


DNA Methylation as a Biomarker for Monitoring Disease Outcome in Patients with Hypovitaminosis and Neurological Disorders




Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Collection and Analysis
2.3. Neuropsychological Assessments
2.4. DNA Extraction
2.5. Quantification of Global DNA Methylation (5mC)
2.6. Statistical Analysis
3. Results
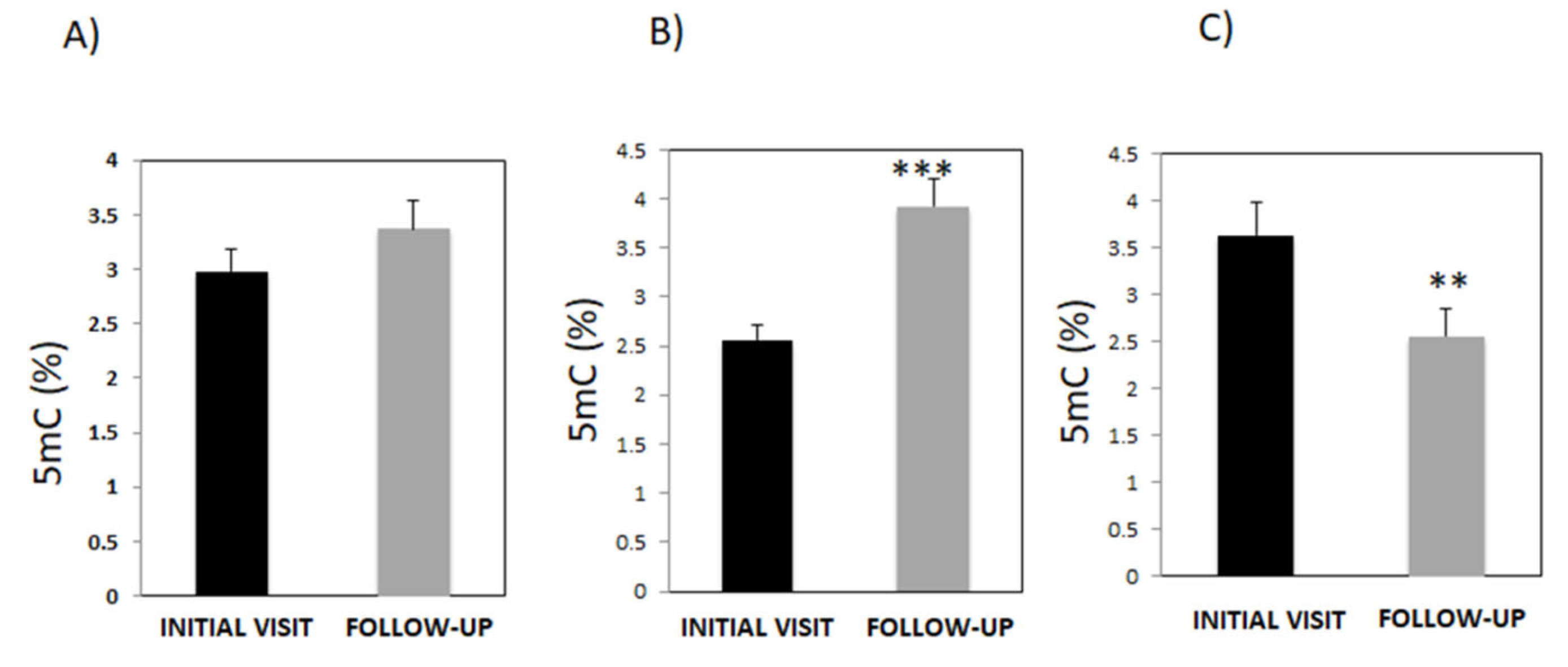
3.1. Evaluation of Global DNA Methylation after the Initial Consult and after Treatment
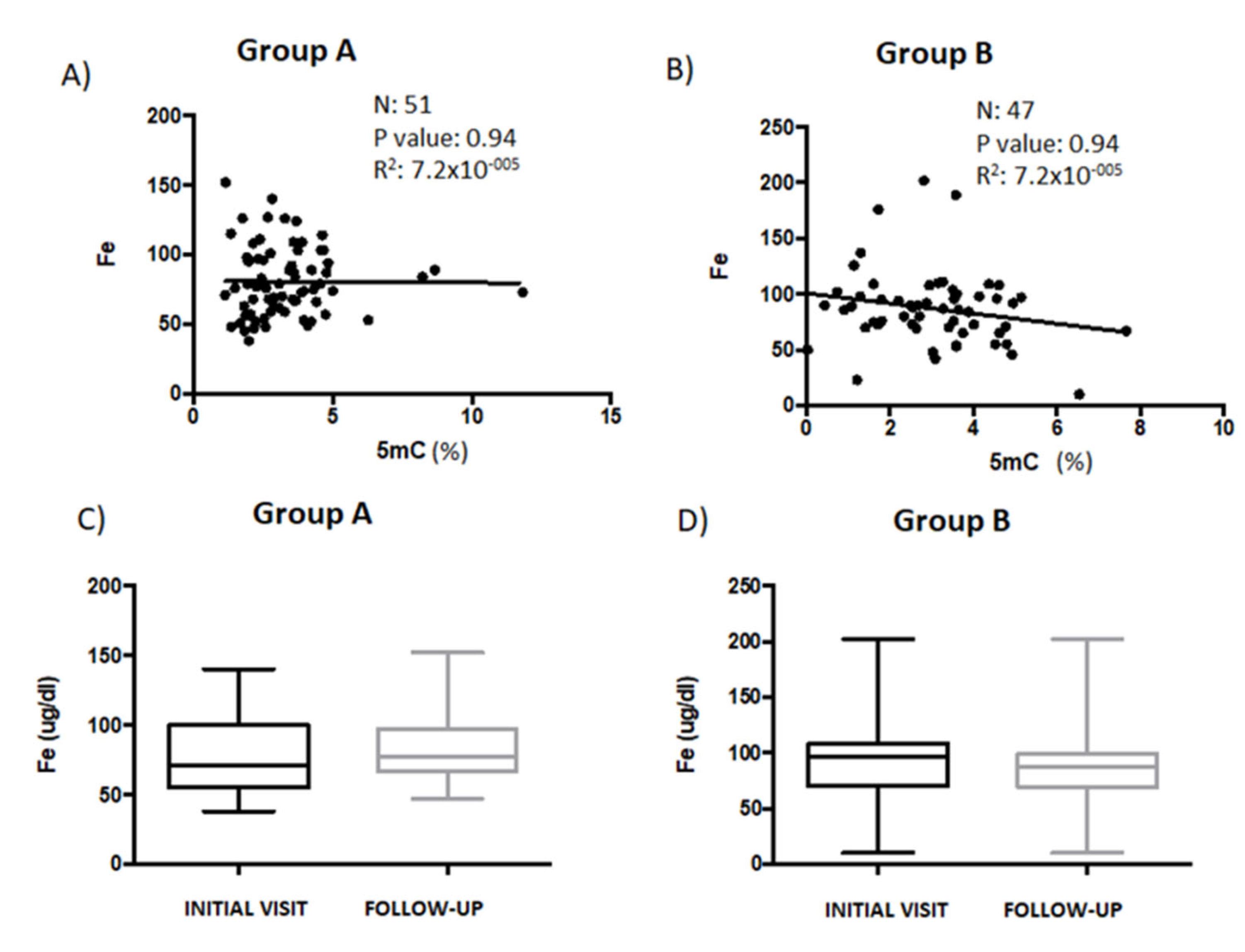
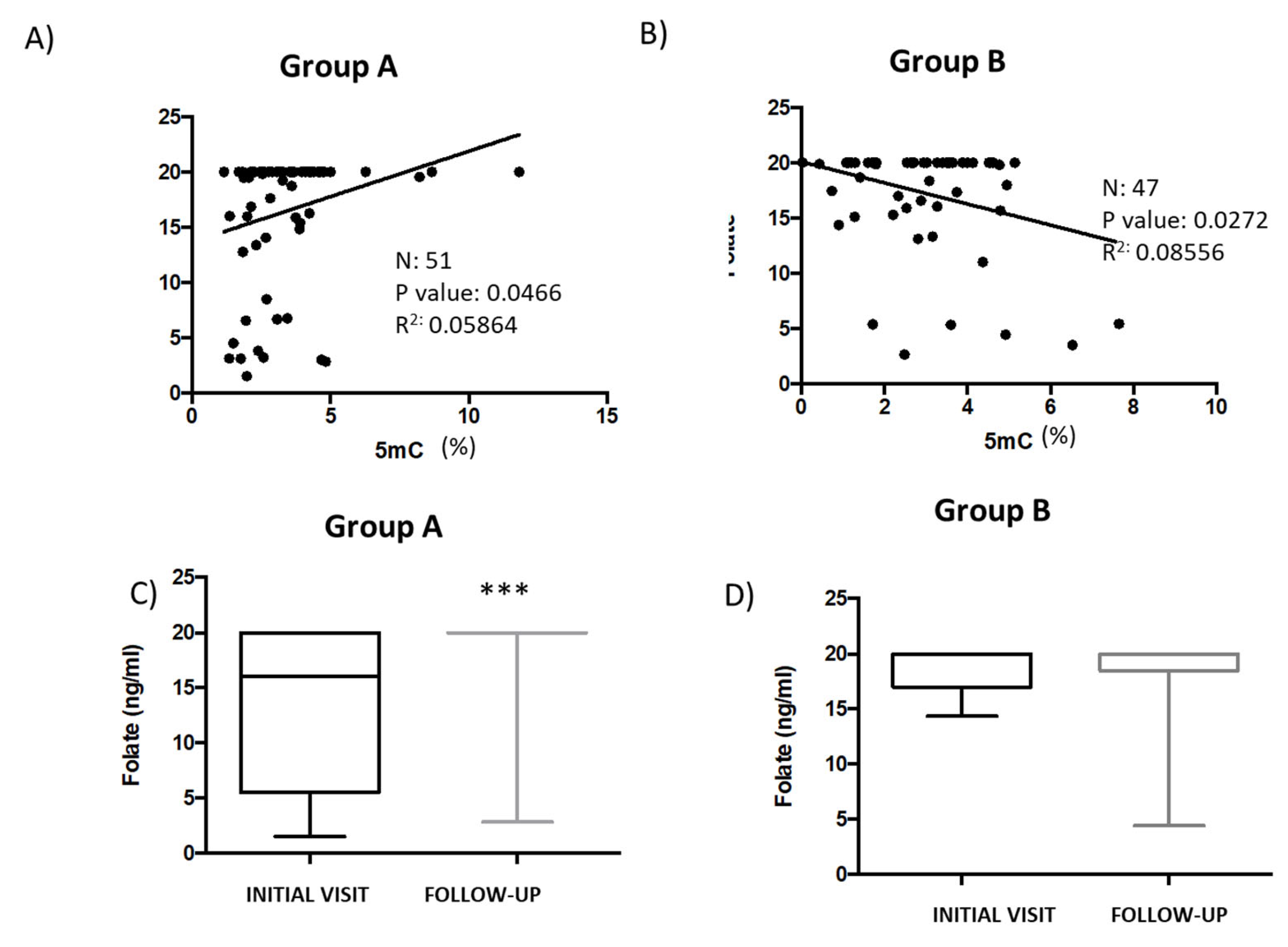
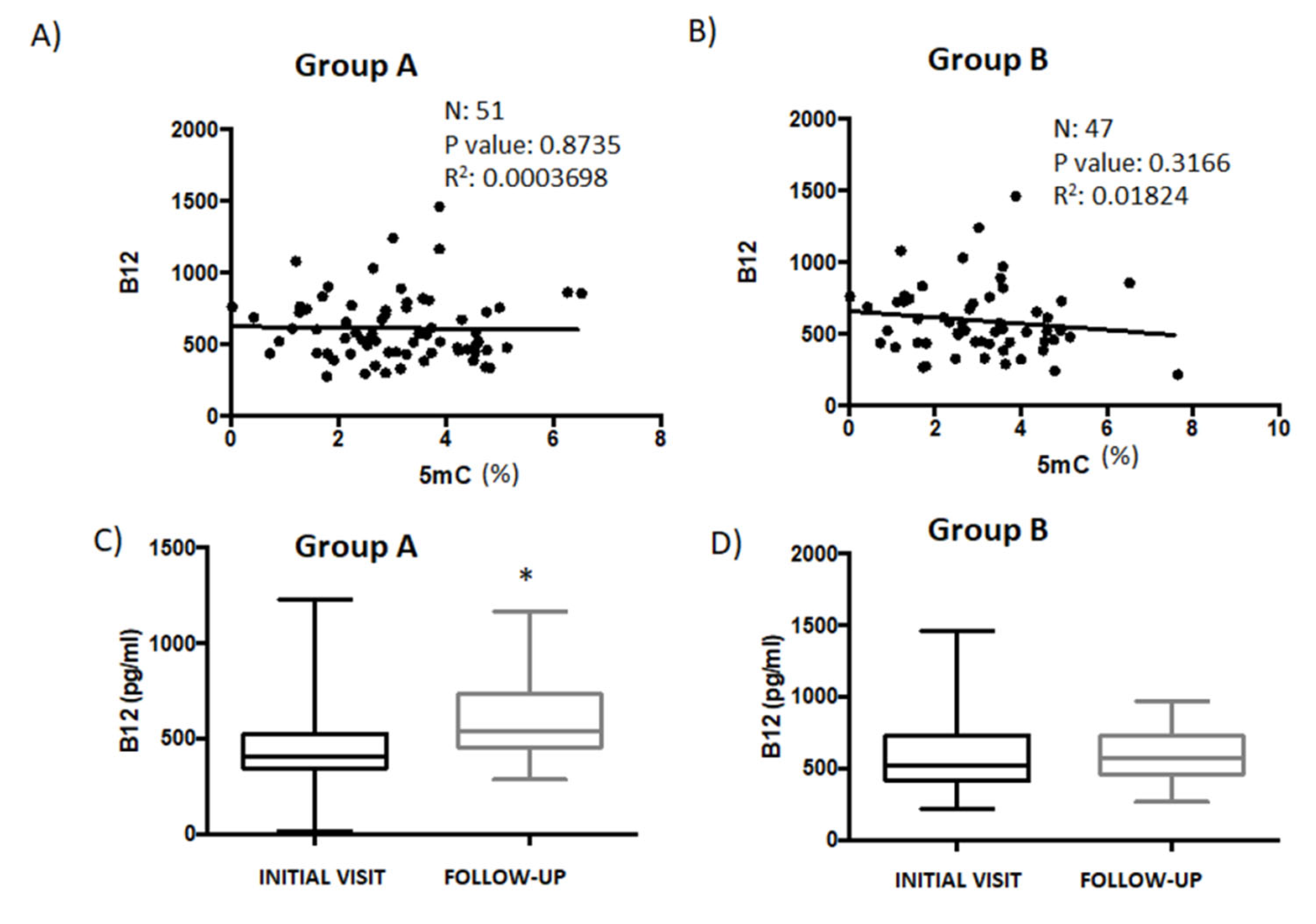
3.2. Correlation Analysis between Global DNA Methylation and Iron, Folate, and Vitamin B12
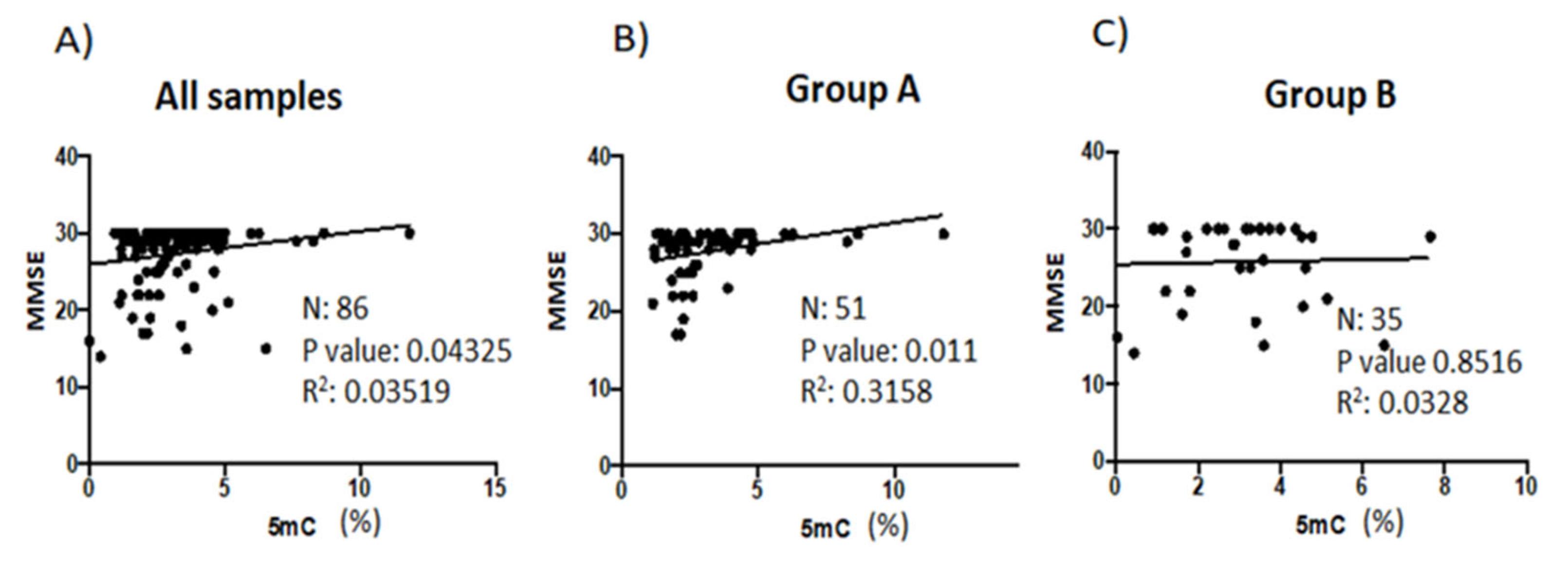
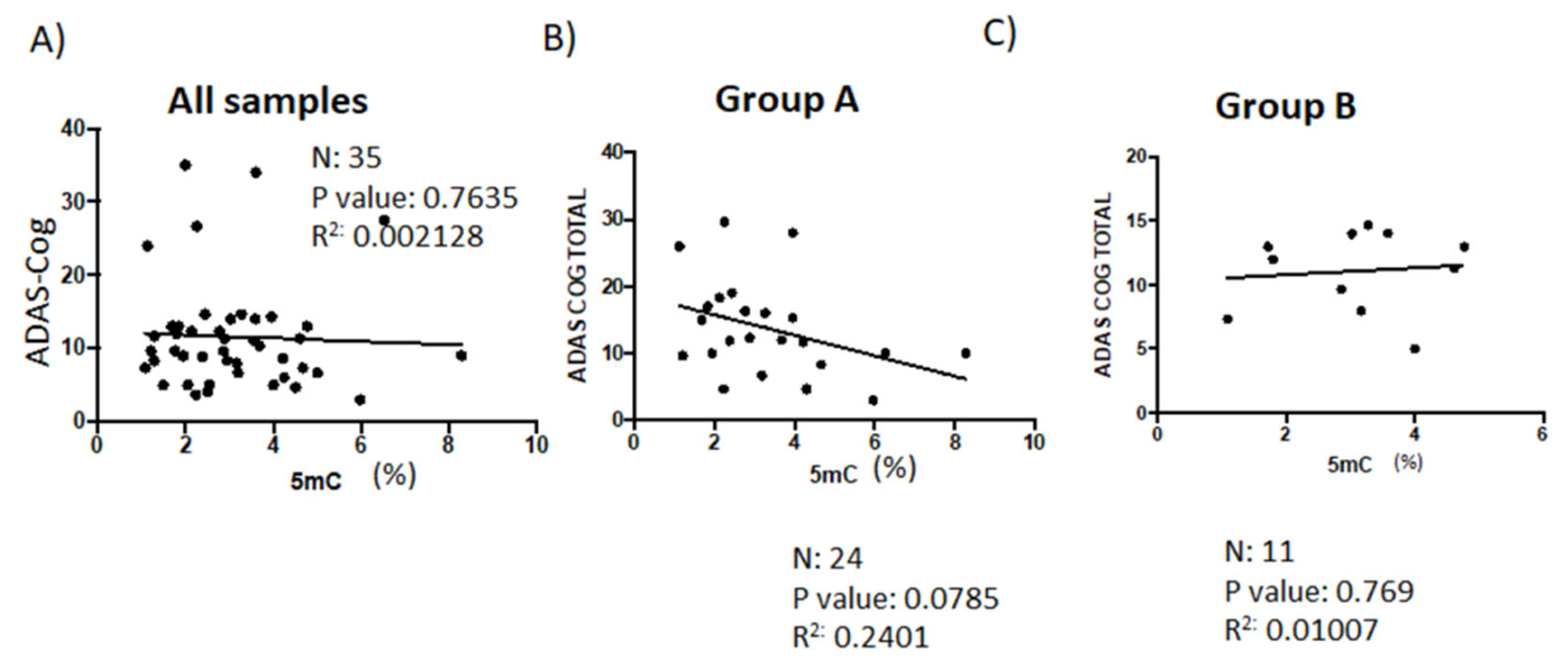
3.3. Correlation Analysis between Global DNA Methylation and Psychometric Assessments
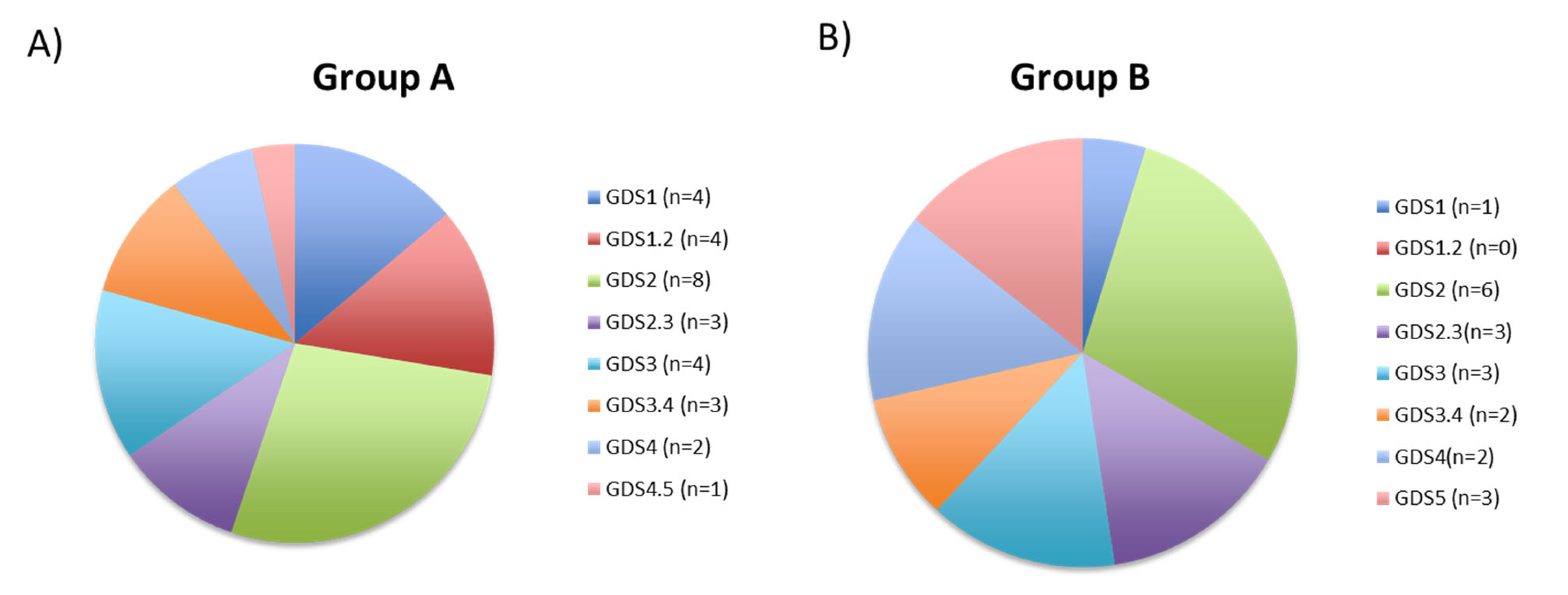
3.4. Correlation Analysis between Global DNA Methylation and Clinical Diagnosis
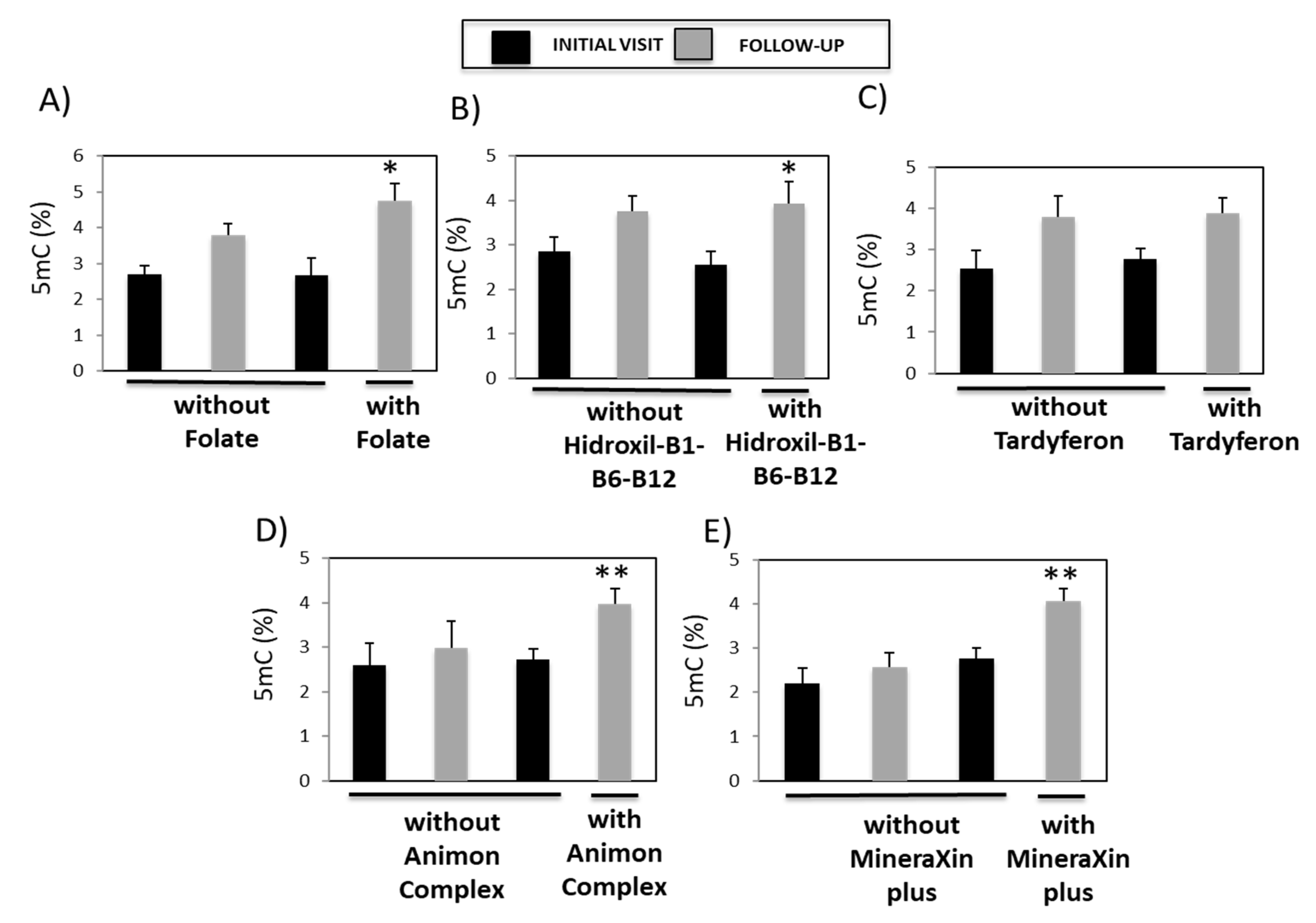
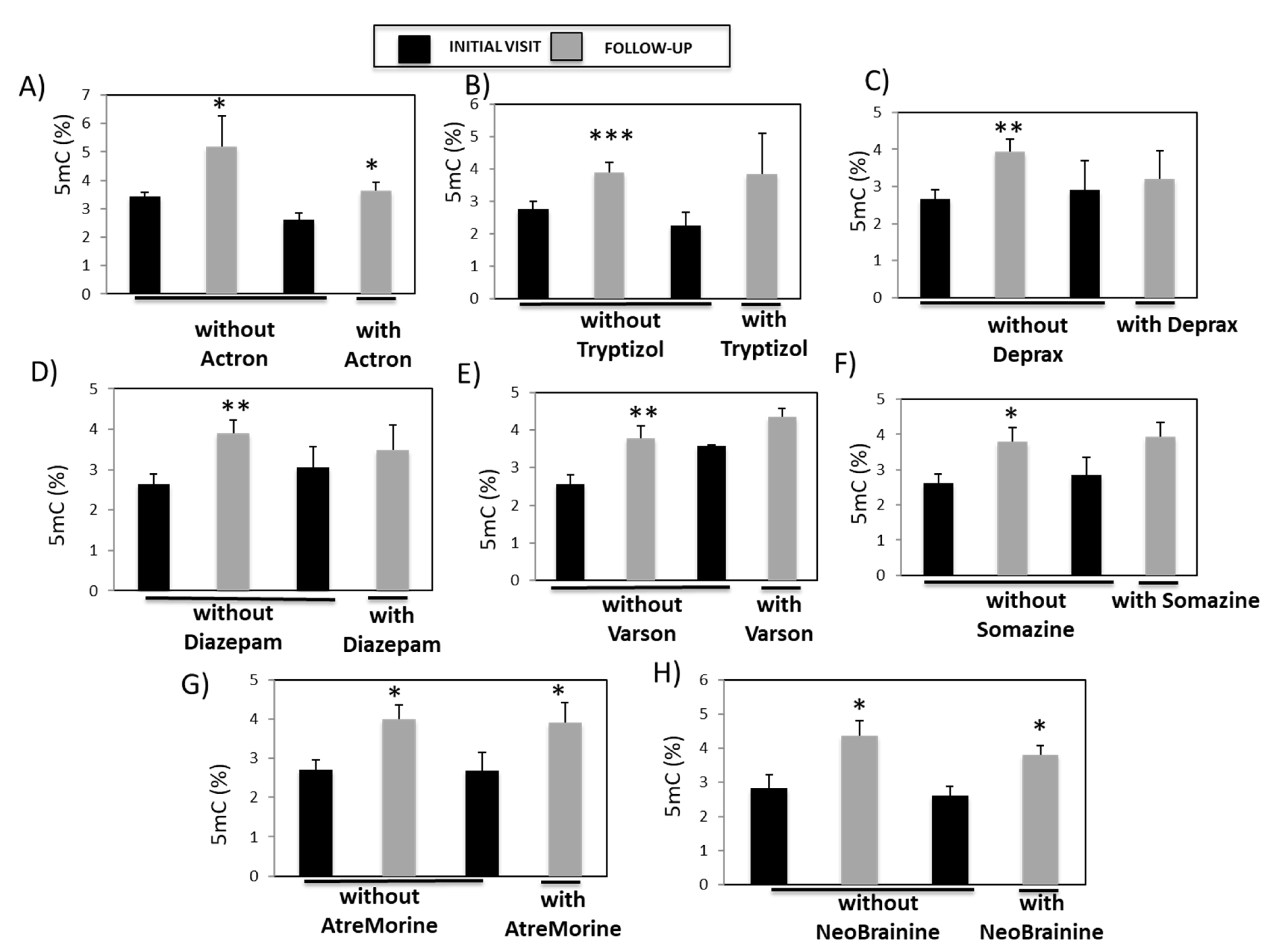
3.5. Analysis of the Association between Global DNA Methylation and Pharmacotherapy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Urdinguio, R.; Sanchez-Mut, J.; Esteller, M. Epigenetic mechanisms in neurological diseases: Genes, syndromes, and therapies. Lancet Neurol. 2009, 8, 1056–1072. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Tellado, I.; Cacabelos, P. The Epigenetic Machinery in the Life Cycle and Pharmacoepigenetics; Elsevier: Amsterdam, The Netherlands, 2019; Volume 10, pp. 1–100. [Google Scholar]
- Delgado-Morales, R.; Esteller, M. Opening up the DNA methylome of dementia. Mol. Psychiatry 2017, 22, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. The essentials of DNA methylation. Cell 1992, 70, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Gräff, J.; Mansuy, I. Epigenetic codes in cognition and behaviour. Behav. Brain Res. 2008, 1, 70–87. [Google Scholar] [CrossRef]
- Nan, X.; Campoy, F.; Bird, A. MeCP2 is a transcriptional repressor with abundant binding sites in genomic chromatin. Cell 1997, 21, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Fan, G. The role of DNA methylation in the central nervous system and Neiropsychiatric Disorders. Int. Rev. Neurobiol. 2009, 89, 67–84. [Google Scholar]
- Al-Mahdawi, S.; Virmouni, S.A.; Pook, M.A. DNA Methylation in Neurodegenerative Diseases. Epigenetic Biomark. Diagn. 2016, Chaper 12, 401–415. [Google Scholar]
- Sung, H.Y.; Choi, E.N.; Jo, S.A.; Oh, S.; Ahn, J.H. Amyloid protein-mediated differential DNA methylation status regulates genes expression in Alzheimer’s disease model cell line. Biochem. Biophys. Res. Commun. 2011, 414, 700–705. [Google Scholar] [CrossRef]
- Martínez-Iglesias, O.; Carrera, I.; Carril, J.C.; Férnandez-Novoa, L.; Cacabelos, N.; Cacabelos, R. DNA methylation in neurodegenerative and cerebrovascular disorders. Int. J. Mol. Sci. 2020, 21, 2220. [Google Scholar] [CrossRef] [Green Version]
- Sales, A.J.; Joca, S. Effects od DNA methylation inhibitors and conventional antideoressants on mice bahaviour and brain DNA methylation levels. Acta Neuroph. 2015, 28, 11–22. [Google Scholar]
- Mastroeni, D.; McKee, A.; Grover, A.; Togers, J.; Coleman, P. Epigenetic differneces in cortical neurons from a pair of monozygotic twins discordant for Alzheimer’s disease. PLoS ONE 2009, 4, e6617. [Google Scholar] [CrossRef] [Green Version]
- Chouliaras, L.; Mastroeni, D.; Delvaux, E.; Grover, A.; Kenis, G.; Hof, P.R.; Steinbusch, H.W.; Coleman, P.D.; Rutten, B.P.; van den Hove, D.L. Consistent decrease in global DNA methylation and hydroxymethylation in the hippocampus of Alzheimer’s disease patients. Neurobiol. Aging 2013, 34, 2091–2099. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Iglesias, O.; Naidoo, V.; Cacabelos, N.; Cacabelos, R. Epigenetic Biomarkers as diagnostic tools for Neurodegenerative Disorders. Int. J. Mol. Sci. 2022, 23, 13. [Google Scholar] [CrossRef]
- Masliah, E.; Dumaop, W.; Galasko, D.; Desplats, P. Distinctive patterns of DNA methylation associated with Parkinson disease: Identification of concordant epigenetic changes in brain and peripheral blood leukocytes. Epigenetics 2013, 1030–1038. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Arosio, B.; Falconi, A.; Di Bonaventura, M.V.M.; Karimi, M.; Mari, D.; Casati, M.; Maccarrone, M.; D’Addario, C. Global changes in DNA methylation in Alzheimer’s disease peripheral blood mononuclear cells. Brain Behav. Immun. 2015, 45, 139–144. [Google Scholar] [CrossRef]
- Salemeh, Y.; Bejaoui, Y.; El Hajj, N. DNA methylation biomarkers in aging and age-related diseases. Front. Genet. 2020, 11, 171. [Google Scholar] [CrossRef]
- Lu, H.; Liu, X.; Deng, Y.; Qing, H. DNA methylation, a hand behind neurodegenerative diseases. Front. Aging Neurosci. 2013, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Bergsma, T.; Rogaeva, E. DNA methylation Clocks and their predictive capacity for aging phenotypes and healthspan. Neurosci. Insights 2020, 15. [Google Scholar] [CrossRef]
- Robertson, K.D. DNA methylation and human disease. Nat, Rev, Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef]
- Jun, Z.; Liu, Y. DNA methylation in human diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef]
- Kadmiri, N.E.; Said, N.; Slassi, I.; Moutawakil, B.E.; Nadifi, S. Biomarkers for Alzheimer Disease: Classical and Novel Candidates’ Review. Neuroscience 2018, 1, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Burtis, C.; Ashwood, E. Tietz Textbook of Clinical Chemistry, 3rd ed; AACC: Washington, DC, USA, 1999. [Google Scholar]
- Tuck, M.K.; Chan, D.W.; Chia, D.; Godwin, A.K.; Grizzle, W.E.; Krueger, K.E.; Rom, W.; Sanda, M.; Sorbara, L.; Stass, S.; et al. Standard operating Procedures for Serum and Plasma Collection: Early Detection Research Network Consensus Statement Standard Operating Procedure Integration Working Group. J. Proteome Res. 2009, 8, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, A.H.B. Tietz Clinical Guide to Laboratory Tests; Elsevier: Amsterdam, The Netherlands, 2006; pp. 916–917. [Google Scholar]
- Folstein, M.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State” a Practical Method for Grading the Cognitive State of Patients for the Clinician. J. Psychiatric Res. 1975, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Mohs, E.C.; Rosen, W.G.; Davis, G.L. The Alzheimer’s Disease Assessment Scale: An instrument for assessing treatment efficacy. Psychopharmacol Bull 1983, 19, 448–450. [Google Scholar]
- Rosen, W.G.; Mohs, R.C.; Davis, K.L. A new rating scale for Alzheimer disease. Am. J. Psychiatry 1984, 14, 1356–1364. [Google Scholar]
- Reisberg, B. Functional assessment staging (FAST). Psychopharmacol Bull 1988, 24, 653–659. [Google Scholar]
- Franquet, P.D.; Lacazare, P.; Saffery, R.; Mcneil, J.; Woods, R.; Ryan, J. Blood DNA methylation as a potential biomarker of Dementia: A systematic review. Alzheimer’s Dement. 2017, 14, 81–103. [Google Scholar] [CrossRef]
- Deng, Q.; Huang, W.; Peng, C.; Gao, J.; Li, Z.; Qiu, X.; Yang, N.; Yuan, B.; Zheng, F. Genomic 5mC contents in peripheral blood leukocytes were in peripheral blood leukocytes were independent protective factors for coronary artery disease with a specific profile in different leukocyte subtypes. Clin. Epigenetics 2018, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Long, T.; Arakawa, K.; Wang, R.; Yu, M.; Laird, P. DNA methylation as a biomarker for cardiovascular disease risk. PLoS ONE 2010, 5, e9692. [Google Scholar] [CrossRef] [Green Version]
- Wo, H.D.; Kim, J. Global DNA hypomethylation in peripheral blood leukocytes as a biomarker for cancer risk: A meta analysis. PLoS ONE 2012, 7, e34615. [Google Scholar] [CrossRef]
- Lien, Y.-C.; Condon, D.E.; Georgieff, M.K.; Simmons, R.A.; Tran, P. Dysregulation of neuronal genes by fetal-neonatal iron deficiency anemia is associated with altered DNA methylation in the rat hippocampus. Nutrients 2019, 11, 1191. [Google Scholar] [CrossRef] [Green Version]
- Irwin, R.E.; Pentieva, K.; Cassidy, T.; Lees-Murdock, D.; McLaughlin, M.; Prasad, G.; McNulty, H.; Walsh, C. The interplay between DNA methylation, folate and neuocognitive development. Epigenomics 2016, 8, 863–879. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, A.; Sapehia, D.; Thakur, S.; Mohanraj, P.; Bagga, R.; Kaur, J. Effect of imbalance in folate and vitamin B12 in maternal/parental diet on global methylation and regultaory miRNAs. Sci. Rep. 2019, 9, 17602. [Google Scholar] [CrossRef] [Green Version]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA methylation: A review of molecular mechanisms and the evidence for folate’s role. Adv. Nutr. 2012, 31, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Cano, S.; Posner, H.; Moline, M.; Hurt, S.; Swartz, J.; Hsu, T.; Hobart, J.C. The ADAS-cog in Alzheimer’s disease clinical trials:psychometric evaluation of the sum and its parts. J. Neurol. Neurosurg. Psych. 2010, 8, 1363–1368. [Google Scholar] [CrossRef] [Green Version]
- Cacabelos, R.; Carril, J.C.; Cacabelos, N.; Kazantsev, A.G.; Vostrov, A.V.; Corzo, L.; Cacabelos, P.; Goldgaber, D. Sirtuins in Alzheimer’s Disease: SIRT2-Related GenoPhenotypes and Implications for PharmacoEpiGenetics. Int. J. Mol. Sci. 2019, 20, 1249. [Google Scholar] [CrossRef] [Green Version]
- Corzo, L.; Fernandez-Novoa, L.; Carrera, I.; Martinez, O.; Rodriguez, S.; Alejo, R.; Cacabelos, R. Nutrition, health and disease: Role of selected marine and vegetal. Nutrients 2020, 12, 747. [Google Scholar] [CrossRef] [Green Version]
- Cacabelos, R.; Alvarez, X.A.; Franco-Maside, A.; Fernández-Novoa, L.; Caamaño, J. Effect of CDP-choline on cognition and immune function in Alzheimer’s disease and multi-infarct dementia. Ann. N. Y. Acad. Sci. 1993, 695, 321–323. [Google Scholar] [CrossRef]
- Romero, A.; Parada, E.; González-Lafuente, L.; Farré-Alins, V.; Ramos, E.; Cacabelos, R.; Egea, J. Neuroprotective effects of E-PodoFavalin-15999 (Atremorine). CNS Neurosci. Ther. 2017, 23, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Carrera, I.; Fernández-Novoa, L.; Sampedro, C.; Tarasov, V.V.; Aliev, G.; Cacabelos, R. Dopaminergic neuroprotection with Atremorine in parkisnon’s Disease. Curr. Med. Chem. 2018, 25, 5372–5388. [Google Scholar] [CrossRef]
- Kaur, G.; Rathod, S.S.; Ghoneim, M.; Alshehri, S.; Ahmad, J.; Mishra, A.; Alhakamy, N. DNA methylation: A promising approach in the management of Alzheimer’s Disease and other Neurodegenerative Disorders. Biology 2022, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Allen, L. Causes of vitamin B12 and folate deficiency. Food Nutr. Bull. 2008, 29, S20–S37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, M.P.; Shea, T.B. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003, 26, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.A.; Sweatt, J.D. Covalent modification of DNA regulates memory formation. Neuron 2007, 53, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fioravanti, M.; Ferrario, E.; Massaia, M.; Cappa, G.; Rivolta, G.; Grossi, E.; Buckley, E. Low folate levels in the cognitive decline of elderly patients and the efficacy of folate as a treatment for improving memory deficits. Arch. Gerontol. Geriatr. 1998, 26, 1–13. [Google Scholar] [CrossRef]
- Fuso, A.; Seminara, L.; Cavallaro, R.; D’Anselmi, F.; Scarpa, S. S-adenosylmethionine/homocysteine cycle alterations modify DNA methylation status with consequent deregulation of PS1 and BACE and beta-amyloid production. Mol. Cell. Neurosci. 2005, 28, 195–204. [Google Scholar] [CrossRef]
- Chen, H.; Liu, S.; Ge, B.; Zhou, D.; Li, M.; Li, W.; Ma, F.; Liu, Z.; Ji, Y. Effects of folic acid and vitamin B12 supplementation on cognitive impairment and inflammation in patients with Alzheimer’s Disease: A randomized, single-blinded, placebo-controlled trial. J. Prevent. Alzheimer’s Dis. 2021, 8, 249–256. [Google Scholar] [CrossRef]
- Amenyah, S.; Hughes, C.; Ward, M.; Rosborough, S.; Deane, J.; Thursby, S.-J.; Walsh, C.; Kok, D.; Strain, J.J.; McNulty, H.; et al. Influence of nutrients involved in one-carbon metabolism on DNA methylation in adults- a systematic review and meta-analysis. Nutr. Rev. 2020, 78, 647–666. [Google Scholar] [CrossRef]
- Fernández-Roig, S.; Lai, S.-C.; Murphy, M.; Fernandez-Ballart, J.; Quadros, E. Vitamin B12 deficiency in the brain leads to DNA hypomethylation in the TCblR/CD320 knockout mouse. Nutr. Metab. 2012, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Boughanem, H.; Hernandez-Alonso, P.; Tinahones, A.; Babio, N.; Salas-salvado, J.; Tinahones, F.; Macias Gonzalez, M. Association between serum vitamin B12 and global DNA methylation in colorectal cancer patients. Nutrients 2020, 12, 3567. [Google Scholar] [CrossRef]
- Chuang, Y.H.; Lu, A.T.; Paul, K.C.; Folle, A.D.; Bronstein, J.M.; Bordelon, Y.; Horvath, S.; Ritz, B. Longitudinal epigenome-wide methylation study of cognitive decline and motor progression in Parkisnon’s Disese. J. Park. Dis. 2019, 9, 389–400. [Google Scholar]
- Bollati, V.; Galimberti, D.; Pergoli, L.; Dalla Valle, E.; Barretta, F.; Cortini, F.; Scarpini, E.; Bertazzi, P.A.; Baccarelli, A. DNA methylation in repetitive elements and Alzheimer disease. Brain Behav. Immun. 2011, 25, 1078–1083. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Iglesias, O.; Naidoo, V.; Carril, J.C.; Carrera, I.; Corzo, L.; Rodríguez, S.; Alejo, R.; Cacabelos, N.; Cacabelos, R. AtreMorine treatment regulates DNA methylation in neurodegenerative disorders: Epigenetic and pharmacogenetic studies. Curr. Pharm. Pers. Med. 2020, 17, 159–171. [Google Scholar] [CrossRef]
- Bekdash, R.A. Choline and the Brain: An epigenetic perspective. Adv. Neurobiol. 2016, 12, 381–399. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GROUP A | GROUP B | ||
|---|---|---|---|
| Clinical diagnosis | % | Clinical diagnosis | % |
| Hypovitaminosis | 80 | Hypovitaminosis | 37.5 |
| Neurocognitive deficit | 13.79 | Neurocognitive deficit | 18.75 |
| Parkinson’s disease | 20.69 | Parkinson’s disease | 6.25 |
| Mixed dementia | 13.79 | Mixed dementia | 18.75 |
| Vascular headache | 6.90 | Cerebrovascular disorder | 18.75 |
| Cerebrovascular disorder | 24.14 | Anxiety/Depression | 26.67 |
| Anxiety/Depression | 27.59 | Others (Bipolar disorder, Epilepsy, Psychotic disorder) | 25 |
| Others (Bipolar disorder, Epilepsy, Psychotic disorder) | 17.24 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Iglesias, O.; Naidoo, V.; Corzo, L.; Pego, R.; Seoane, S.; Rodríguez, S.; Alcaraz, M.; Muñiz, A.; Cacabelos, N.; Cacabelos, R. DNA Methylation as a Biomarker for Monitoring Disease Outcome in Patients with Hypovitaminosis and Neurological Disorders. Genes 2023, 14, 365. https://doi.org/10.3390/genes14020365
Martínez-Iglesias O, Naidoo V, Corzo L, Pego R, Seoane S, Rodríguez S, Alcaraz M, Muñiz A, Cacabelos N, Cacabelos R. DNA Methylation as a Biomarker for Monitoring Disease Outcome in Patients with Hypovitaminosis and Neurological Disorders. Genes. 2023; 14(2):365. https://doi.org/10.3390/genes14020365
Chicago/Turabian StyleMartínez-Iglesias, Olaia, Vinogran Naidoo, Lola Corzo, Rocío Pego, Silvia Seoane, Susana Rodríguez, Margarita Alcaraz, Adriana Muñiz, Natalia Cacabelos, and Ramón Cacabelos. 2023. "DNA Methylation as a Biomarker for Monitoring Disease Outcome in Patients with Hypovitaminosis and Neurological Disorders" Genes 14, no. 2: 365. https://doi.org/10.3390/genes14020365
APA StyleMartínez-Iglesias, O., Naidoo, V., Corzo, L., Pego, R., Seoane, S., Rodríguez, S., Alcaraz, M., Muñiz, A., Cacabelos, N., & Cacabelos, R. (2023). DNA Methylation as a Biomarker for Monitoring Disease Outcome in Patients with Hypovitaminosis and Neurological Disorders. Genes, 14(2), 365. https://doi.org/10.3390/genes14020365