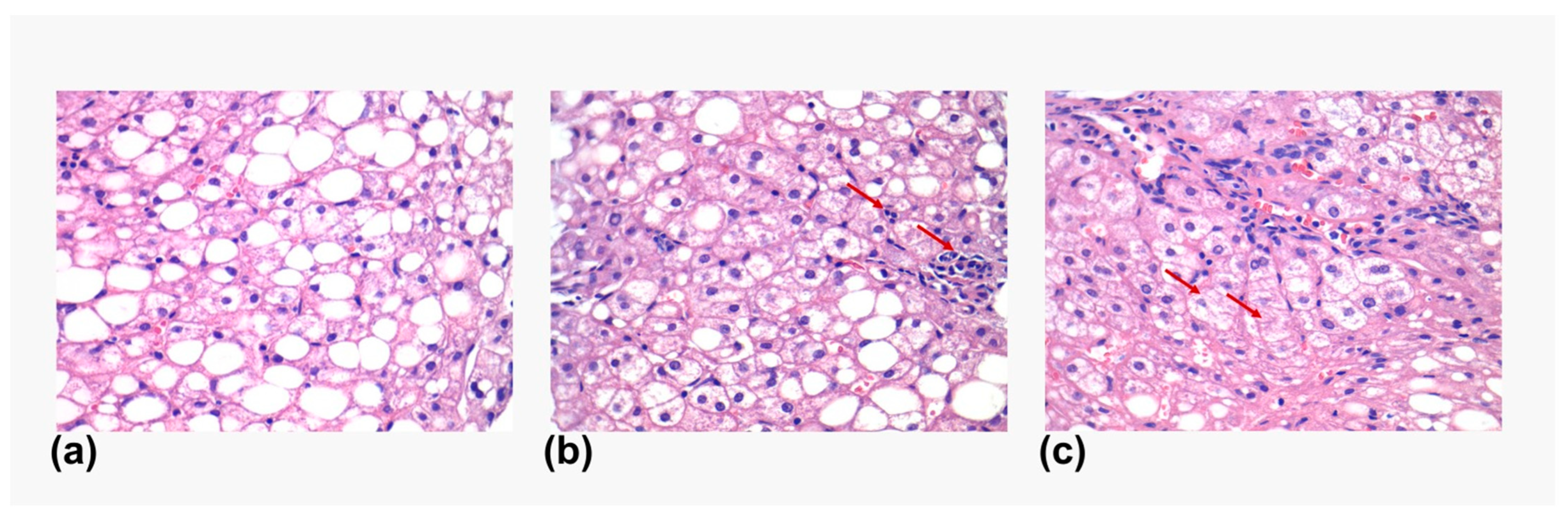
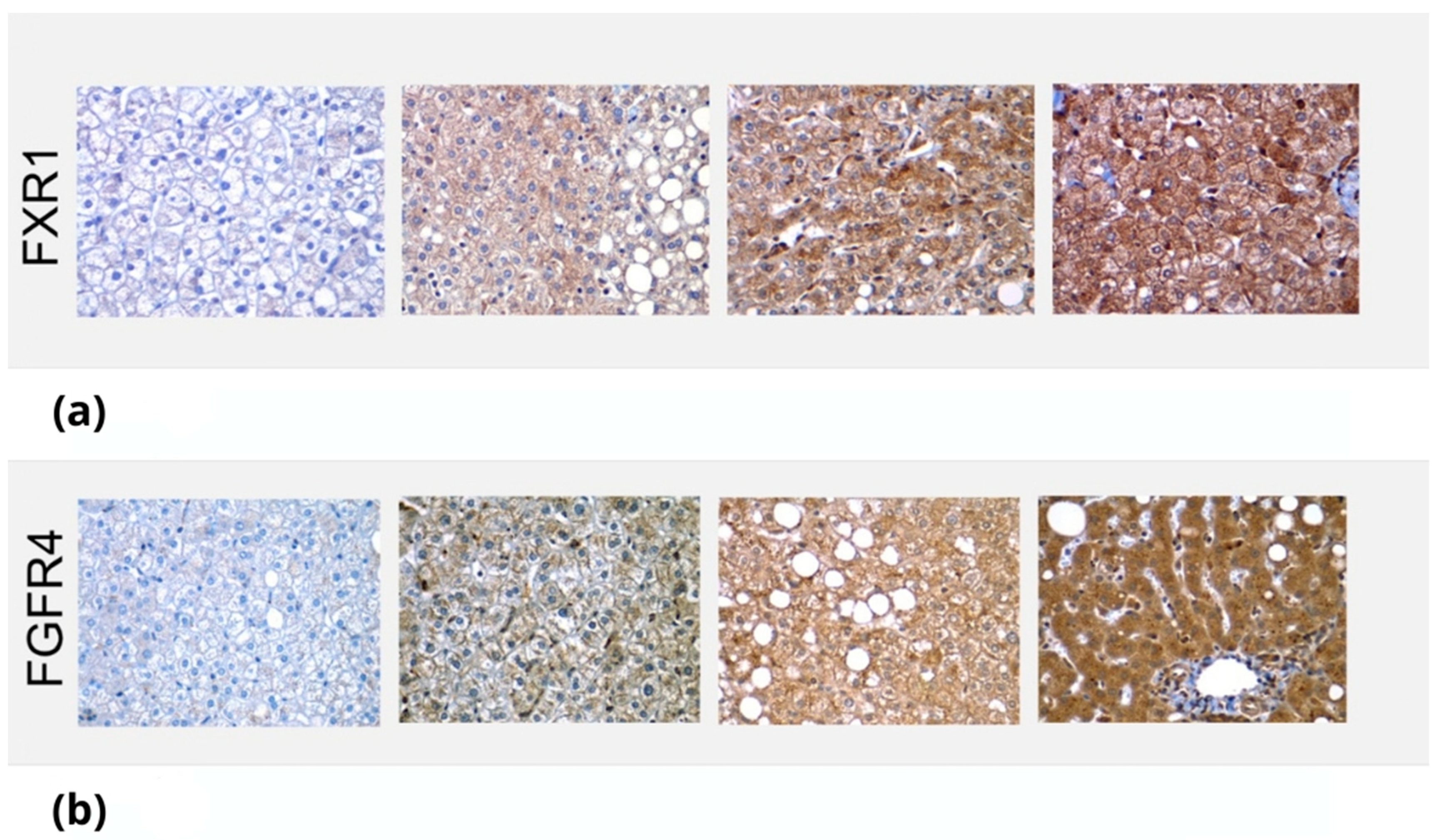
2.3. Immunohistochemistry Methodology
2.4. Statistical Analysis
3. Results
3.1. Baseline Characteristics
3.2. Immunologic Analysis of FXR1 and FGFR4 in Liver Biopsies
3.3. RT-qPCR Results for Liver Biopsies
3.4. 6-Month Follow-Up
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yue, W.; Sun, X.; Du, T. Cholecystectomy versus central obesity or insulin resistance in relation to the risk of nonalcoholic fatty liver disease: The third US National Health and Nutrition Examination Survey. BMC Endocr. Disord. 2019, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Sánchez, N.; Cerda-Reyes, E.; Higuera-de-la-Tijera, F.; Salas-García, A.K.; Cabrera-Palma, S.; Cabrera-Álvarez, G.; Cortez-Hernández, C.; Pérez-Arredondo, L.A.; Purón-González, E.; Coronado-Alejandro, E.; et al. Dyslipidemia as a risk factor for liver fibrosis progression in a multicentric population with non-alcoholic steatohepatitis. F1000Research 2020, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.C.; Méndez-Sánchez, N. Screening for MAFLD: Who, when and how? Ther. Adv. Endocrinol. Metab. 2023, 14, 20420188221145650. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.C.; Méndez-Sánchez, N. Insulin resistance and adipose tissue interactions as the cornerstone of metabolic (dysfunction)-associated fatty liver disease pathogenesis. World J. Gastroenterol. 2023, 29, 3999–4008. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.C.; Eslam, M.; Mendez-Sanchez, N. Detangling the interrelations between MAFLD, insulin resistance, and key hormones. Hormones 2022, 21, 573–589. [Google Scholar] [CrossRef]
- Dan, W.Y.; Yang, Y.S.; Peng, L.H.; Sun, G.; Wang, Z.K. Gastrointestinal microbiome and cholelithiasis: Current status and perspectives. World J. Gastroenterol. 2023, 29, 1589–1601. [Google Scholar] [CrossRef]
- Mendez-Sanchez, N.; Cruz-Ramon, V.C.; Ramirez-Perez, O.L.; Hwang, J.P.; Barranco-Fragoso, B.; Cordova-Gallardo, J. New Aspects of Lipotoxicity in Nonalcoholic Steatohepatitis. Int. J. Mol. Sci. 2018, 19, 2034. [Google Scholar] [CrossRef]
- Méndez-Sánchez, N.; Valencia-Rodríguez, A.; Coronel-Castillo, C.; Vera-Barajas, A.; Contreras-Carmona, J.; Ponciano-Rodríguez, G.; Zamora-Valdés, D. The cellular pathways of liver fibrosis in non-alcoholic steatohepatitis. Ann. Transl. Med. 2020, 8, 400. [Google Scholar] [CrossRef]
- Sánchez-Valle, V.; Chávez-Tapia, N.C.; Uribe, M.; Méndez-Sánchez, N. Role of oxidative stress and molecular changes in liver fibrosis: A review. Curr. Med. Chem. 2012, 19, 4850–4860. [Google Scholar] [CrossRef]
- Almeda-Valdes, P.; Aguilar Olivos, N.E.; Barranco-Fragoso, B.; Uribe, M.; Méndez-Sánchez, N. The Role of Dendritic Cells in Fibrosis Progression in Nonalcoholic Fatty Liver Disease. BioMed Res. Int. 2015, 2015, 768071. [Google Scholar] [CrossRef]
- Gutt, C.; Schläfer, S.; Lammert, F. The Treatment of Gallstone Disease. Dtsch. Arztebl. Int. 2020, 117, 148–158. [Google Scholar] [CrossRef]
- Luo, D.; Chen, X.P.; Dai, Y.; Kuang, F.; Kang, M.J.; Li, B.; Su, S. Cholecystectomy and risk of liver disease: A systematic review and meta-analysis of 27 million individuals. Int. J. Surg. 2023, 109, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Di Ciaula, A.; Garruti, G.; Wang, D.Q.; Portincasa, P. Cholecystectomy and risk of metabolic syndrome. Eur. J. Intern. Med. 2018, 53, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, S.; Tian, Y. Cholecystectomy as a risk factor of metabolic syndrome: From epidemiologic clues to biochemical mechanisms. Lab. Investig. 2018, 98, 7–14. [Google Scholar] [CrossRef]
- Konyn, P.; Alshuwaykh, O.; Dennis, B.B.; Cholankeril, G.; Ahmed, A.; Kim, D. Gallstone Disease and Its Association With Nonalcoholic Fatty Liver Disease, All-Cause and Cause-Specific Mortality. Clin. Gastroenterol. Hepatol. 2023, 21, 940–948.e942. [Google Scholar] [CrossRef]
- Méndez-Sánchez, N.; Valencia-Rodríguez, A.; Cerda-Reyes, E.; Montejo-Velázquez, C.; Higuera de la Tijera, F.; Servin-Caamaño, A.; Icaza-Chávez, M.E.; Chi-Cervera, L.A.; Montalvo-Gordon, I.; Qi, X.; et al. Long-standing effect of cholecystectomy in patients with metabolic-associated fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2021, 33, e824–e830. [Google Scholar] [CrossRef]
- Cortés, V.; Quezada, N.; Uribe, S.; Arrese, M.; Nervi, F. Effect of cholecystectomy on hepatic fat accumulation and insulin resistance in non-obese Hispanic patients: A pilot study. Lipids Health Dis. 2017, 16, 129. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Antonio, I.; López-Sánchez, G.N.; Reyes-Gómez, V.A.; Contreras-Flores, E.H.; Farías-García, F.; Espejel-Deloiza, M.; Durán-Padilla, M.A.; Chablé-Montero, F.; Uribe, M.; Chávez-Tapia, N.C.; et al. Laparoscopic cholecystectomy: Histopathological analysis of metabolic associated fatty liver disease and fibrosis. Ann. Hepatol. 2022, 27, 100651. [Google Scholar] [CrossRef]
- Arrese, M.; Cortés, V.; Barrera, F.; Nervi, F. Nonalcoholic fatty liver disease, cholesterol gallstones, and cholecystectomy: New insights on a complex relationship. Curr. Opin. Gastroenterol. 2018, 34, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, A.; Canbay, A. Why Bile Acids Are So Important in Non-Alcoholic Fatty Liver Disease (NAFLD) Progression. Cells 2019, 8, 1358. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.Y.; Chen, S.M.; Pan, C.X.; Li, Y. FXR: Structures, biology, and drug development for NASH and fibrosis diseases. Acta Pharmacol. Sin. 2022, 43, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Saha, P.K.; Chan, L.; Moore, D.D. Farnesoid X receptor is essential for normal glucose homeostasis. J. Clin. Investig. 2006, 116, 1102–1109. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, Y.; An, Y.; Zhao, N.; Zhao, Y.; Yu, C. Soluble FGFR4 extracellular domain inhibits FGF19-induced activation of FGFR4 signaling and prevents nonalcoholic fatty liver disease. Biochem. Biophys. Res. Commun. 2011, 409, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, F.; Jin, C.; Wu, X.; Chan, W.K.; McKeehan, W.L. Increased carbon tetrachloride-induced liver injury and fibrosis in FGFR4-deficient mice. Am. J. Pathol. 2002, 161, 2003. [Google Scholar] [CrossRef]
- Ocker, M. Fibroblast growth factor signaling in non-alcoholic fatty liver disease and non-alcoholic steatohepatitis: Paving the way to hepatocellular carcinoma. World J. Gastroenterol. 2020, 26, 279–290. [Google Scholar] [CrossRef]
- Kim, I.; Ahn, S.H.; Inagaki, T.; Choi, M.; Ito, S.; Guo, G.L.; Kliewer, S.A.; Gonzalez, F.J. Differential regulation of bile acid homeostasis by the farnesoid X receptor in liver and intestine. J. Lipid Res. 2007, 48, 2664–2672. [Google Scholar] [CrossRef]
- Schaap, F.G.; Trauner, M.; Jansen, P.L. Bile acid receptors as targets for drug development. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 55–67. [Google Scholar] [CrossRef]
- Al-Khaifi, A.; Rudling, M.; Angelin, B. An FXR Agonist Reduces Bile Acid Synthesis Independently of Increases in FGF19 in Healthy Volunteers. Gastroenterology 2018, 155, 1012–1016. [Google Scholar] [CrossRef]
- Chen, C.; Patel, S.; Corisdeo, S.; Liu, X.; Micolochick, H.; Xue, J.; Yang, Q.; Lei, Y.; Wang, B.; Soltis, D. Generation and characterization of a panel of monoclonal antibodies specific for human fibroblast growth factor receptor 4 (FGFR4). Hybridoma 2005, 24, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.; Wang, L.; Chiang, J.Y.; Zhang, Y.; Klaassen, C.D.; Guo, G.L. Mechanism of tissue-specific farnesoid X receptor in suppressing the expression of genes in bile-acid synthesis in mice. Hepatology 2012, 56, 1034–1043. [Google Scholar] [CrossRef]
- Watanabe, M.; Houten, S.M.; Wang, L.; Moschetta, A.; Mangelsdorf, D.J.; Heyman, R.A.; Moore, D.D.; Auwerx, J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J. Clin. Investig. 2004, 113, 1408–1418. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Parra-Landazury, N.M.; Cordova-Gallardo, J.; Méndez-Sánchez, N. Obesity and Gallstones. Visc. Med. 2021, 37, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Moreau, F.; Brunao, B.B.; Liu, X.-Y.; Tremblay, F.; Fitzgerald, K.; Avila-Pacheco, J.; Clish, C.; Kahn, R.C.; Softic, S. Liver-specific FGFR4 knockdown in mice on an HFD increases bile acid synthesis and improves hepatic steatosis. J. Lipid Res. 2023, 64, 100324. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shi, X.; Zheng, Q.; Wang, X.; Liu, X.; Tan, M.; Lv, G.; Zhang, P.; Martin, R.C.; Li, Y. Aberrant FGFR4 signaling worsens nonalcoholic steatohepatitis in FGF21KO mice. Int. J. Biol. Sci. 2021, 17, 2576–2589. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, X.; Valanejad, L.; Vasilenko, A.; More, V.; Qiu, X.; Chen, W.; Lai, Y.; Slitt, A.; Stoner, M.; et al. Bile salt export pump is dysregulated with altered farnesoid X receptor isoform expression in patients with hepatocellular carcinoma. Hepatology 2013, 57, 1530–1541. [Google Scholar] [CrossRef]
- Méndez-Sánchez, N.; Pal, S.C.; Córdova-Gallardo, J. How far are we from an approved drug for nonalcoholic steatohepatitis? Expert Opin. Pharmacother. 2023, 24, 1021–1038. [Google Scholar] [CrossRef]
- Gillard, J.; Clerbaux, L.A.; Nachit, M.; Sempoux, C.; Staels, B.; Bindels, L.B.; Tailleux, A.; Leclercq, I.A. Bile acids contribute to the development of non-alcoholic steatohepatitis in mice. JHEP Rep. 2022, 4, 100387. [Google Scholar] [CrossRef]
- Xie, Z.Q.; Li, H.X.; Tan, W.L.; Yang, L.; Ma, X.W.; Li, W.X.; Wang, Q.B.; Shang, C.Z.; Chen, Y.J. Association of Cholecystectomy With Liver Fibrosis and Cirrhosis Among Adults in the USA: A Population-Based Propensity Score-Matched Study. Front. Med. 2021, 8, 787777. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Gene—Genbank Entry Primers | Exon Location Primer Sequences | Amplicon Size bp |
|---|---|---|
| FGFR4 NM_002011 | 13–14 | 180 |
| FGFR4-F | tgctggtgactgaggacaat | |
| FGFR4-R | ggatcccaaaagaccacacg | |
| FXR NM_001206979 | 8–9 | 250 |
| FXR-F | cgacaagtgacctcgacaac | |
| FXR-R | ggtccaaagtctgaaatcctgg | |
| HPRT NM_000194 | 1–3 | 176 |
| HPRT-F | cctggcgtcgtgattagtga | |
| HPRT-R | gctacaatgtgatggcctcc | |
| HMBS NM_001024382 | 1–2 | 233 |
| HMBS-F | gatgagagtgattcgcgtgg | |
| HMBS-R | gaacaaccaggtccacttca |
| Characteristic | Total (N = 12) |
|---|---|
| Age (years) | 48 (33–56) |
| BMI (kg/m2) | 29.47 (21.22–40.62) |
| Patients with liver steatosis | |
| Grade 0 | 6 (50%) |
| Grade 1 | 1 (8.33%) |
| Grade 2 | 3 (25%) |
| Grade 3 | 2 (16.66%) |
| Patients with liver fibrosis | |
| Grade 1 | 9 (75%) |
| Grade 2 | 3 (3%) |
| Bilirubin (mg/dL) | 0.81 (0.58–1.52) |
| ALT (U/L) | 36.00 (18.50–51.75) |
| AST (U/L) | 29 (16.25–34.25) |
| GGT (U/L) | 28.50 (14.00–131.00) |
| ALP (U/dL) | 70.00 (50.00–101.75) |
| Albumin (g/L) | 3.92 (3.72–4.21) |
| Cholesterol (mg/dL) | 174.10 (146.25–191.00) |
| Cholesterol HDL (mg/dL) | 39.10 (36.25–48.65) |
| Cholesterol LDL (mg/dL) | 111.50 (87.25–128.00) |
| Triglycerides (mg/dL) | 94.71 (76.88–127.04) |
| Glucose (mg/dL) | 100.80 (85.82–120.15) |
| HbA1c (%) | 5.50 (5.25–5.75) |
| Insulin (μIU/mL) | 7.60 (4.90–11.60) |
| Efficiency | 1.54 | 1.67 | 1.605 | 1.68 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 28.55 | 33.06 | dCt (Target) | E (dCt Target) | dCt (hkg) | E (dCt hkg) | Ratio | ||
| Sample | HPRT | HMBS | MG HK | FXR | |||||
| 1 | 29.85 | 27.24 | 28.55 | 33.06 | 0.00 | 1.00 | 0.00 | 1.00 | 1.00 |
| 2 | 26.84 | 24.75 | 25.80 | 32.54 | 0.52 | 1.31 | 2.76 | 3.68 | 0.36 |
| 3 | 16.67 | 15.90 | 16.29 | 18.18 | 14.88 | 2252.17 | 12.27 | 331.25 | 6.80 |
| 4 | 15.52 | 14.74 | 15.13 | 16.68 | 16.38 | 4904.18 | 13.42 | 572.10 | 8.57 |
| 5 | 15.70 | 14.78 | 15.24 | 16.51 | 16.55 | 5356.35 | 13.31 | 543.09 | 9.86 |
| 6 | 15.41 | 14.65 | 15.03 | 16.14 | 16.92 | 6489.83 | 13.52 | 599.82 | 10.82 |
| 7 | 15.38 | 14.45 | 14.92 | 16.26 | 16.80 | 6098.12 | 13.64 | 633.36 | 9.63 |
| 8 | 18.20 | 17.93 | 18.07 | 20.25 | 12.81 | 769.51 | 10.49 | 142.69 | 5.39 |
| 9 | 19.19 | 18.19 | 18.69 | 19.17 | 13.89 | 1347.55 | 9.86 | 106.17 | 12.69 |
| 10 | 22.52 | 20.69 | 21.61 | 23.36 | 9.80 | 161.45 | 6.95 | 26.73 | 6.04 |
| 11 | 22.11 | 20.76 | 21.44 | 23.04 | 10.02 | 180.97 | 7.12 | 28.97 | 6.25 |
| 12 | 21.86 | 20.56 | 21.21 | 23.17 | 9.89 | 169.16 | 7.34 | 32.22 | 5.25 |
| Efficiency | 1.54 | 1.67 | 1.605 | 1.68 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 28.55 | 27.66 | dCt (Target) | E (dCt Target) | dCt (hkg) | E (dCt hkg) | Ratio | ||
| Sample | HPRT | HMBS | MG HK | FGFR | |||||
| 1 | 29.85 | 27.24 | 28.55 | 27.66 | 0.00 | 1.00 | 0.00 | 1.00 | 1.00 |
| 2 | 26.84 | 24.75 | 25.80 | 27.64 | 0.02 | 1.01 | 2.76 | 3.68 | 0.27 |
| 3 | 16.67 | 15.90 | 16.29 | 20.44 | 7.22 | 38.83 | 12.27 | 331.25 | 0.12 |
| 4 | 15.52 | 14.74 | 15.13 | 18.68 | 8.98 | 94.75 | 13.42 | 572.10 | 0.17 |
| 5 | 15.70 | 14.78 | 15.24 | 17.81 | 9.85 | 147.25 | 13.31 | 543.09 | 0.27 |
| 6 | 15.41 | 14.65 | 15.03 | 19.32 | 8.34 | 68.50 | 13.52 | 599.82 | 0.11 |
| 7 | 15.38 | 14.45 | 14.92 | 18.57 | 9.09 | 100.18 | 13.64 | 633.36 | 0.16 |
| 8 | 18.20 | 17.93 | 18.07 | 21.79 | 5.87 | 19.59 | 10.49 | 142.69 | 0.14 |
| 9 | 19.19 | 18.19 | 18.69 | 22.21 | 5.45 | 15.83 | 9.86 | 106.17 | 0.15 |
| 10 | 22.52 | 20.69 | 21.61 | 22.51 | 5.15 | 13.60 | 6.95 | 26.73 | 0.51 |
| 11 | 22.11 | 20.76 | 21.44 | 22.90 | 4.76 | 11.16 | 7.12 | 28.97 | 0.39 |
| 12 | 21.86 | 20.56 | 21.21 | 23.88 | 3.78 | 6.79 | 7.34 | 32.22 | 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, S.C.; Castillo-Castañeda, S.M.; Díaz-Orozco, L.E.; Ramírez-Mejía, M.M.; Dorantes-Heredia, R.; Alonso-Morales, R.; Eslam, M.; Lammert, F.; Méndez-Sánchez, N. Molecular Mechanisms Involved in MAFLD in Cholecystectomized Patients: A Cohort Study. Genes 2023, 14, 1935. https://doi.org/10.3390/genes14101935
Pal SC, Castillo-Castañeda SM, Díaz-Orozco LE, Ramírez-Mejía MM, Dorantes-Heredia R, Alonso-Morales R, Eslam M, Lammert F, Méndez-Sánchez N. Molecular Mechanisms Involved in MAFLD in Cholecystectomized Patients: A Cohort Study. Genes. 2023; 14(10):1935. https://doi.org/10.3390/genes14101935
Chicago/Turabian StylePal, Shreya C., Stephany M. Castillo-Castañeda, Luis E. Díaz-Orozco, Mariana M. Ramírez-Mejía, Rita Dorantes-Heredia, Rogelio Alonso-Morales, Mohammed Eslam, Frank Lammert, and Nahum Méndez-Sánchez. 2023. "Molecular Mechanisms Involved in MAFLD in Cholecystectomized Patients: A Cohort Study" Genes 14, no. 10: 1935. https://doi.org/10.3390/genes14101935
APA StylePal, S. C., Castillo-Castañeda, S. M., Díaz-Orozco, L. E., Ramírez-Mejía, M. M., Dorantes-Heredia, R., Alonso-Morales, R., Eslam, M., Lammert, F., & Méndez-Sánchez, N. (2023). Molecular Mechanisms Involved in MAFLD in Cholecystectomized Patients: A Cohort Study. Genes, 14(10), 1935. https://doi.org/10.3390/genes14101935