
Differential Expression of Circadian Clock Genes in the Bovine Neuroendocrine Adrenal System
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Procedures
2.2. Sample Preparation and RNA Extraction
2.3. RNA Sequencing and Annotation
2.4. Statistical Analyses
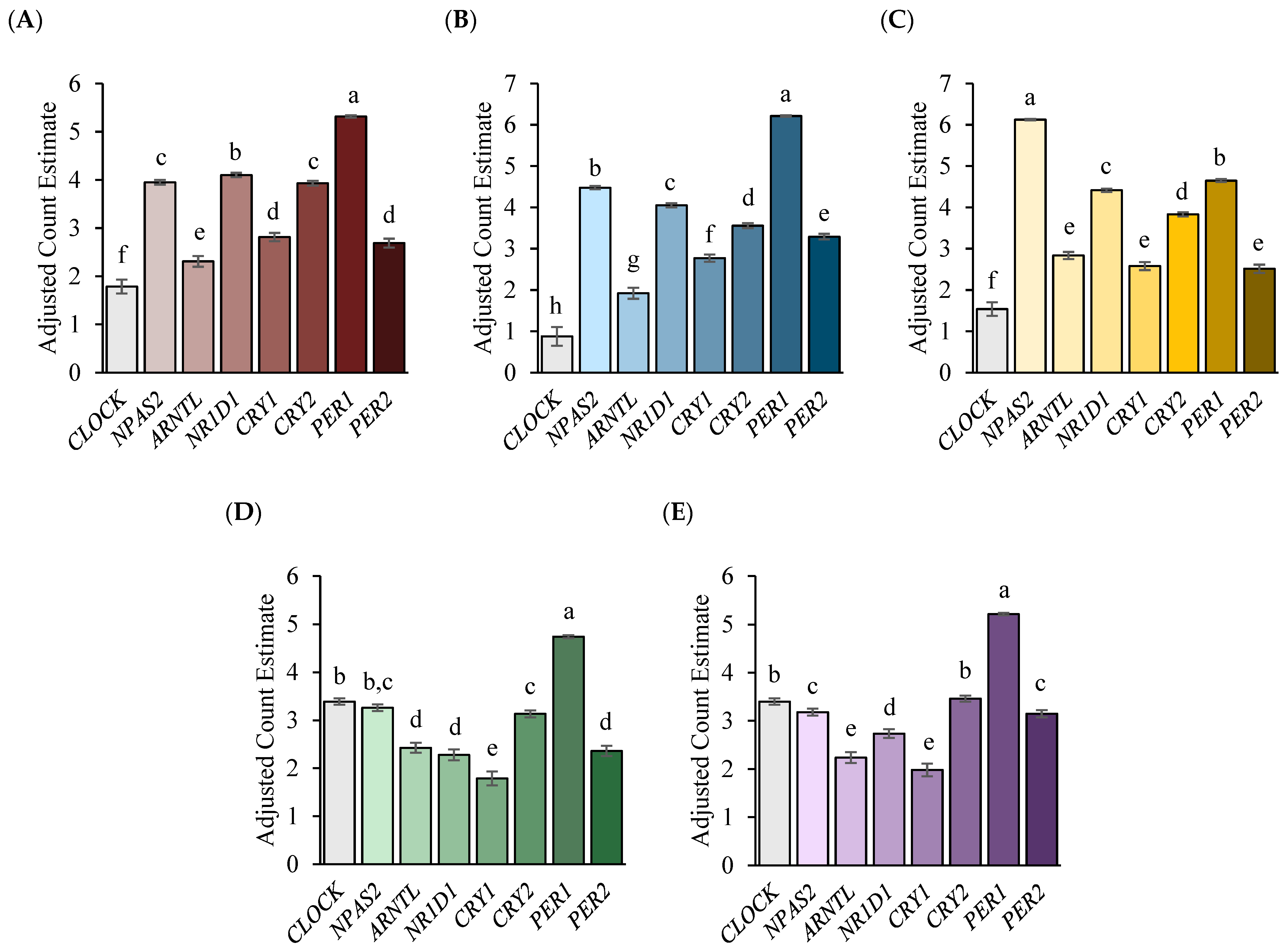
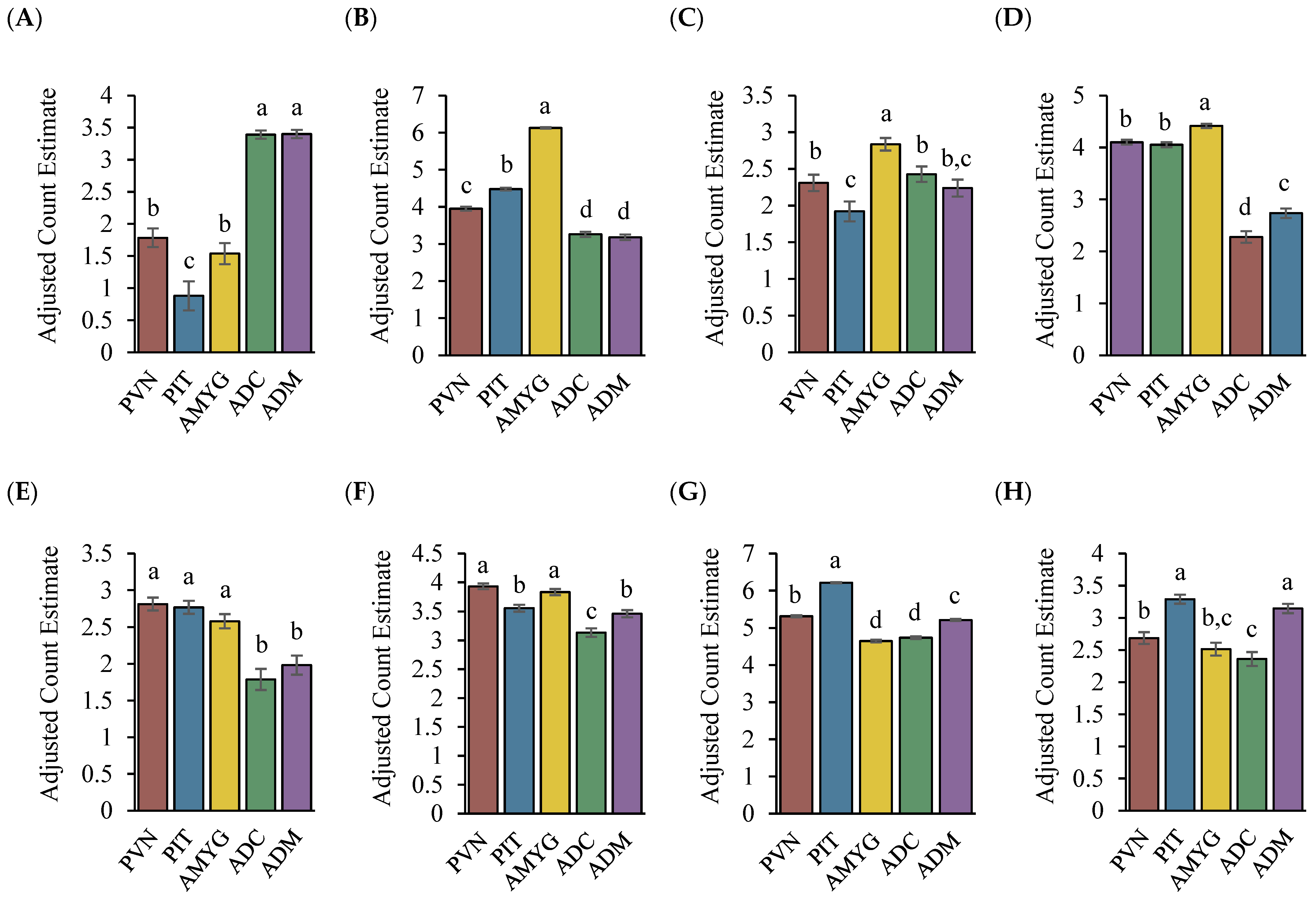
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albrecht, U. Invited review: Regulation of mammalian circadian clock genes. J. Appl. Physiol. 2002, 92, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Son, G.H.; Kim, K. Adrenal peripheral oscillator in generating the circadian glucocorticoid rhythm. Ann. N. Y. Acad. Sci. 2011, 1220, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Dumbell, R.; Matveeva, O.; Oster, H. Circadian clocks, stress, and immunity. Front. Endocrinol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Son, G.H.; Cha, H.K.; Chung, S.; Kim, K. Multimodal regulation of circadian glucocorticoid rhythm by central and adrenal clocks. J. Endocr. Soc. 2018, 2, 444–459. [Google Scholar] [CrossRef]
- Hastings, M. The brain, circadian rhythms, and clock genes. Br. Med. J. 1998, 317, 1704–1707. [Google Scholar] [CrossRef] [PubMed]
- Rosenwasser, A.M.; Turek, F.W. Neurobiology of circadian rhythm regulation. Sleep Med. Clin. 2015, 10, 403–412. [Google Scholar] [CrossRef]
- Balsalobre, A. Clock genes in mammalian peripheral tissues. Cell Tissue Res. 2002, 309, 193–199. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164–179. [Google Scholar] [CrossRef]
- Crumbley, C.; Burris, T.P. Direct regulation of CLOCK expression by REV-ERB. PLoS ONE 2011, 6, e17290. [Google Scholar] [CrossRef]
- DeBruyne, J.P.; Weaver, D.R.; Reppert, S.M. CLOCK and NPAS2 have overlapping roles in the suprachiasmatic circadian clock. Nat. Neurosci. 2007, 10, 543–545. [Google Scholar] [CrossRef]
- Landgraf, D.; Wang, L.L.; Diemer, T.; Welsh, D.K. NPAS2 compensates for loss of CLOCK in peripheral circadian oscillators. PLoS Genet. 2016, 12, e1005882. [Google Scholar] [CrossRef] [PubMed]
- Tonsfeldt, K.J.; Chappell, P.E. Clocks on top: The role of the circadian clock in the hypothalamic and pituitary regulation of endocrine physiology. Mol. Cell. Endocrinol. 2012, 349, 3–12. [Google Scholar] [CrossRef]
- Lin, X.W.; Blum, I.D.; Storch, K.F. Clocks within the master gland: Hypophyseal rhythms and their physiological significance. J. Biol. Rhythm. 2015, 30, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Segall, L.A.; Milet, A.; Tronche, F.; Amir, S. Brain glucocorticoid receptors are necessary for the rhythmic expression of the clock protein, PERIOD2, in the central extended amygdala in mice. Neurosci. Lett. 2009, 457, 58–60. [Google Scholar] [CrossRef]
- Albrecht, A.; Stork, O. Circadian rhythms in fear conditioning: An overview of behavioral, brain system, and molecular interactions. Neural Plast. 2017, 2017, 3750307. [Google Scholar] [CrossRef]
- Shimizu, T.; Hirai, Y.; Murayama, C.; Miyamoto, A.; Miyazaki, H.; Miyazaki, K. Circadian Clock genes Per2 and clock regulate steroid production, cell proliferation, and luteinizing hormone receptor transcription in ovarian granulosa cells. Biochem. Biophys. Res. Commun. 2011, 412, 132–135. [Google Scholar] [CrossRef]
- Ginther, O.J.; Pinaffi, F.L.; Khan, F.A.; Duarte, L.F.; Beg, M.A. Circadian influence on the preovulatory LH surge, ovulation, and prolactin concentrations in heifers. Theriogenology 2013, 79, 528–533. [Google Scholar] [CrossRef]
- Casey, T.M.; Plaut, K. Lactation Biology Symposium: Circadian clocks as mediators of the homeorhetic response to lactation. J. Anim. Sci. 2012, 90, 744–754. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, Z.; Khan, M.J.; Gao, J.; Loor, J.J. Clock circadian regulator (CLOCK) gene network expression patterns in bovine adipose, liver, and mammary gland at 3 time points during the transition from pregnancy into lactation. J. Dairy Sci. 2015, 98, 4601–4612. [Google Scholar] [CrossRef]
- Lefcourt, A.M.; Bitman, J.; Kahl, S.; Wood, D.L. Circadian and ultradian rhythms of peripheral cortisol concentrations in lactating dairy cows. J. Dairy Sci. 1993, 76, 2607–2612. [Google Scholar] [CrossRef]
- Piccione, G.; Grasso, F.; Giudice, E. Circadian rhythm in the cardiovascular system of domestic animals. Res. Vet. Sci. 2005, 79, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Nebzydoski, S.J.; Pozzo, S.; Nemec, L.; Rankin, M.K.; Gressley, T.F. The effect of dexamethasone on clock gene mRNA levels in bovine neutrophils and lymphocytes. Vet. Immunol. Immunopathol. 2010, 138, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Fazio, F.; Caola, G.; Refinetti, R. Daily rhythmicity of glycemia in four species of domestic animals under various feeding regimes. J. Physiol. Sci. 2008, 58, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Ying, Y.; Bartell, P.A.; Harvatine, K.J. The effects of feeding time on milk production, total-tract digestibility, and daily rhythms of feeding behavior and plasma metabolites and hormones in dairy cows. J. Dairy Sci. 2014, 97, 7764–7776. [Google Scholar] [CrossRef]
- Federation of Animal Science Societies. Guide for Care and Use of Agricultural Animals in Research and Teaching, 3rd ed.; Federation of Animal Science Societies: Champaign, IL, USA, 2010. [Google Scholar]
- Rhodes, C.H.; Morriell, J.I.; Pfaff, D.W. Immunohistochemical analysis of magnocellular elements in rat hypothalamus: Distribution and numbers of cells containing neurophysin, oxytocin, and vasopressin. J. Comp. Neurol. 1981, 198, 45–64. [Google Scholar] [CrossRef]
- Qi, Y.; Namavar, M.R.; Iqbal, J.; Oldfield, B.J.; Clarke, I.J. Characterization of the projections to the hypothalamic paraventricular and periventricular nuclei in the female sheep brain, using retrograde tracing and immunohistochemistry. Neuroendocrinology 2009, 90, 31–53. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Valenzuela, F.J.; Torres-Farfan, C.; Richter, H.G.; Mendez, N.; Campino, C.; Torrealba, F.; Valenzuela, G.J.; Seron-Ferre, M. Clock gene expression in adult primate suprachiasmatic nuclei and adrenal: Is the adrenal a peripheral clock responsive to melatonin? Endocrinology 2008, 149, 1454–1461. [Google Scholar] [CrossRef]
- Chun, L.E.; Christensen, J.; Woodruff, E.R.; Morton, S.J.; Hinds, L.R.; Spencer, R.L. Adrenal-dependent and -independent stress-induced Per1 mRNA in hypothalamic paraventricular nucleus and prefrontal cortex of male and female rats. Stress 2018, 21, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Cannella, V.; Monteverde, V.; Bertolucci, C.; Frigato, E.; Congiu, F.; Guercio, A. Circadian gene expression in peripheral blood of Bos taurus under different experimental condition. J. Appl. Biomed. 2014, 12, 271–275. [Google Scholar] [CrossRef]
- Bozek, K.; Relogio, A.; Kielbasa, S.M.; Heine, M.; Dame, C.; Kramer, A.; Herzel, H. Regulation of clock-controlled genes in mammals. PLoS ONE 2009, 4, e4882. [Google Scholar] [CrossRef] [PubMed]
- So, A.Y.; Bernal, T.U.; Pillsbury, M.L.; Yamamoto, K.R.; Feldman, B.J. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 17582–17587. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Nakahata, Y.; Tanaka, M.; Yoshida, M.; Soma, H.; Shinohara, K.; Yasuda, A.; Mamine, T.; Takumi, T. Acute physical stress elevates mouse period1 mRNA expression in mouse peripheral tissues via a glucocorticoid-responsive element. J. Biol. Chem. 2005, 280, 42036–42043. [Google Scholar] [CrossRef] [PubMed]
- Al-Safadi, S.; Branchaud, M.; Rutherford, S.; Amir, S. Glucocorticoids and stress-induced changes in the expression of PERIOD1 in the rat forebrain. PLoS ONE 2015, 10, e0130085. [Google Scholar] [CrossRef]
- Bur, I.M.; Zouaoui, S.; Fontanaud, P.; Coutry, N.; Molino, F.; Martin, A.O.; Mollard, P.; Bonnefont, X. The comparison between circadian oscillators in mouse liver and pituitary gland reveals different integration of feeding and light schedules. PLoS ONE 2010, 5, e15316. [Google Scholar] [CrossRef]
- Sitzmann, B.D.; Lemos, D.R.; Ottinger, M.A.; Urbanski, H.F. Effects of age on clock gene expression in the rhesus macaque pituitary gland. Neurobiol. Aging 2010, 31, 696–705. [Google Scholar] [CrossRef]
- Wunderer, F.; Kuhne, S.; Jilg, A.; Ackermann, K.; Sebesteny, T.; Maronde, E.; Stehle, J.H. Clock gene expression in the human pituitary gland. Endocrinology 2013, 154, 2046–2057. [Google Scholar] [CrossRef]
- Reick, M.; Garcia, J.A.; Dudley, C.; McKnight, S.L. NPAS2: An analog of clock operative in the mammalian forebrain. Science 2001, 293, 506–509. [Google Scholar] [CrossRef]
- Garcia, J.A.; Zhang, D.; Estill, S.J.; Michnoff, C.; Rutter, J.; Reick, M.; Scott, K.; Diaz-Arrastia, R.; McKnight, S.L. Impaired cued and contextual memory in NPAS2-deficient mice. Science 2000, 288, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- Ozburn, A.R.; Kern, J.; Parekh, P.K.; Logan, R.W.; Liu, Z.; Falcon, E.; Becker-Krail, D.; Purohit, K.; Edgar, N.M.; Huang, Y.; et al. NPAS2 Regulation of anxiety-like behavior and GABAA receptors. Front. Mol. Neurosci. 2017, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Ressler, K.J. Amygdala activity, fear, and anxiety: Modulation by stress. Biol. Psychiatry 2010, 67, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Hermans, E.J.; Battaglia, F.P.; Atsak, P.; de Voogd, L.D.; Fernandez, G.; Roozendaal, B. How the amygdala affects emotional memory by altering brain network properties. Neurobiol. Learn. Mem. 2014, 112, 2–16. [Google Scholar] [CrossRef]
- Kondratova, A.A.; Dubrovsky, Y.V.; Antoch, M.P.; Kondratov, R.V. Circadian clock proteins control adaptation to novel environment and memory formation. Aging 2010, 2, 285–297. [Google Scholar] [CrossRef]
- Haque, S.N.; Booreddy, S.R.; Welsh, D.K. Effects of BMAL1 manipulation on the brain’s master circadian clock and behavior. Yale J. Biol. Med. 2019, 92, 251–258. [Google Scholar] [PubMed]
- Astiz, M.; Heyde, I.; Oster, H. Mechanisms of communication in the mammalian circadian timing system. Int. J. Mol. Sci. 2019, 20, 343. [Google Scholar] [CrossRef]
- Leliavski, A.; Dumbell, R.; Ott, V.; Oster, H. Adrenal clocks and the role of adrenal hormones in the regulation of circadian physiology. J. Biol. Rhythm. 2015, 30, 20–34. [Google Scholar] [CrossRef]
- Jager, J.; O’Brien, W.T.; Manlove, J.; Krizman, E.N.; Fang, B.; Gerhart-Hines, Z.; Robinson, M.B.; Klein, P.S.; Lazar, M.A. Behavioral changes and dopaminergic dysregulation in mice lacking the nuclear receptor Rev-erb alpha. Mol. Endocrinol. 2014, 28, 490–498. [Google Scholar] [CrossRef]
- Gerhart-Hines, Z.; Lazar, M.A. Rev-erb alpha and the circadian transcriptional regulation of metabolism. Diabetes Obes. Metab. 2015, 17 (Suppl. S1), 12–16. [Google Scholar] [CrossRef]
- Woldt, E.; Sebti, Y.; Solt, L.A.; Duhem, C.; Lancel, S.; Eeckhoute, J.; Hesselink, M.K.; Paquet, C.; Delhaye, S.; Shin, Y.; et al. Rev-erb-alpha modulates skeletal muscle oxidative capacity by regulating mitochondrial biogenesis and autophagy. Nat. Med. 2013, 19, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Mayeuf-Louchart, A.; Thorel, Q.; Delhaye, S.; Beauchamp, J.; Duhem, C.; Danckaert, A.; Lancel, S.; Pourcet, B.; Woldt, E.; Boulinguiez, A.; et al. Rev-erb-alpha regulates atrophy-related genes to control skeletal muscle mass. Sci. Rep. 2017, 7, 14383. [Google Scholar] [CrossRef] [PubMed]
- Pariollaud, M.; Gibbs, J.E.; Hopwood, T.W.; Brown, S.; Begley, N.; Vonslow, R.; Poolman, T.; Guo, B.; Saer, B.; Jones, D.H.; et al. Circadian clock component REV-ERB-alpha controls homeostatic regulation of pulmonary inflammation. J. Clin. Investig. 2018, 128, 2281–2296. [Google Scholar] [CrossRef] [PubMed]
- Griffin, P.; Dimitry, J.M.; Sheehan, P.W.; Lananna, B.V.; Guo, C.; Robinette, M.L.; Hayes, M.E.; Cedeno, M.R.; Nadarajah, C.J.; Ezerskiy, L.A.; et al. Circadian clock protein Rev-erb-alpha regulates neuroinflammation. Proc. Natl. Acad. Sci. USA 2019, 116, 5102–5107. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Mizuno, M.; Matsumoto, A.; Yang, Z.; Jimbo, E.F.; Tabata, H.; Yamagata, T.; Nagata, K.I. Role of a circadian-relevant gene NR1D1 in brain development: Possible involvement in the pathophysiology of autism spectrum disorders. Sci. Rep. 2017, 7, 43945. [Google Scholar] [CrossRef]
- Torra, I.P.; Tsibulsky, V.; Delaunay, F.; Saladin, R.; Laudet, V.; Fruchart, J.C.; Kosykh, V.; Staels, B. Circadian and glucocorticoid regulation of Rev-erb-alpha expression in liver. Endocrinology 2000, 141, 3799–3806. [Google Scholar] [CrossRef]
- Murayama, Y.; Yahagi, N.; Takeuchi, Y.; Aita, Y.; Saber, Z.M.; Wada, N.; Li, E.; Piao, X.; Sawada, Y.; Shikama, A.; et al. Glucocorticoid receptor suppresses gene expression of Rev-erb-alpha (Nr1d1) through interaction with the CLOCK complex. FEBS Lett. 2019, 593, 423–432. [Google Scholar] [CrossRef]
- Gauger, M.A.; Sancar, A. Cryptochrome, circadian cycle, cell cycle checkpoints, and cancer. Cancer Res. 2005, 65, 6828–6834. [Google Scholar] [CrossRef]
- Engelen, E.; Janssens, R.C.; Yagita, K.; Smits, V.A.; van der Horst, G.T.; Tamanini, F. Mammalian TIMELESS is involved in period determination and DNA damage-dependent phase advancing of the circadian clock. PLoS ONE 2013, 8, e56623. [Google Scholar] [CrossRef]
- Androulakis, I.P. Circadian rhythms and the HPA axis: A systems view. WIREs Mech. Dis. 2021, 13, e1518. [Google Scholar] [CrossRef]
- Talamanca, L.; Gobet, C.; Naef, F. Sex-dimorphic and age-dependent organization of 24-hour gene expression rhythms in humans. Science 2023, 379, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, W.; Lee, K.H. The role of microRNAs in the molecular link between circadian rhythm and autism spectrum disorder. Anim. Cells Syst. 2023, 27, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Costello, H.M.; Crislip, G.R.; Cheng, K.Y.; Lynch, I.J.; Juffre, A.; Bratanatawira, P.; Mckee, A.; Thelwell, R.S.; Mendez, V.M.; Wingo, C.S.; et al. Adrenal-specific KO of the circadian clock protein BMAL1 alters blood pressure rhythm and timing of eating behavior. Function 2023, 4, zqad001. [Google Scholar] [CrossRef] [PubMed]
- Horta, N.A.C.; Fernandes, P.; Cardoso, T.S.R.; Machado, F.S.M.; Drummond, L.R.; Coimbra, C.C.; Wanner, S.P.; Castrucci, A.M.L.; Poletini, M.O. TRPV1 inactivation alters core body temperature and serum corticosterone levels: Impacts on clock genes expression in the liver and adrenal glands. J. Therm. Biol. 2023, 114, 103514. [Google Scholar] [CrossRef]
- Wong, S.D.; Wright, K.P., Jr.; Spencer, R.L.; Vetter, C.; Hicks, L.M.; Jenni, O.G.; LeBourgeois, M.K. Development of the circadian system in early life: Maternal and environmental factors. J. Physiol. Anthropol. 2022, 41, 22. [Google Scholar] [CrossRef]
- Lehmann, M.; Haury, K.; Oster, H.; Astiz, M. Circadian glucocorticoids throughout development. Front. Neurosci. 2023, 17, 1165230. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Common Name | Gene Symbol | Ensembl Number |
|---|---|---|
| Circadian locomotor output cycles protein kaput | CLOCK | ENSBTAG00000044044 |
| Neuronal PAS domain protein 2 | NPAS2 | ENSBTAG00000019697 |
| Aryl hydrocarbon receptor nuclear translocator like | ARNTL | ENSBTAG00000013029 |
| Nuclear receptor subfamily 1 group D member 1 | NR1D1 | ENSBTAG00000012178 |
| Cryptochrome circadian regulator 1 | CRY1 | ENSBTAG00000010149 |
| Cryptochrome circadian regulator 2 | CRY2 | ENSBTAG00000021223 |
| Period circadian regulator 1 | PER1 | ENSBTAG00000003889 |
| Period circadian regulator 2 | PER2 | ENSBTAG00000001879 |
| PVN | CLOCK | NPAS2 | ARNTL | NR1D1 | CRY1 | CRY2 | PER1 | PER2 |
| CLOCK | - | −0.737 | 0.421 | 0.356 | −0.256 | 0.125 | −0.646 | 0.006 |
| NPAS2 | 0.037 | - | −0.643 | −0.769 | 0.337 | −0.558 | 0.174 | −0.434 |
| ARNTL | 0.299 | 0.085 | - | 0.447 | −0.778 | 0.316 | −0.442 | −0.185 |
| NR1D1 | 0.386 | 0.026 | 0.267 | - | −0.173 | 0.852 | 0.215 | 0.548 |
| CRY1 | 0.540 | 0.414 | 0.023 | 0.681 | - | −0.091 | 0.298 | 0.349 |
| CRY2 | 0.767 | 0.151 | 0.446 | 0.007 | 0.830 | - | 0.265 | 0.323 |
| PER1 | 0.084 | 0.680 | 0.273 | 0.609 | 0.474 | 0.527 | - | 0.652 |
| PER2 | 0.989 | 0.283 | 0.662 | 0.160 | 0.397 | 0.435 | 0.080 | - |
| PIT | CLOCK | NPAS2 | ARNTL | NR1D1 | CRY1 | CRY2 | PER1 | PER2 |
| CLOCK | - | −0.595 | 0.071 | −0.414 | 0.710 | 0.285 | −0.208 | −0.223 |
| NPAS2 | 0.120 | - | 0.297 | 0.892 | −0.663 | −0.718 | −0.298 | −0.232 |
| ARNTL | 0.868 | 0.475 | - | −0.025 | 0.449 | −0.680 | −0.723 | −0.614 |
| NR1D1 | 0.308 | 0.003 | 0.953 | - | −0.789 | −0.471 | −0.065 | −0.066 |
| CRY1 | 0.048 | 0.073 | 0.263 | 0.020 | - | 0.154 | −0.349 | −0.230 |
| CRY2 | 0.494 | 0.045 | 0.063 | 0.239 | 0.716 | - | 0.830 | 0.701 |
| PER1 | 0.620 | 0.474 | 0.043 | 0.878 | 0.397 | 0.011 | - | 0.818 |
| PER2 | 0.596 | 0.581 | 0.106 | 0.877 | 0.584 | 0.053 | 0.013 | - |
| AMYG | CLOCK | NPAS2 | ARNTL | NR1D1 | CRY1 | CRY2 | PER1 | PER2 |
| CLOCK | - | −0.670 | 0.813 | −0.331 | 0.537 | 0.917 | −0.120 | 0.171 |
| NPAS2 | 0.054 | - | −0.741 | −0.214 | −0.251 | −0.821 | −0.173 | −0.364 |
| ARNTL | 0.014 | 0.035 | - | −0.162 | 0.458 | 0.854 | 0.058 | 0.500 |
| NR1D1 | 0.424 | 0.611 | 0.701 | - | −0.215 | −0.173 | 0.246 | 0.570 |
| CRY1 | 0.170 | 0.548 | 0.253 | 0.610 | - | 0.277 | 0.055 | 0.013 |
| CRY2 | 0.001 | 0.012 | 0.007 | 0.681 | 0.506 | - | −0.079 | 0.351 |
| PER1 | 0.778 | 0.682 | 0.891 | 0.558 | 0.896 | 0.852 | - | −0.157 |
| PER2 | 0.686 | 0.375 | 0.207 | 0.140 | 0.976 | 0.394 | 0.710 | - |
| ADC | CLOCK | NPAS2 | ARNTL | NR1D1 | CRY1 | CRY2 | PER1 | PER2 |
| CLOCK | - | −0.577 | 0.625 | −0.494 | −0.052 | 0.362 | 0.100 | 0.197 |
| NPAS2 | 0.135 | - | −0.540 | 0.636 | 0.145 | −0.366 | 0.016 | 0.485 |
| ARNTL | 0.098 | 0.167 | - | −0.738 | 0.421 | 0.377 | −0.363 | −0.291 |
| NR1D1 | 0.213 | 0.090 | 0.037 | - | −0.198 | −0.148 | 0.149 | −0.329 |
| CRY1 | 0.902 | 0.732 | 0.299 | 0.638 | - | 0.021 | 0.217 | 0.734 |
| CRY2 | 0.379 | 0.372 | 0.357 | 0.726 | 0.961 | - | 0.691 | 0.561 |
| PER1 | 0.815 | 0.377 | 0.969 | 0.725 | 0.606 | 0.058 | - | 0.649 |
| PER2 | 0.640 | 0.485 | 0.223 | 0.426 | 0.038 | 0.148 | 0.081 | - |
| ADM | CLOCK | NPAS2 | ARNTL | NR1D1 | CRY1 | CRY2 | PER1 | PER2 |
| CLOCK | - | −0.049 | 0.469 | −0.237 | −0.206 | −0.243 | −0.301 | −0.450 |
| NPAS2 | 0.909 | - | −0.405 | −0.703 | −0.365 | −0.762 | −0.510 | −0.332 |
| ARNTL | 0.241 | 0.320 | - | −0.003 | 0.283 | 0.422 | 0.164 | 0.277 |
| NR1D1 | 0.571 | 0.052 | 0.995 | - | 0.066 | 0.395 | 0.560 | 0.402 |
| CRY1 | 0.625 | 0.374 | 0.497 | 0.876 | - | 0.438 | 0.095 | 0.341 |
| CRY2 | 0.563 | 0.028 | 0.298 | 0.333 | 0.278 | - | 0.742 | 0.564 |
| PER1 | 0.468 | 0.197 | 0.697 | 0.149 | 0.822 | 0.035 | - | 0.807 |
| PER2 | 0.264 | 0.422 | 0.506 | 0.324 | 0.409 | 0.145 | 0.015 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Earnhardt-San, A.L.; Baker, E.C.; Riley, D.G.; Ghaffari, N.; Long, C.R.; Cardoso, R.C.; Randel, R.D.; Welsh, T.H., Jr. Differential Expression of Circadian Clock Genes in the Bovine Neuroendocrine Adrenal System. Genes 2023, 14, 2082. https://doi.org/10.3390/genes14112082
Earnhardt-San AL, Baker EC, Riley DG, Ghaffari N, Long CR, Cardoso RC, Randel RD, Welsh TH Jr. Differential Expression of Circadian Clock Genes in the Bovine Neuroendocrine Adrenal System. Genes. 2023; 14(11):2082. https://doi.org/10.3390/genes14112082
Chicago/Turabian StyleEarnhardt-San, Audrey L., Emilie C. Baker, David G. Riley, Noushin Ghaffari, Charles R. Long, Rodolfo C. Cardoso, Ronald D. Randel, and Thomas H. Welsh, Jr. 2023. "Differential Expression of Circadian Clock Genes in the Bovine Neuroendocrine Adrenal System" Genes 14, no. 11: 2082. https://doi.org/10.3390/genes14112082