Abstract
Malate dehydrogenase (MDH) is one kind of oxidation–reduction enzyme that catalyzes the reversible conversion of oxaloacetic acid to malic acid. It has vital functions in plant development, photosynthesis, abiotic stress responses, and so on. However, there are no reports on the genome-wide identification and gene expression of the MDH gene family in Arachis hypogaea. In this study, the MDH gene family of A. hypogaea was comprehensively analyzed for the first time, and 15 AhMDH sequences were identified. According to the phylogenetic tree analysis, AhMDHs are mainly separated into three subfamilies with similar gene structures. Based on previously reported transcriptome sequencing results, the AhMDH expression quantity of roots and leaves exposed to manganese (Mn) toxicity were explored in A. hypogaea. Results revealed that many AhMDHs were upregulated when exposed to Mn toxicity, suggesting that those AhMDHs might play an important regulatory role in A. hypogaea’s response to Mn toxicity stress. This study lays foundations for the functional study of AhMDHs and further reveals the mechanism of the A. hypogaea signaling pathway responding to high Mn stress.
1. Introduction
MDH (malate dehydrogenase) is one of the oxidation–reduction enzymes widely present in plants, animals, and microorganisms, and it has highly conserved properties [1]. MDH relies on the NADH/NADPH cofactor to participate in the biochemical reaction of the reversible conversion of oxaloacetic acid to malic acid [2,3]. According to the different cofactors on which MDH depends, MDH contains two categories: NAD-MDH and NADP-MDH, among which NAD-MDH is mainly present in the cytoplasm, mitochondria, glyoxysome, peroxidation object, and chloroplast, while NADP-MDH only exists in the chloroplast [4,5]. MDH can participate in the regulation of oxaloacetate changing to malate and NAD+ to NADH, providing reducing power for the nitric acid reduction process [6,7]. NADH produced through biological oxidation in the cytosol cannot pass through the inner mitochondrial membrane [8,9]. However, NADH can deliver hydrogen to oxaloacetic acid under the action of MDH to produce malic acid, which can be transported to the mitochondrial matrix through the malate–aspartate shuttle system [8,9]. Furthermore, the malic acid in the mitochondrial matrix is reoxidized to oxaloacetic acid and NADH and then oxidized through the respiratory chain [8,9]. NADP-MDH is a light-activated enzyme participating in the regulation of C4 photosynthesis, and NADP-MDH is also light-activated in C3 plants; however, NADP-MDH is relatively inactive in the dark [10].
Plant MDH is involved in a variety of metabolic pathways, such as the tricarboxylic acid cycle, the glyoxylic acid cycle, amino acid metabolism, redox equilibrium, and gluconeogenesis [11]. MDH is closely associated with different organelles through the malic acid shuttle system [12,13]. The malic acid in cells is indirectly transferred to different organelles, thus maintaining the physiological and biochemical metabolism balance in plants and playing an important part in the exchange of substances between the organelles and cytosol [12,13]. Furthermore, many scientific studies reports have shown that plant MDHs play vital parts in seed growth, photosynthesis and coping with abiotic stress and biological stress [14].
With the sequencing of whole genomes and the development of bioinformatics, many types of MDHs have been identified in different plants: 12 MDHs have been found in rice (Oryza sativa) [15], 16 MDHs have been discovered in soybean (Glycine max) [16], 12 MDHs have been found in Chinese fir (Cunninghamia lanceolata) [13], 13 MDHs have been identified in cotton (Gossypium hirsutum) [17], 7 MDHs have been detected in Stylosanthes (Stylosanthes guianensis) [18], 20 MDHs have been discovered in apple (Malus pumila) [19], and 16 MDH genes have been found in poplar (Populus trichocarpa) [20].
Peanut (Arachis hypogaea) is one of the main oil crops in tropical and subtropical areas [21]. It provides edible oil and edible protein for people worldwide and accounts for 30% of China’s total oilseed production [22]. Peanut is rich in protein, fat, carbohydrates, and vitamins, with a fat content of 44–45% and a protein content of 24–36%, making it one of the most widely consumed nuts in China [23,24]. The nutritional value of peanut is high, and it is easily absorbed and utilized by the human body, making it comparable to animal foods such as eggs, milk, and lean meat [25].
Seeds with vigorous vitality and moderate dormancy guarantee seedling size and seedling strength after planting and directly affect peanut yield [26]. During the process of water absorption and the expansion of seeds, mitochondrial respiration is rapidly enhanced, malate dehydrogenase activity increases, and stored substances in seeds are mobilized to provide energy for seed germination [27]. The decreased activity of malate dehydrogenase and cytochrome C oxidase may lead to an insufficient mitochondrial energy supply and the accumulation of reactive oxygen species (ROS), leading to decreased or even a loss of seed vitality [28,29]. Some research reports have found that AhMDH1 encodes mitochondrial malate dehydrogenase, which is a hydrophobic protein, and the gene expression is strongly linked with the water absorption process during seed germination, indicating that MDH is necessary for the mobilization of storage substances during seed germination [30].
Manganese (Mn) is an essential element in almost all living organisms and can play different functional roles in plants, either as an enzyme cofactor or as a catalytically active metal in biological clusters [31]. Mn plays vital parts in photosynthesis, respiration, protein biosynthesis, and hormone activation in plants [32]. When the available Mn content in the soil exceeds the normal Mn concentration needed by a plant, the plant is likely to suffer from Mn toxicity. Plants, as a result, display obvious signs of poisoning on their leaves, such as dark patches, crinkled leaves, and chlorosis; Mn poisoning also affects roots, resulting in reduced lateral root count and dry weight, in addition to the signs on the leaves [33]. Excess Mn prevents the absorption and transfer of other essential elements, such as calcium, magnesium, iron, and phosphorus, possibly because of the similar ionic radii or binding strength of ligands [34]. At the molecular level, excess Mn can prevent the absorption and transfer of other essential elements, such as calcium, magnesium, iron, and phosphorus [35], and inhibit the synthesis of chlorophyll [34], resulting in a decrease in the rate of photosynthesis [36]. The transcriptome sequencing results of peanut roots and leaves treated with 10 and 300 μM of manganese sulfate show that the peanut MDH gene family was strongly induced by manganese toxicity stress, and the expression levels changed significantly [37]; these results need to be further verified through fluorescent quantitative PCR. However, the role of the MDH gene family in Mn toxicity stress in peanut has not been reported.
The MDH gene in the peanut genome has not yet been fully elucidated, and its function needs to be verified. In this study, AhMDHs in the genome of peanut were identified using multi-analysis. The analysis of the basic information of AhMDHs is helpful to elucidate the molecular regulation mechanisms of peanut development and response to Mn toxicity stress, providing a theoretical grounding for the molecular breeding of peanut.
2. Materials and Methods
2.1. Analysis of AhMDH Structures
The peanut genome was sourced from the website of National Center of Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/) (accessed on 1 October 2023), and the transcriptome data (NCBI accession number: PRJNA901194) [37] were used for MDH gene identification. Genes including the domain of MDH were called MDH1 to MDH15, and genes that matched the published AhMDH1 sequence [30] were named AhMDH1. The website of global species databases (GSDs) (http://gsds.gao-lab.org/) (accessed on 2 October 2023) was used based on the annotated information for the peanut genome gff. The structural model of the MDH in peanut was visualized. In this study, TBtools (Toolkit for Biologists integrating various biological data handling tools) [38] was adopted to visualize the structure and intuitively draw the predicted results.
2.2. Constructing a Phylogeny Tree of the MDHs
MEGA11 (molecular evolutionary genetics analysis version 11) software [39] was adopted to compare the amino acid sequences of the MDH family of peanut with those of Arabidopsis thaliana, Glycine max, Phaseolus vulgaris, and Stylosanthes guianensis. Maximum likelihood estimate (MLE) was used to construct a phylogeny tree of the MDHs. Through phylogeny exploration of the peanut MDHs and other species, bootstrap analysis was used to build a neighbor-joining (NJ) tree with 1000 repeats. The cutoff value for site coverage was 60%. According to a previous report [40], EOT (Evolview online tools) (https://www.evolgenius.info/evolview/#/treeview) (accessed on 5 October 2023) were used to construct a phylogenetic tree.
2.3. Chromosome Localization of the MDH Gene
According to the peanut genome annotation information downloaded from NCBI, the positions of 15 peanut MDH genes on chromosomes were obtained, and TBtools software V1.098 was used to locate the chromosomes [38].
2.4. MDH Conservative Motif Analysis
The protein sequence of peanut MDH was presented to the MEME (Multiple Em for Motif Elicitation) (http://meme-suite.org/) (accessed on 6 October 2023) [41] took for the prediction of motifs. The discovery number of the motif displayed was 10; the motif width was set to 6–50. TBtools was used to visualize the MDH motif domain of peanut.
2.5. Cis-Acting Element Analysis
Based on the previously reported peanut genome information [20], peanut MDH gene sequences upstream of 2 kb were presented to Plant Care websites (HTML/http://bioinformatics.psb.ugent.be/webtools/plantcare/) (accessed on 8 October 2023) on cis-acting element prediction [42]. The Plant Care analysis results were simplified and visualized using TBTools.
2.6. Transcriptome Sequencing Results of AhMDHs in Peanut Roots and Leaves under Manganese Toxicity Stress
The results of the AhMDHs in peanut roots and leaves under manganese toxicity stress treatment were derived from RNA sequencing [37]. FPKM (fragments per kilobase of transcript per million mapped reads) values (shown in Table S1) were used to reflect gene expression [43], using the bioinformatics website (http://www.bioinformatics.com.cn/plot_basic_cluster_heatmap_plot_024) (accessed on 9 October 2023) for data visualization.
2.7. Peanut Culture Conditions
The peanut variety used was called ZY-62 (Zhanyou 62); it was cultivated by the ZAAS (Zhanjiang Academy of Agricultural Sciences), Guangdong Province, China. The peanut seeds were germinated in quartz sand and cultured for eight days before being treated with manganese. Then, the uniformly growing seedlings of peanut were transferred to plastic incubators with added nutrient solution (volume of 15 L) for water planting [44]. In short, the nutrient solution contained 1500 µM KNO3, 400 µM NH4NO3, 25 µM MgCl2, 1200 µM Ca(NO3)2·4H2O, 40 µM Fe-EDTA(Na), 500 µM MgSO4·7H2O, 0.16 µM (NH4)5MoO24·4H2O, 300 µM K2SO4, 300 µM (NH4)2SO4, 0.5 µM CuSO4·5H2O, 1.5 µM ZnSO4·7H2O, 500 µM KH2PO4, and 2.5 µM NaB4O7·10H2O. All chemicals used were of analytical grade (Kemer, Tianjin, China). Simultaneously, 10 and 300 µM manganese sulfate (MnSO4) [37] (Kemer, China) were supplemented to the nutrient solution for manganese treatment. The control group consisted of a nutrient solution containing 10 µM Mn, while the manganese toxicity treatment group contained 300 µM Mn. Every experiment was conducted in 4 biological repetitions. The temperature used to regulate plant development was at 24–29 °C (day)/19–23 °C (night). The period of light was approximately 12 h/d, the light intensity was 2000 lux, and the nutrient solution was renewed every five days. The pH value in the nutrient solution was adjusted to 5.0 with 1 M KOH or H2SO4 every two days (Kermel, Tianjin, China).
2.8. qRT-PCR Analysis of MDH Gene Expression in Peanut
When the peanut seedlings were 20 days old, the roots and leaves were collected, and total RNA was extracted from the peanut leaves and roots using a ribonucleic acid extraction kit (Yisheng, Shanghai, China). After the removal of gDNA (genomic DNA), cDNA was formed by adopting a reverse-transcription kit (Yisheng, China). For conducting quantitative RT-PCR (qRT-PCR) exploration, a fluorescence ration PCR instrument (CFX Connect Optics Module, Bio-Rad, Hercules, CA, USA) was used [37]. In brief, the samples were thinned 30 times for qRT-PCR. The reaction procedure was performed at 95 °C for 30 s, followed by 95 °C for 5 s, 60 °C for 15 s, and 72 °C for 30 s. The internal control gene named AhUbiquitin (DQ887087.1) was used to calculate the relativity transcription level based on the transcription ratio of the AhMDHs to the AhUbiquitin. The primers used for qRT-PCR detection are listed in Table S2.
3. Results
3.1. Analysis of Basic Information of AhMDHs
All of the 15 studied AhMDHs were identified from the whole genome of A. hypogaea; 3 of the genes were located on chromosome 09; 2 genes were located on chromosomes 03, 12, and 19; and the other 6 genes were distributed on chromosomes 01, 02, 05, 10, 11, and 13 (Table 1). The CDS length of the AhMDHs was approximately 1000 bp, among which AhMDH2 was the longest, 1359 bp, and its encoded protein was 452 amino acids (Table 1). The relative molecular mass of the AhMDH protein ranged from 35.43 to 47.91 kDa, and the isoelectric points ranged from 6.01 to 9.00. Fifteen AhMDHs were predicted; the results showed that AhMDH1 and AhMDH10 were located only in the mitochondrion, and AhMDH3, AhMDH9, and AhMDH14 were located only in the chloroplast. The remaining 10 AhMDHs were located both in mitochondria and chloroplasts (Table 1).

Table 1.
Summary of basic information of AhMDHs.
3.2. Structural Analysis of AhMDHs
All 15 MDH genes (AhMDH1–AhMDH15) were verified in the genome of peanut (Table 1). After analyzing their genetic structure, the research findings displayed that most genes are between 3 kb and 4 kb in length, and most contain more than six exons. The domain prediction revealed that all of the genes contained PLN (plant-specific proteins) domains, and only the number of PLN domains was different (Figure 1).
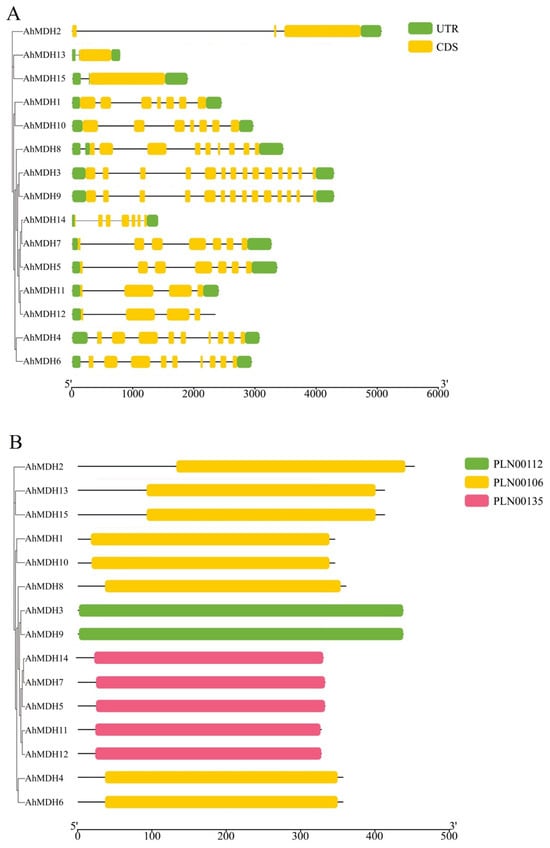
Figure 1.
Structure and domain analysis of peanut MDH. (A) Genetic structure; (B) domain analysis. Conserved protein domain families PLN00112, 00106, and 00135 are classified as models that may span more than one domain.
3.3. Evolutionary Tree Analysis of MDHs
MEGA11 was used to construct NJ phylogenetic trees of 55 MDH genes from Arachis hypogaea, Glycine max, Arabidopsis thaliana, Phaseolus vulgaris, and Stylosanthes guianensis. On the basis of the phylogeny tree, peanut MDH was distinguished from three subfamilies (I, II, and III), and the MDH of peanut and Arabidopsis thaliana had cross-distribution in subfamilies I and III (Figure 2). There were at most 27 proteins in subfamily III and at least 5 proteins in subfamily II, whereas peanut MDH proteins were only distributed in subfamilies I and III (Figure 2).
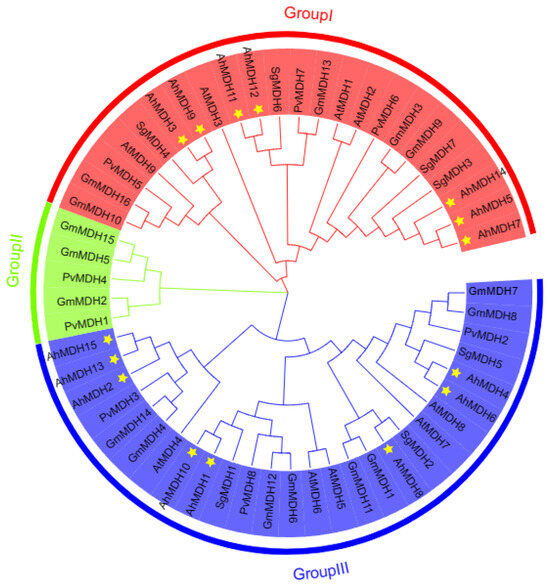
Figure 2.
Phylogenetic tree of MDH in Arachis hypogaea, Glycine max, Arabidopsis thaliana, Phaseolus vulgaris, and Stylosanthes guianensis. Phaseolus vulgaris: PvMDH1 (XP_007157340.1), PvMDH2 (XP_007159190.1), PvMDH3 (XP_007155306.1), PvMDH4 (XP_007155646.1), PvMDH5 (XP_007144250.1), PvMDH6 (XP_007145851.1), PvMDH7 (XP_007134537.1), and PvMDH8 (XP_007133189.1); Arabidopsis thaliana: AtMDH1 (At1g04410), AtMDH2 (At5g43330), AtMDH3 (At5g56720), AtMDH4 (At3g47520), AtMDH5 (At3g15020), AtMDH6 (At1g53240), AtMDH7 (At2g22780), AtMDH8 (At5g09660), and AtMDH9 (At5g58330); Stylosanthes guianensis: SgMDH1 (KJ123727), SgMDH2 (OK188912), SgMDH3 (OK188913), SgMDH4 (OK188914), SgMDH5 (OK188915), SgMDH6 (OK188916), and SgMDH7 (OK188917); Glycine max: GmMDH1 (Glyma.01G197700), GmMDH2 (Glyma.01G210400), GmMDH3 (Glyma.02G005500), GmMDH4 (Glyma.05G026300), GmMDH5 (Glyma.05G058100), GmMDH6 (Glyma.06G231500), GmMDH7 (Glyma.07G185400), GmMDH8 (Glyma.08G063800), GmMDH9 (Glyma.10G006500), GmMDH10 (Glyma.10G197700), GmMDH11 (Glyma.11G043900), GmMDH12 (Glyma.12G159300), GmMDH13 (Glyma.13G104800), GmMDH14 (Glyma.17G100600), GmMDH15 (Glyma.17G140600), and GmMDH16 (Glyma.20G192200); Arachis hypogaea: AhMDH1 (LOC112790043), AhMDH2 (LOC112718423), AhMDH3 (LOC112776877), AhMDH4 (LOC112791339), AhMDH5 (LOC112711210), AhMDH6 (LOC112728667), AhMDH7 (LOC112776383), AhMDH8 (LOC112800443), AhMDH9 (LOC112711491), AhMDH10 (LOC112737907), AhMDH11 (LOC112735613), AhMDH12 (LOC112726743), AhMDH13 (LOC112710233), AhMDH14 (LOC112711212), and AhMDH15 (LOC112722502). Yellow asterisks indicate the peanut MDH gene.
3.4. Conservative Analysis of Peanut MDH
A conservative motif analysis of peanut MDH sequences was performed using the MEME tool. The amount of MDH motifs in the peanut was basically 8–9 (Figure 3). All genes had motifs 1, 2, 5, and 6, in which motif 1 encoded 41 amino acids, motif 2 encoded 50 amino acids, motif 5 encoded 21 amino acids, and motif 6 encoded 21 amino acids. The amino acid coding sequences created by these four motifs were the key motifs of MDH. Twelve genes contained motif 4, ten genes contained motif 8, and eight genes included motifs 3, 7, 9, and 10. Thus, these domains may play an important role.
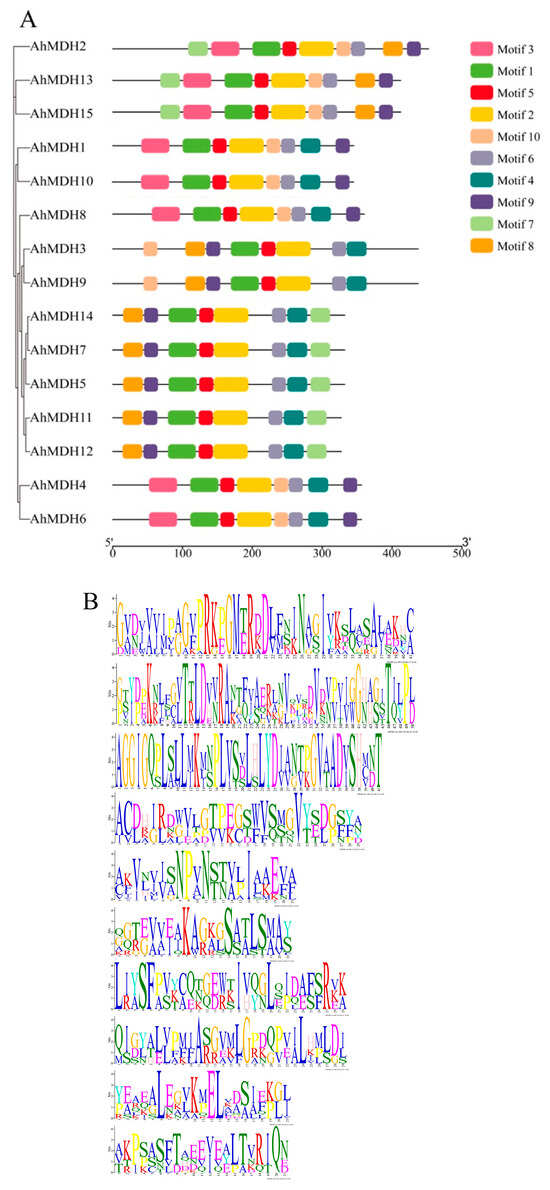
Figure 3.
Analysis of MDH gene domain in peanut. (A) Domain analysis; different colors represent different motifs, and all genes have motif 1, motif 2, motif 5, and motif 6 domains. (B) Motifs analysis; the values are motif 1–motif 10 from top to bottom. Different lengths represent the length of the encoded amino acid.
3.5. Chromosome Location of Peanut MDH
Chromosome mapping of the 15 peanut MDH genes was performed (Figure 4). Most of the genes were located in regions where genes were dense (i.e., yellow), while AhMDH5, AhMDH14, AhMDH15, and AhMDH7 were located in regions where genes were dispersed (blue). According to the results, three genes were situated on chromosome 09, two genes were situated on chromosomes 03, 12, and 19, and only one MDH gene was located on another chromosome. Nine genes were located between 90 and 150 Mb, four genes were located between 0 and 30 Mb (AhMDH11, AhMDH8, AhMDH2, and AhMDH12), and the remaining two genes were located between 30 and 60 Mb (AhMDH1 and AhMDH10).
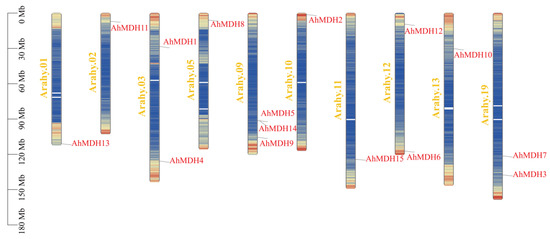
Figure 4.
Chromosome location of MDH genes in peanut. Red represents genes and luminous yellow represents chromosome numbers. AhMDH1, AhMDH2, AhMDH3, AhMDH4, AhMDH6, AhMDH8, AhMDH9, AhMDH10, AhMDH11, AhMDH12, and AhMDH13 were located in regions where genes were dense (yellow), while AhMDH5, AhMDH14, AhMDH15, and AhMDH7 were located in regions where genes were dispersed (blue).
3.6. Identification of Cis-Acting Factors of the AhMDHs
The length of 2 kb upstream sequences was selected, respectively, from the promoter of 15 AhMDHs. The cis-acting elements of the AhMDHs promoters were predicted using the PlantCARE online tool. All peanut MDH genes contained 3–11 photoresponsive elements (Figure 5). In addition, the promoter region of the MDH gene in peanut mainly consisted of 46 MeJA (jasmonic acid methyl ester) response elements, 31 anaerobic induction factors, and 30 ABA (abscisic acid) response factors. In addition, there were 20 MYB binding site response elements and a few regulatory factors, such as auxin response factors, salicylic acid response factors, low-temperature response factors, endosperm expression factors, meristem expression factors, erythromycin response factors, zein metabolism regulation elements, stress and defense response factors, circadian rhythm control factors, and cell cycle regulation factors (Figure 5).
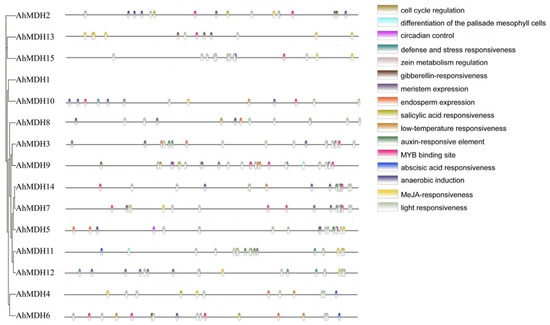
Figure 5.
Analysis of cis-acting elements of peanut MDH gene. Homeopathic component distribution.
3.7. Expression Profile of the Peanut MDH Gene
To study the expression profiles of AhMDHs, the quantitative analysis of 15 AhMDHs expression in roots and leaves treated with Mn were detected through transcriptome sequencing [37]. The experimental results revealed that MDH had a differential expression in both the roots and leaves of peanut (Figure 6). As shown in the Figure 6, AhMDH4, AhMDH3, AhMDH6, AhMDH7, AhMDH9, AhMDH5, AhMDH1, and AhMDH14 were mainly expressed in the leaves, whereas AhMDH12, AhMDH11, AhMDH2, AhMDH13, and AhMDH10 were mainly expressed in the roots. Among them, AhMDH1, AhMDH8, and AhMDH14 were highly expressed in leaves treated with 300 μM Mn, AhMDH13 was highly expressed in roots treated with 300 μM Mn, AhMDH14 was not expressed in normal leaves, and AhMDH15 was not expressed in leaves treated with 300 μM Mn. Overall, 15 AhMDHs were expressed differentially in both peanut leaves and roots.
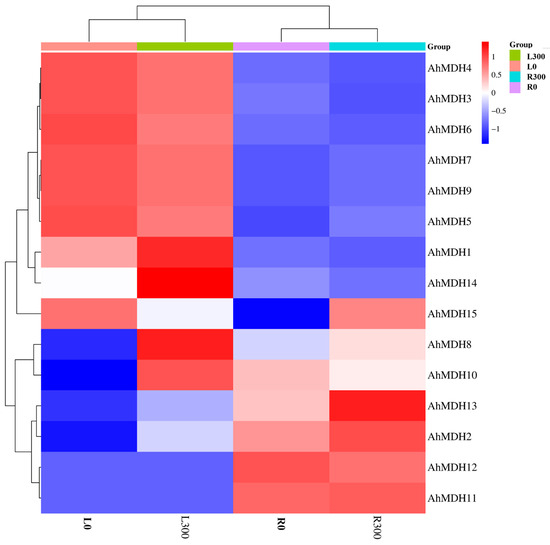
Figure 6.
Expression analysis of AhMDHs in roots and leaves of peanut treated with manganese detected through transcriptome sequencing. L300: Leaves treated with 300 μM Mn. L0: Leaves treated with 10 μM Mn (control group). R300: Roots treated with 300 μM Mn. R0: Roots treated with 10 μM Mn (control group).
3.8. AhMDH Gene Expression Analysis under Mn Stress
AhMDHs in peanut leaves and roots after Mn treatment were analyzed using qRT-PCR, and the results of the gene expression of those 15 AhMDHs in roots and leaves after Mn treatment were further verified (Figure 7). All 15 genes showed different degrees of tissue-specific expression in all tissues. When peanuts sustained Mn toxicity stress, they displayed various expression patterns in roots and leaves. All 15 AhMDHs in the peanut roots were significantly upregulated or downregulated. Nevertheless, the expression of AhMDH7, AhMDH9, AhMDH11, and AhMDH13 in leaves under Mn toxicity stress had no significant difference.
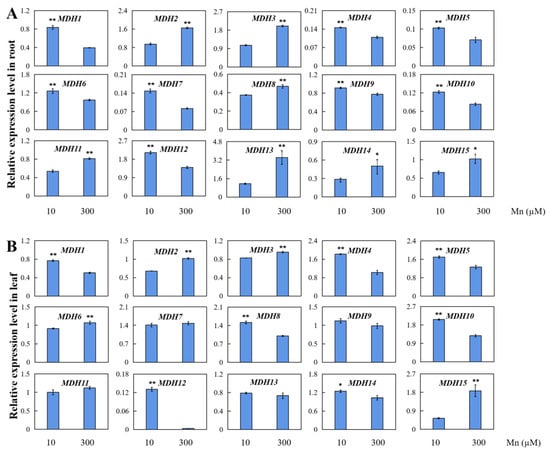
Figure 7.
qRT-PCR detection of 15 AhMDHs in peanut leaves and roots at concentrations of 10 and 300 µM manganese. Relative expression levels of AhMDHs in peanut (A) roots and (B) leaves. The data are represented by the mean and standard deviation of the four experimental replicates. T-test was used to assess the difference between control group and manganese poisoning (* p < 0.05, ** p < 0.01).
4. Discussion
With the development of bioinformatics technology, an increasing number of MDH members have been authenticated in plants, such as Oryza sativa [15], Glycine max [45], and Stylosanthes guianensis [18]. Eight MDH genes in beans (Phaseolus vulgaris) have been identified and found to be mainly located in the cytoplasm and chloroplasts [1]. Seven SgMDHs were found in stylosanthes, and subcellular localization showed that SgMDH mainly exists in the cytoplasm, chloroplasts, peroxidase bodies, and vacuolar membranes [18]. In addition, 12 OsMDHs have been confirmed in O. sativa [15], and 12 SlMDHs have also been verified in tomato (Solanum lycopersicum) [17]. Twenty MpMDHs have been identified in the whole genome of apple (Malus pumila) [18], and studies have shown that the number of MDHs is different in different species of plants and that the MDHs are distributed in different subregions of plant cells. In this study, 15 AhMDHs were identified in peanut, and they were all located in the mitochondria and chloroplasts. Chloroplasts and mitochondria are important organelles for energy metabolism [46]. This indicates that peanut AhMDHs might participate in regulating energy metabolism, regulating peanut growth, and responding to abiotic stress.
The analysis results of conservative motifs showed that there were basically 8–9 conserved motifs in all MDH proteins (Figure 4), indicating that the AhMDH protein has a highly conservative protein composition. Those findings agree with the result of MDH in poplar (Populus trichocarpa) [19] and cotton (Gossypium hirsutum) [47], suggesting that the MDHs are relatively conservative in the long process of natural selection and species evolution. The comparison with MDHs in various plants showed that MDHs experience wide extension during evolution [14,15].
AhMDH cis-acting factor interpretation revealed different plant development factors and stress response factors (Figure 5). Many cis factors were found in the AhMDHs, such as MYB binding site respond factors, auxin respond factors, SA (salicylic acid) respond factors, low-temperature respond factors, endosperm expression factors, meristem expression elements, erythromycin response elements, zein metabolism regulation elements, stress and defense respond factors, circadian rhythm control factors, and cell cycle regulation factors. This suggests that AhMDHs play a vital regulatory part in response to hormone synthesis, plant growth, biotic and abiotic stresses, and so on.
Gene expression profiles provide a crucial basis for confirming the function of genes [48]. A few MDHs have been proven to be specifically expressed in some plant tissues and to play vital roles in the development of seeds [49], root growth [50], light respiration [51], and stress resistance [46]. In the present study, various expression profiles of AhMDHs were discovered in the roots and leaves of peanut. Furthermore, all of the AhMDHs in peanut leaves and roots were found to be differentially expressed. These findings imply that AhMDHs may play a vital regulatory role in peanut root and leaf development.
Plant MDHs are essential for seed development, photosynthesis, abiotic stress tolerance, and biological stress response [14]. MDH also plays a crucial part in plants under heavy-metal stress [15,52]. When plants are subjected to heavy-metal stress, MDH can catalyze the change from oxaloacetic acid to malate and increase the content of malate [15,52]. Some studies indicate that organic acids such as malate can interact with heavy metals to transform them into non-poisonous or low-toxicity combined states, decreasing the poisonous results of heavy metals in plants [18,52]. It has been found that MDH is involved in the differential regulation of various metal ions [53]. Studies on the leguminous crop Stylosanther showed that, under the stress of different heavy metals (iron, manganese, cuprum, zinc, aluminum, cadmium, and lanthanum), SgMDH expression levels were different, indicating that SgMDH has different response functions under different metal stresses [18]. A study on SgMDH1 in S. guianensis showed that the plant can alleviate manganese stress in soil by enhancing the content of endogenous malic acid and promoting the secretion of malic acid, thus enhancing the Mn tolerance of the plant [54]. Some research reports suggest that rice OsMDH may enhance plant stress resistance through the metabolic pathway of salicylic acid [14], and the overexpression of OsMDH can improve the stress tolerance of rice to iron [55]. mMDH2 participates in cadmium toxic stress in A. thaliana, and the mMDH2 regulates cadmium resistance negatively by regulating ROS (reactive oxygen species)-mediated signaling and ROS content [56]. Further studies have found that Arabidopsis plants overexpressing mMDH2 show a tolerant phenotype under iron-deficient conditions, and it is suggested that mMDH2 may mediate the expression of NAS4 to indirectly respond to iron deficiency stress [57]. In plants, manganese is essential for respiration, protein synthesis, hormone biosynthesis, and photosynthesis [32]. Mn toxicity occurs when the available Mn content in the soil exceeds the typical Mn concentration required by the plant. Plants show evident evidence of poisoning on their leaves as a consequence, such as dark spots, crinkled leaves, and chlorosis; Mn poisoning also affects roots, resulting in a decrease in lateral root count and dry weight, in addition to the signs found on the leaves [33]. The findings in this study revealed that AhMDHs are differentially expressed in the roots and leaves of peanut subjected to Mn toxicity stress. These results indicated that AhMDHs might play a significant regulatory role in peanut when responding to Mn toxicity stress. The specific molecular regulatory mechanisms involved are unclear and require further research.
5. Conclusions
In the present study, 15 AhMDHs in the genome of peanut were identified using multi-analysis. The physical and chemical properties, gene structure, phylogeny, and gene expression modes of AhMDHs were verified. The phyletic evolution of the 15 AhMDHs were separated into three categories. Based on the phylogeny tree interpretation, AhMDHs were only included in groups I and III, implying that those peanut MDH genes might play different roles in comparison with MDH in other species. The conservative motifs, protein interaction regulatory networks, and gene structure of AhMDHs were of high conservation, indicating that they were functionally conserved.
The cis-element analysis of the AhMDH gene revealed that all 15 AhMDHs were participated in regulation of stress response, hormone synthesis, and photosynthesis. The gene expression profiles of the AhMDHs based on transcriptome sequencing results revealed various gene expression patterns in the peanut roots and leaves. qRT-PCR testing results demonstrated that AhMDH expressions in peanut roots and leaves were upregulated or downregulated under high Mn stress. These consequences lay the foundations for the thorough elucidation of AhMDH functions in peanut signal transduction in response to high Mn stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14122109/s1, Table S1: FPKM values of AhMDHs in peanut roots and leaves under manganese toxicity stress treatment derived from RNA sequencing; Table S2: Primers of AhMDHs family for qRT-PCR detection.
Author Contributions
Y.L., S.Y. and Y.X. conceived and designed the experiments. Y.L., M.Z., J.S., S.Y. and Y.X. performed the experiments. Y.L., M.Z., J.S., S.Y. and Y.X. analyzed the data. Y.L., M.Z., S.Y. and Y.X. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Project of Marine Young Talent Innovation of Zhanjiang City (210818004542097), the Project of Rural Science and Technology Team “One-to-One” Service to Help the Town Full Coverage of Action Funding in Zhanjiang City (A23081), the Postgraduate Education Innovation Project of Guangdong Ocean University-Excellent Dissertation Cultivation Project (040502752327), the Project of Social Services of Rural Science and Technology Specialists of Guangdong Ocean University (080503052212), the National Natural Science Foundation of China (32002131), and the Maintenance Expenses of Key Construction Disciplines in High-level University of Crop Science (080508052201).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yıldız, S.; Okay, A.; Büyük, İ. Defining the roles of PvMDH genes in response to salt stress and detailed characterization of the gene family. J. Plant Biochem. Biot. 2021, 31, 380–393. [Google Scholar] [CrossRef]
- Matthews, C.J.; Andrews, E.S.V.; Patrick, W.M. Enzyme-based amperometric biosensors for malic acid—A review. Anal. Chim. Acta 2021, 1156, 338218. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, J.H.; Chen, Y.R.; Xu, Q.; Song, P.; Li, Y.F.; Li, K.; Liu, H. Recent advances in microbial production of L-malic acid. Appl. Microbiol. Biot. 2022, 106, 7973–7992. [Google Scholar] [CrossRef]
- Minárik, P.; Tomaskova, N.; Kollarova, M.; Antalík, M. Malate dehydrogenases-structure and function. Gen. Physiol. Biophys. 2002, 21, 257–266. [Google Scholar]
- Pradedova, E.V.; Nimaeva, O.D.; Rakevich, A.L.; Salyaev, R.K. Comparative analyses of glutathione system of vacuoles and leucoplasts isolated from the storage parenchyma cells of dormant red beetroots (Beta vulgaris L.). Plant Physiol. Biochem. 2019, 145, 52–63. [Google Scholar] [CrossRef]
- Scheibe, R. Malate valves to balance cellular energy supply. Physiol Plant. 2004, 120, 21–26. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Bykova, N.V. Mitochondria in photosynthetic cells: Coordinating redox control and energy balance. Plant Physiol. 2023, 191, 2104–2119. [Google Scholar] [CrossRef]
- Goward, C.R.; Nicholls, D.J. Malate dehydrogenase: A model for structure, evolution, and catalysis. Protein Sci. 1994, 3, 1883–1888. [Google Scholar] [CrossRef]
- Møller, I.M.; Rasmusson, A.G.; Aken, O.V. Plant mitochondria–past, present and future. Plant J. 2021, 108, 912–959. [Google Scholar] [CrossRef]
- Ashton, A.R.; Trevanion, S.J.; Carr, P.D.; Verger, D.; Ollis, D.L. Structural basis for the light regulation of chloroplast NADP malate dehydrogenase. Physiol. Plant. 2000, 110, 314–321. [Google Scholar] [CrossRef]
- Zhao, J.F.; Du, Y.W.; Yu, A.L. Study on abiotic stress response of MDH gene in millet. Chin. J. Nucl. Agric. 2020, 34, 2152–2160. (In Chinese) [Google Scholar]
- Shameer, S.; Ratcliffe, R.G.; Sweetlove, L.J. Leaf energy balance requires mitochondrial respiration and export of chloroplast NADPH in the light. Plant Physiol. 2019, 180, 1947–1961. [Google Scholar] [CrossRef]
- Liszka, A.; Schimpf, R.; Cartuche Zaruma, K.I.; Buhr, A.; Seidel, T.; Walter, S.; Knuesting, J.; Dreyer, A.; Dietz, K.J.; Scheibe, R.; et al. Three cytosolic NAD-malate dehydrogenase isoforms of Arabidopsis thaliana: On the crossroad between energy fluxes and redox signaling. Biochem. J. 2020, 477, 3673–3693. [Google Scholar] [CrossRef]
- Lin, Y.W.; Chen, W.T.; Yang, Q.; Zhang, Y.J.; Ma, X.Q.; Li, M. Genome-wide characterization and gene expression analyses of malate dehydrogenase (MDH) genes in low-phosphorus stress tolerance of Chinese fir (Cunninghamia lanceolata). Int. J. Mol. Sci. 2023, 24, 4414. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Wang, Y.L.; Sun, X.M.; Yuan, J.; Zhao, Z.Q.; Gao, J.; Wen, X.R.; Tang, F.; Kang, M.; Abliz, B.; et al. Genome-wide identification of MDH family genes and their association with salt tolerance in rice. Plants 2022, 11, 1498. [Google Scholar] [CrossRef]
- Zhu, S.N.; Chen, Z.J.; Xie, B.X.; Guo, Q.; Chen, M.H.; Liang, C.Y.; Bai, Z.L.; Wang, X.R.; Wang, H.C.; Liao, H.; et al. A phosphate starvation responsive malate dehydrogenase, GmMDH12 mediates malate synthesis and nodule size in soybean (Glycine max). Environ. Exp. Bot. 2021, 189, 104560. [Google Scholar] [CrossRef]
- Imran, M.; Tang, K.; Liu, J.Y. Comparative genome-wide analysis of the malate dehydrogenase gene families in cotton. PLoS ONE 2016, 11, e0166341. [Google Scholar] [CrossRef]
- Song, J.L.; Zou, X.Y.; Liu, P.D.; Cardoso, J.A.; Schultze-Kraft, R.; Liu, G.D.; Luo, L.J.; Chen, Z.J. Differential expressions and enzymatic properties of malate dehydrogenases in response to nutrient and metal stresses in Stylosanthes guianensis. Plant Physiol. Biochem. 2022, 170, 325–337. [Google Scholar] [CrossRef]
- Ma, B.Q.; Yuan, Y.Y.; Gao, M.; Xing, L.B.; Li, C.Y.; Li, M.J.; Ma, F.W. Genome-wide identification, classification, molecular evolution and expression analysis of malate dehydrogenases in apple. Int. J. Mol. Sci. 2018, 19, 3312. [Google Scholar] [CrossRef]
- Chen, X.H.; Zhang, J.; Zhang, C.; Wang, S.J.; Yang, M.S. Genome-wide investigation of malate dehydrogenase gene family in poplar (Populus trichocarpa) and their expression analysis under salt stress. Acta Physiol. Plant. 2021, 43, 28. [Google Scholar] [CrossRef]
- Zhuang, W.J.; Varshney, R.K. Editorial: Peanut genomics and biotechnology in breeding applications. Front. Plant Sci. 2023, 14, 1226637. [Google Scholar] [CrossRef]
- Liao, B.S. Current situation and potential analysis of peanut production in China. Chin. J. Oil Crop. 2020, 42, 161–166. (In Chinese) [Google Scholar]
- Bonku, R.; Yu, J. Health aspects of peanuts as an outcome of its chemical composition. Food Sci. Hum. Well. 2020, 9, 21–30. [Google Scholar] [CrossRef]
- Li, M.R.; Guo, S.; Ho, C.T.; Bai, N.S. Review on chemical compositions and biological activities of peanut (Arachis hypogeae L.). J. Food Biochem. 2022, 46, e14119. [Google Scholar] [CrossRef]
- Syed, F.; Arif, S.; Ahmed, I.; Khalid, N. Groundnut (Peanut) (Arachis hypogaea). In Oilseeds: Health Attributes and Food Applications; Tanwar, B., Goyal, A., Eds.; Springer: Singapore, 2021; pp. 93–122. [Google Scholar] [CrossRef]
- Zou, Z.Y.; Chen, J.; Wu, W.J.; Luo, J.H.; Long, T.; Wu, Q.S.; Wang, Q.L.; Zhen, J.B.; Zhao, Y.P.; Wang, Y.C.; et al. Detection of peanut seed vigor based on hyperspectral imaging and chemometrics. Front. Plant Sci. 2023, 14, 1127108. [Google Scholar] [CrossRef]
- Sew, Y.S.; Ströher, E.; Fenske, R.; Millar, A.H. Loss of mitochondrial malate dehydrogenase activity alters seed metabolism impairing seed maturation and post-germination growth in Arabidopsis. Plant Physiol. 2016, 171, 849–863. [Google Scholar] [CrossRef]
- Yin, G.K.; Whelan, J.; Wu, S.H.; Zhou, J.; Chen, B.Y.; Chen, X.L.; Zhang, J.M.; He, J.J.; Xin, X.; Lu, X.X. Comprehensive mitochondrial metabolic shift during the critical node of seed ageing in rice. PLoS ONE 2016, 11, e0148013. [Google Scholar] [CrossRef]
- Xia, F.S.; Cheng, H.; Chen, L.L.; Zhu, H.S.; Mao, P.S.; Wang, M.Y. Influence of exogenous ascorbic acid and glutathione priming on mitochondrial structural and functional systems to alleviate aging damage in oat seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef]
- Li, P.X.; Tang, G.Y.; Xu, P.L.; Zhu, J.Q.; Shan, L.; Wan, S.B. Structure and expression analysis of mitochondrial malate dehydrogenase gene AhMMDH1 in peanut. Chin. J. Oil Crop. 2020, 42, 1090–1099. (In Chinese) [Google Scholar]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef]
- Skórka, M.; Sieprawska, A.; Telk, A. The implication of manganese surplus on plant cell homeostasis: A review. J. Plant Growth Regul. 2023, 42, 1327–1341. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Li, J.Y.; Pan, Y.H.; Chen, J.Y.; Liu, Y. Metabolic responses to manganese toxicity in soybean roots and leaves. Plants 2023, 12, 3615. [Google Scholar] [CrossRef]
- Li, J.F.; Jia, Y.D.; Dong, R.S.; Huang, R.; Liu, P.D.; Li, X.Y.; Wang, Z.Y.; Liu, G.D.; Chen, Z.J. Advances in the mechanisms of plant tolerance to manganese toxicity. Int. J. Mol. Sci. 2019, 20, 5096. [Google Scholar] [CrossRef] [PubMed]
- Le¡ková, A.; Giehl, R.F.H.; Hartmann, A.; Farga¡ová, A.; Wirén, N. Heavy metals induce iron deficiency responses at different hierarchic and regulatory levels. Plant Physiol. 2017, 174, 1648–1668. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, M.; Chen, J.Y.; Yang, S.X.; Chen, J.P.; Xue, Y.B. Comparative transcriptome analysis reveals complex physiological response and gene regulation in peanut roots and leaves under Mn toxicity stress. Int. J. Mol. Sci. 2023, 24, 1161. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Sheng, X.G.; Zhao, Z.Q.; Wang, J.S.; Yu, H.F.; Shen, Y.S.; Zeng, X.Y.; Gu, H.H. Genome wide analysis of MADS-box gene family in Brassica oleracea reveals conservation and variation in flower development. BMC Plant Biol. 2019, 19, 106. [Google Scholar] [CrossRef]
- Shen, G.; Jia, Y.; Wang, W.L. Evolutionary divergence of motifs in B-class MADS-box proteins of seed plants. J. Biol. Res. 2021, 28, 12. [Google Scholar] [CrossRef]
- Zheng, Y.C.; Chen, X.J.; Wang, P.J.; Sun, Y.; Yue, C.; Ye, N.X. Genome-wide and expression pattern analysis of JAZ family involved in stress responses and postharvest processing treatments in Camellia sinensis. Sci. Rep. 2020, 10, 2792. [Google Scholar] [CrossRef]
- Shao, Z.W.; He, M.H.; Zeng, Z.P.; Chen, Y.Z.; Hanna, A.; Zhu, H.B. Genome-wide identification and expression analysis of the MADS-Box gene family in sweet potato [Ipomoea batatas (L.) Lam]. Front. Genet. 2021, 12, 750137. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, Y.B.; Xie, B.X.; Zhu, S.N.; Lu, X.; Liang, C.Y.; Tian, J. Complex gene regulation between young and old soybean leaves in responses to manganese toxicity. Plant Physiol. Biochem. 2020, 155, 231–242. [Google Scholar] [CrossRef]
- Zhang, L.H.; Wang, C.Z.; Jia, R.P.; Yang, N.X.; Jin, L.; Zhu, L.C.; Ma, B.Q.; Yao, Y.X.; Ma, F.W.; Li, M.J. Malate metabolism mediated by the cytoplasmic malate dehydrogenase gene MdcyMDH affects sucrose synthesis in apple fruit. Hort. Res. 2022, 9, uhac194. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, R.; Yang, M.; Law, Y.S.; Sun, F.; Hon, N.L.; Ngai, S.M.; Lim, B.L. A balance between the activities of chloroplasts and mitochondria is crucial for optimal plant growth. Antioxidants 2021, 10, 935. [Google Scholar] [CrossRef]
- Wang, Z.A.; Li, Q.; Ge, X.Y.; Yang, C.L.; Luo, X.L.; Zhang, A.H.; Xiao, J.L.; Tian, Y.C.; Xia, G.X.; Chen, X.Y.; et al. The mitochondrial malate dehydrogenase 1 gene GhmMDH1 is involved in plant and root growth under phosphorus deficiency conditions in cotton. Sci. Rep. 2015, 5, 10343. [Google Scholar] [CrossRef]
- Yin, X.; Yuan, Y.; Han, X.; Han, S.; Li, Y.; Ma, D.; Fang, Z.; Gong, S.; Yin, J. Genome-wide identification, characterization, and expression profiling of TaDUF668 gene family in Triticum aestivum. Agronomy 2023, 13, 2178. [Google Scholar] [CrossRef]
- Beeler, S.; Liu, H.C.; Stadler, M.; Schreier, M.; Eicke, S.; Lue, W.L.; Truernit, E.; Zeeman, S.; Chen, J.; Kötting, O. Plastidial NAD-dependent malate dehydrogenase is critical for embryo development and heterotrophic metabolism in Arabidopsis. Plant Physiol. 2014, 164, 1175–1190. [Google Scholar] [CrossRef]
- Menckhoff, L.; Mielke-Ehret, N.; Buck, F.; Vuletić, M.; Lüthje, S. Plasma membrane-associated malate dehydrogenase of maize (Zea mays L.) roots: Native versus recombinant protein. J. Proteom. 2013, 80, 66–77. [Google Scholar] [CrossRef]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.; Ströher, E.; Smith, S.; Gardeström, P.; Millar, A.H. Mitochondrial malate dehydrogenase lowers leaf respiration and alters photorespiration and plant growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef]
- Roussi, Z.; Ben Mrid, R.; Ennoury, A.; Nhhala, N.; Zouaoui, Z.; Omari, R.E.; Nhiri, M. Insight into Cistus salviifolius extract for potential biostimulant effects in modulating cadmium-induced stress in sorghum plant. Physiol. Mol. Biol. Plants 2022, 28, 1323–1334. [Google Scholar] [CrossRef]
- Crombie, A.T. The effect of lanthanum on growth and gene expression in a facultative methanotroph. Environ. Microbiol. 2022, 24, 596–613. [Google Scholar] [CrossRef]
- Chen, Z.J.; Sun, L.L.; Liu, P.D.; Liu, G.D.; Tian, J.; Liao, H. Malate synthesis and secretion mediated by a manganese-enhanced malate dehydrogenase confers superior manganese tolerance in Stylosanthes guianensis. Plant Physiol. 2015, 167, 176–188. [Google Scholar] [CrossRef]
- Peng, W.M.H. Functional Study of Rice Malate Dehydrogenase Gene OsMDH; Yangzhou University: Yangzhou, China, 2022. (In Chinese) [Google Scholar]
- Wu, X.; Han, Y.Y.; Zhu, X.Y.; Shah, A.; Wang, W.; Sheng, Y.B.; Fan, T.T.; Cao, A.Q. Negative regulation of cadmium tolerance in Arabidopsis by MMDH2. Plant Mol. Biol. 2019, 101, 507–516. [Google Scholar] [CrossRef]
- Zhu, X.Y. Functional Analysis of mMDH2 Gene in Regulating Iron Deficiency Response of Arabidopsis Thaliana; Hefei University of Technology: Hefei, China, 2021. (In Chinese) [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).