
Temporal and Spatial Gene Expression Profile of Stroke Recovery Genes in Mice
 and
and 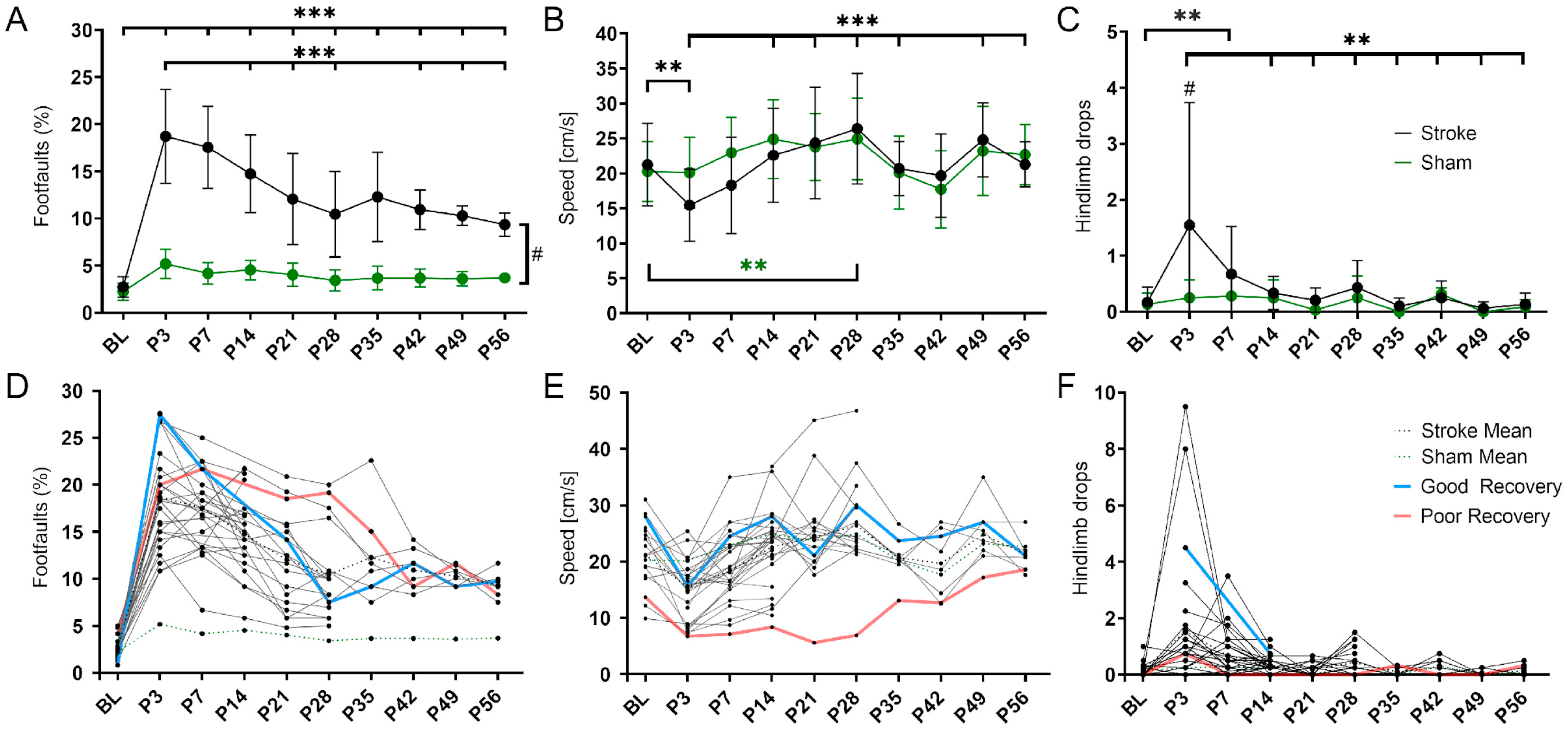{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Study Design and Animals
2.2. Photothrombosis
2.3. Behavioral Testing and Scoring
2.4. MRI Data Acquisition and Processing
2.5. Sample Preparation and qPCR Protocol
2.6. Data Analysis and Statistics
2.7. Recovery Rate Analysis
2.8. qPCR Analysis
3. Results
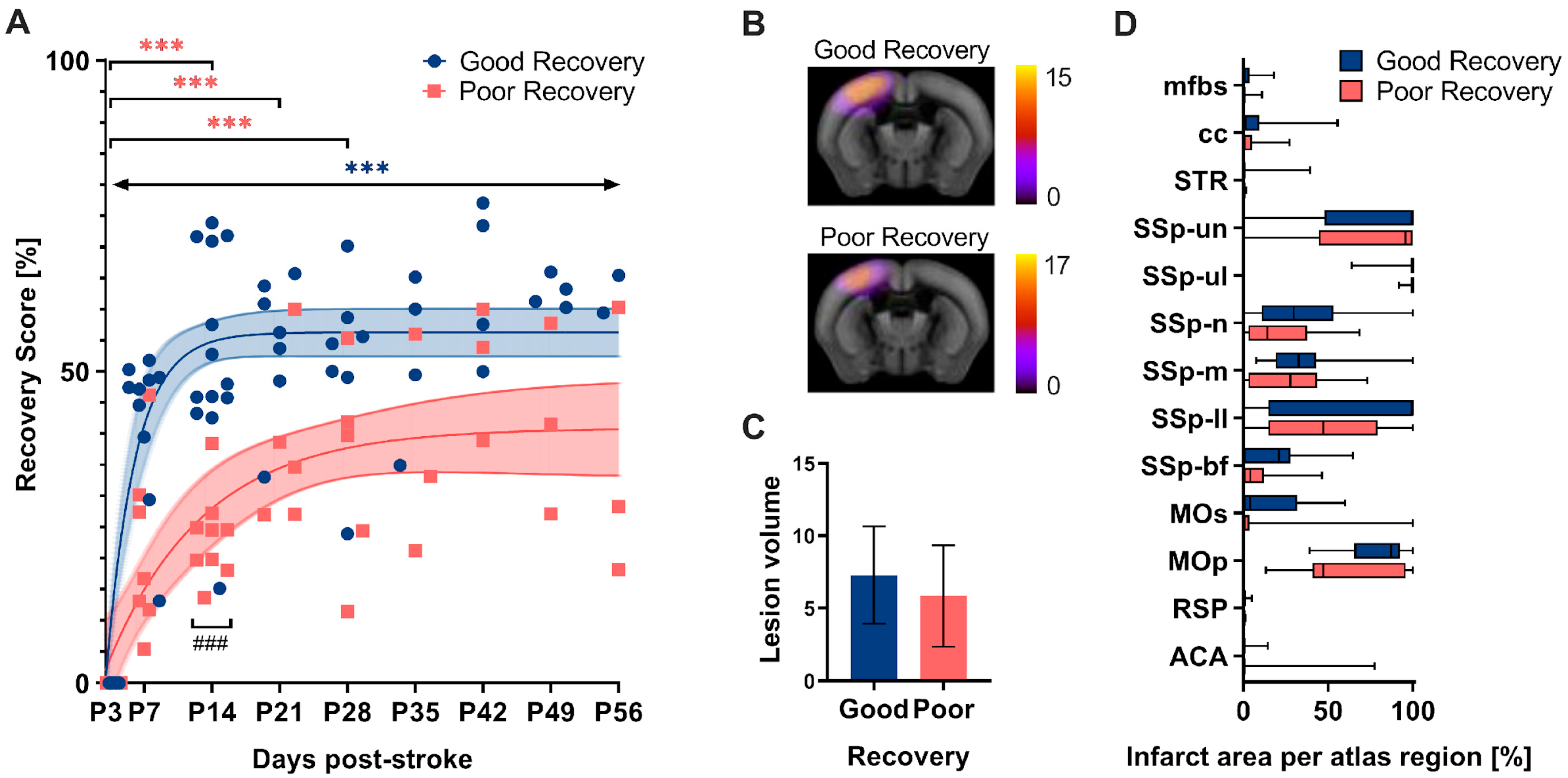
3.1. Spontaneous Functional Recovery
3.2. Gene Expression Analysis
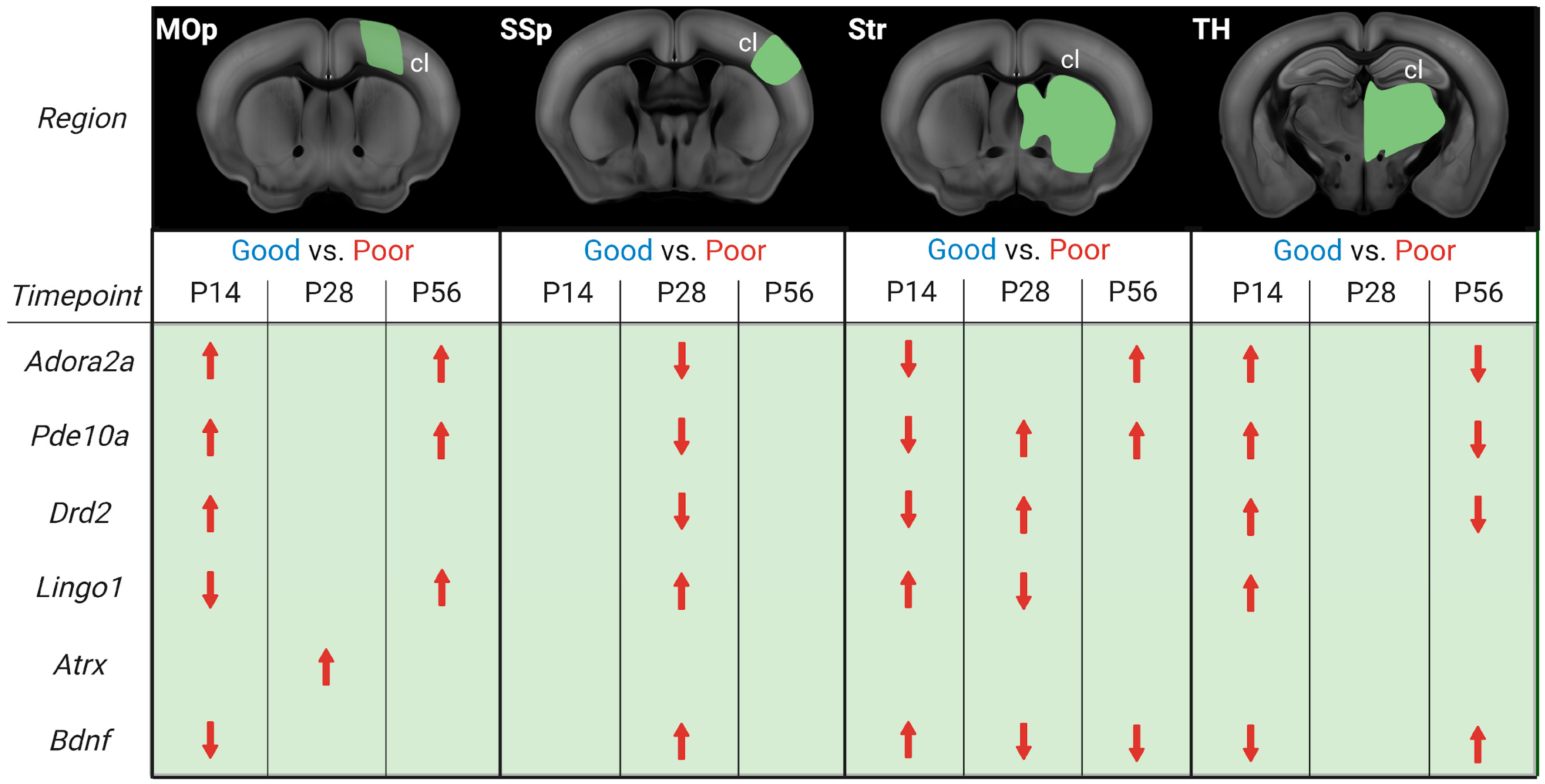
3.2.1. Primary Motor Cortex
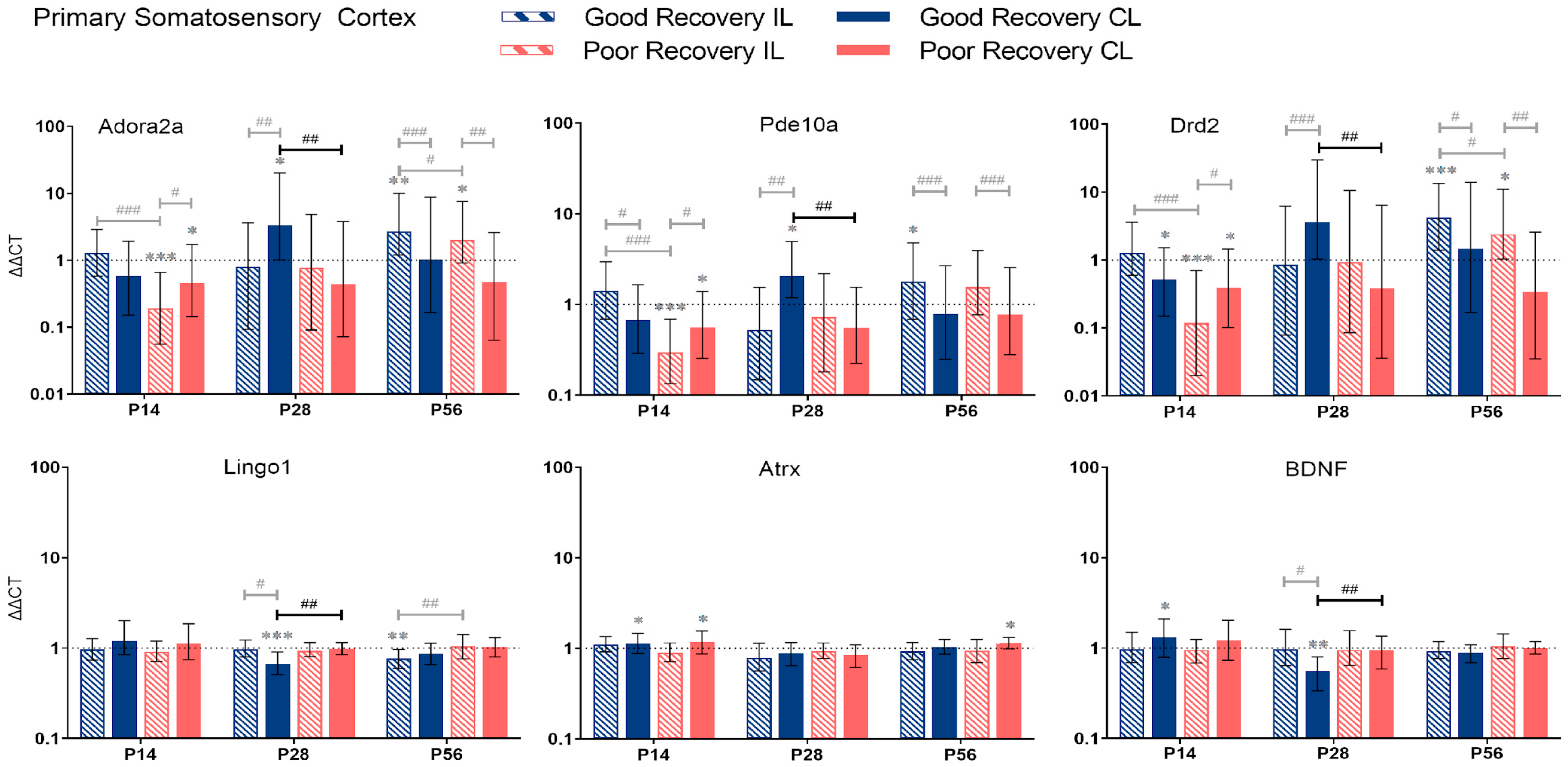
3.2.2. Primary Somatosensory Cortex
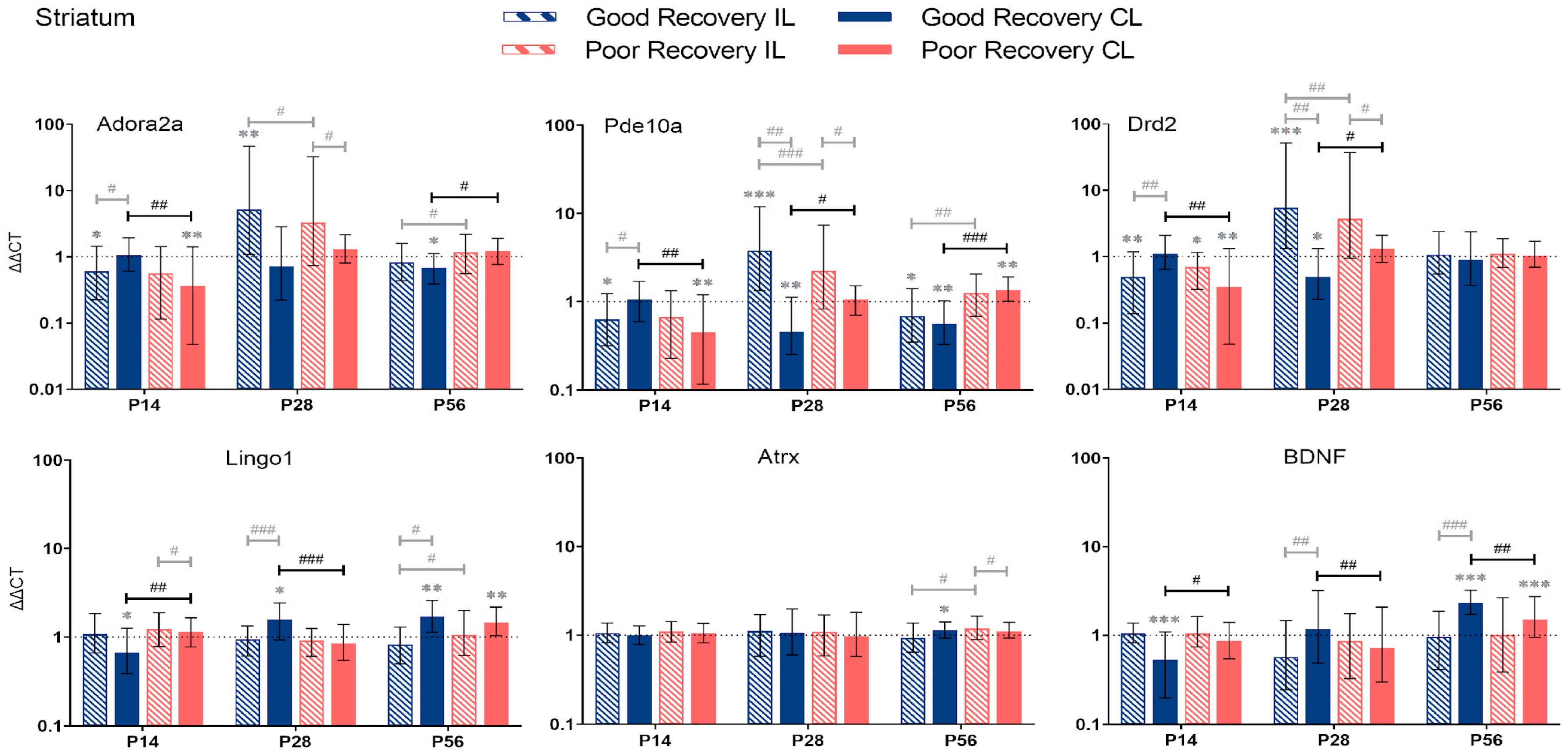
3.2.3. Striatum
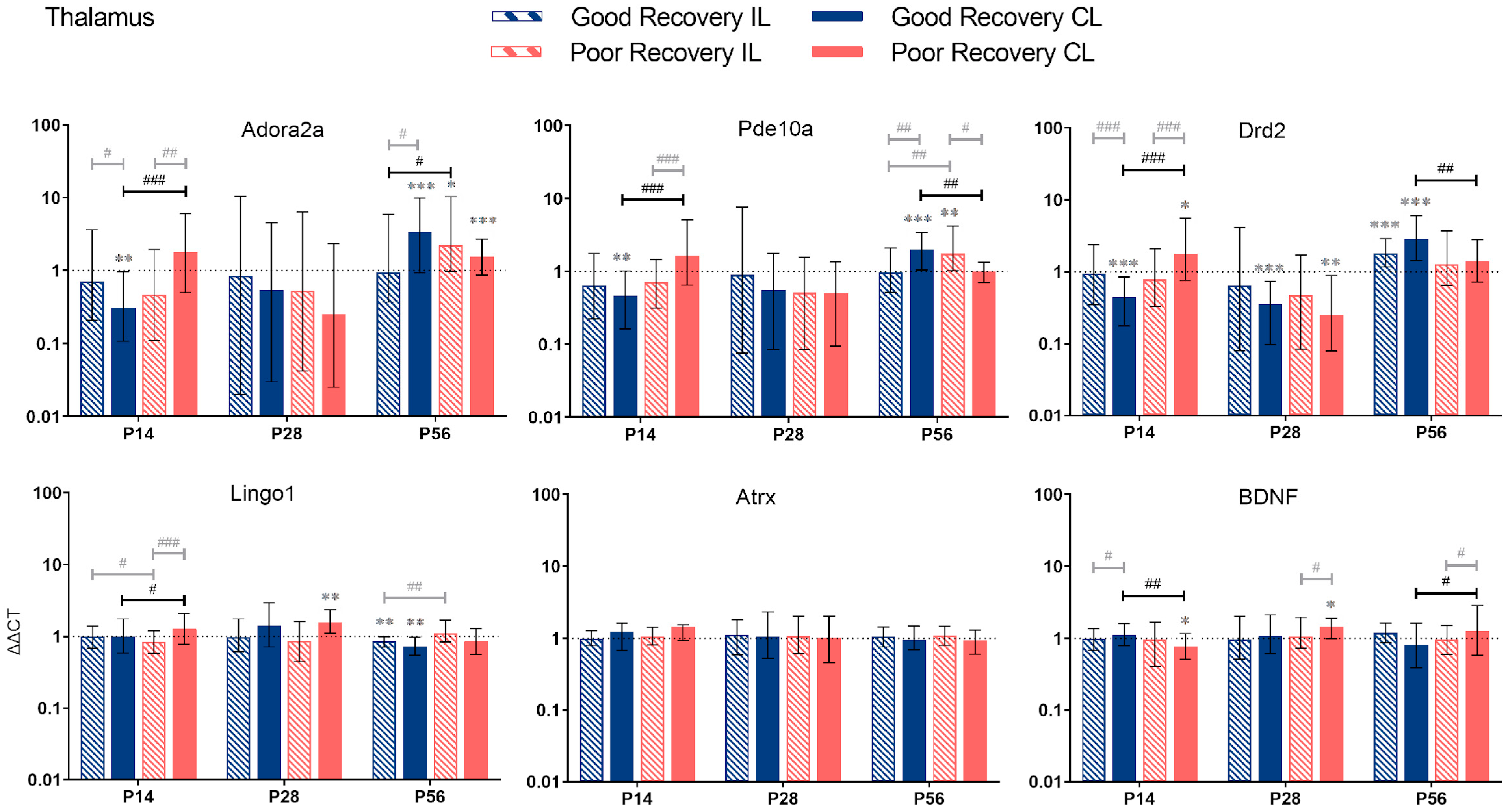
3.2.4. Thalamus
4. Discussion
4.1. Spontaneous Functional Recovery
4.2. Temporal and Spatial Dynamics of Stroke Recovery Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Donkor, E.S. Stroke in the 21(st) Century: A Snapshot of the Burden, Epidemiology, and Quality of Life. Stroke Res. Treat. 2018, 2018, 3238165. [Google Scholar] [PubMed]
- Prabhakaran, S.; Zarahn, E.; Riley, C.; Speizer, A.; Chong, J.Y.; Lazar, R.M.; Marshall, R.S.; Krakauer, J.W. Inter-individual Variability in the Capacity for Motor Recovery After Ischemic Stroke. Neurorehabilit. Neural Repair 2008, 22, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, J.; Hayward, K.; Kwakkel, G.; Ward, N.; Wolf, S.L.; Borschmann, K.; Krakauer, J.W.; Boyd, L.A.; Carmichael, S.T.; Corbett, D.; et al. Agreed definitions and a shared vision for new standards in stroke recovery research: The Stroke Recovery and Rehabilitation Roundtable taskforce. Int. J. Stroke 2017, 12, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Winters, C.; Van Wegen, E.E.H.; Daffertshofer, A.; Kwakkel, G. Generalizability of the Proportional Recovery Model for the Upper Extremity After an Ischemic Stroke. Neurorehabilit. Neural Repair 2015, 29, 614–622. [Google Scholar] [CrossRef]
- Söderholm, M.; Pedersen, A.; Lorentzen, E.; Stanne, T.M.; Bevan, S.; Olsson, M.; Cole, J.W.; Fernandez-Cadenas, I.; Hankey, G.J.; Jimenez-Conde, J.; et al. Genome-wide association meta-analysis of functional outcome after ischemic stroke. Neurology 2019, 92, e1271–e1283. [Google Scholar] [CrossRef]
- Acosta, J.N.; Brown, S.C.; Falcone, G.J. Genetic underpinnings of recovery after stroke: An opportunity for gene discovery, risk stratification, and precision medicine. Genome Med. 2019, 11, 58. [Google Scholar] [CrossRef]
- Androvic, P.; Kirdajova, D.; Tureckova, J.; Zucha, D.; Rohlova, E.; Abaffy, P.; Kriska, J.; Valny, M.; Anderova, M.; Kubista, M.; et al. Decoding the Transcriptional Response to Ischemic Stroke in Young and Aged Mouse Brain. Cell Rep. 2020, 31, 107777. [Google Scholar] [CrossRef]
- Sieber, M.W.; Guenther, M.; Jaenisch, N.; Albrecht-Eckardt, D.; Kohl, M.; Witte, O.W.; Frahm, C. Age-specific transcriptional response to stroke. Neurobiol. Aging 2014, 35, 1744–1754. [Google Scholar] [CrossRef]
- Rakers, C.; Schleif, M.; Blank, N.; Matušková, H.; Ulas, T.; Händler, K.; Torres, S.V.; Schumacher, T.; Tai, K.; Schultze, J.L.; et al. Stroke target identification guided by astrocyte transcriptome analysis. Glia 2019, 67, 619–633. [Google Scholar] [CrossRef]
- Ito, M.; Aswendt, M.; Lee, A.G.; Ishizaka, S.; Cao, Z.; Wang, E.H.; Levy, S.L.; Smerin, D.L.; McNab, J.A.; Zeineh, M.; et al. RNA-Sequencing Analysis Revealed a Distinct Motor Cortex Transcriptome in Spontaneously Recovered Mice After Stroke. Stroke 2018, 49, 2191–2199. [Google Scholar] [CrossRef]
- Pedata, F.; Pugliese, A.M.; Coppi, E.; Dettori, I.; Maraula, G.; Cellai, L.; Melani, A. Adenosine A2A receptors modulate acute injury and neuroinflammation in brain ischemia. Mediat. Inflamm. 2014, 2014, 805198. [Google Scholar] [CrossRef]
- Qiu, J.; Yan, Z.; Tao, K.; Li, Y.; Li, Y.; Li, J.; Dong, Y.; Feng, D.; Chen, H. Sinomenine activates astrocytic dopamine D2 receptors and alleviates neuroinflammatory injury via the CRYAB/STAT3 pathway after ischemic stroke in mice. J. Neuroinflamm. 2016, 13, 263. [Google Scholar] [CrossRef]
- Menniti, F.S.; Chappie, T.; Schmidt, C. PDE10A Inhibitors-Clinical Failure or Window into Antipsychotic Drug Action? Front. Neurosci. 2020, 14, 600178. [Google Scholar] [CrossRef]
- Birjandi, S.Z.; Abduljawad, N.; Nair, S.; Dehghani, M.; Suzuki, K.; Kimura, H.; Carmichael, S.T. Phosphodiesterase 10A Inhibition Leads to Brain Region-Specific Recovery Based on Stroke Type. Transl. Stroke Res. 2021, 12, 303–315. [Google Scholar] [CrossRef]
- Beker, M.C.; Caglayan, A.B.; Altunay, S.; Ozbay, E.; Ates, N.; Kelestemur, T.; Caglayan, B.; Kilic, U.; Doeppner, T.R.; Hermann, D.M.; et al. Phosphodiesterase 10A Is a Critical Target for Neuroprotection in a Mouse Model of Ischemic Stroke. Mol. Neurobiol. 2022, 59, 574–589. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X.; O’Connor, M.; Wang, G.; Han, F. Brain-Derived Neurotrophic Factor and Its Potential Therapeutic Role in Stroke Comorbidities. Neural Plast. 2020, 2020, 1969482. [Google Scholar] [CrossRef]
- Li, S.; Overman, J.J.; Katsman, D.; Kozlov, S.V.; Donnelly, C.J.; Twiss, J.L.; Giger, R.J.; Coppola, G.; Geschwind, D.H.; Carmichael, S.T. An age-related sprouting transcriptome provides molecular control of axonal sprouting after stroke. Nat. Neurosci. 2010, 13, 1496–1504. [Google Scholar] [CrossRef]
- Trifunovski, A.; Josephson, A.; Ringman, A.; Brené, S.; Spenger, C.; Olson, L. Neuronal activity-induced regulation of Lingo-1. Neuroreport 2004, 15, 2397–2400. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Alfieri, A.; Allan, S.M.; Carswell, H.V.; Deuchar, G.A.; Farr, T.D.; Flecknell, P.; Gallagher, L.; Gibson, C.; Haley, M.; et al. The IMPROVE Guidelines (Ischaemia Models: Procedural Refinements Of in Vivo Experiments). J. Cereb. Blood Flow Metab. 2017, 37, 3488–3517. [Google Scholar] [CrossRef]
- Pallast, N.; Wieters, F.; Nill, M.; Fink, G.R.; Aswendt, M. Cloud-based relational database for multimodal animal data. Database 2018, 2018, bay124. [Google Scholar] [CrossRef]
- Aswendt, M.; Pallast, N.; Wieters, F.; Baues, M.; Hoehn, M.; Fink, G.R. Lesion Size- and Location-Dependent Recruitment of Contralesional Thalamus and Motor Cortex Facilitates Recovery after Stroke in Mice. Transl. Stroke Res. 2021, 12, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.Y.; Wang, E.H.; Woodson, W.J.; Wang, S.; Sun, G.; Lee, A.G.; Arac, A.; Fenno, L.E.; Deisseroth, K.; Steinberg, G.K. Optogenetic neuronal stimulation promotes functional recovery after stroke. Proc. Natl. Acad. Sci. USA 2014, 111, 12913–12918. [Google Scholar] [CrossRef] [PubMed]
- Balkaya, M.; Kröber, J.M.; Rex, A.; Endres, M. Assessing Post-Stroke Behavior in Mouse Models of Focal Ischemia. J. Cereb. Blood Flow Metab. 2013, 33, 330–338. [Google Scholar] [CrossRef]
- Baskin, Y.K.; Dietrich, W.; Green, E.J. Two effective behavioral tasks for evaluating sensorimotor dysfunction following traumatic brain injury in mice. J. Neurosci. Methods 2003, 129, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sanberg, P.R.; Li, Y.; Wang, L.; Lu, M.; Willing, A.E.; Sanchez-Ramos, J.; Chopp, M. Intravenous Administration of Human Umbilical Cord Blood Reduces Behavioral Deficits After Stroke in Rats. Stroke 2001, 32, 2682–2688. [Google Scholar] [CrossRef]
- Pallast, N.; Wieters, F.; Nill, M.; Fink, G.R.; Aswendt, M. Graph theoretical quantification of white matter reorganization after cortical stroke in mice. Neuroimage 2020, 217, 116873. [Google Scholar] [CrossRef]
- Pallast, N.; Diedenhofen, M.; Blaschke, S.; Wieters, F.; Wiedermann, D.; Hoehn, M.; Fink, G.R.; Aswendt, M. Processing Pipeline for Atlas-Based Imaging Data Analysis of Structural and Functional Mouse Brain MRI (AIDAmri). Front. Neuroinform. 2019, 13, 42. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Livak, K.J.; TSchmittgen, D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wieloch, T.; Nikolich, K. Mechanisms of neural plasticity following brain injury. Curr. Opin. Neurobiol. 2006, 16, 258–264. [Google Scholar] [CrossRef]
- Aswendt, M.; Wilhelmsson, U.; Wieters, F.; Stokowska, A.; Schmitt, F.J.; Pallast, N.; de Pablo, Y.; Mohammed, L.; Hoehn, M.; Pekna, M.; et al. Reactive astrocytes prevent maladaptive plasticity after ischemic stroke. Prog. Neurobiol. 2022, 209, 102199. [Google Scholar] [CrossRef]
- Parker, K.; Berretta, A.; Saenger, S.; Sivaramakrishnan, M.; Shirley, S.A.; Metzger, F.; Clarkson, A.N. PEGylated insulin-like growth factor-I affords protection and facilitates recovery of lost functions post-focal ischemia. Sci. Rep. 2017, 7, 241. [Google Scholar] [CrossRef]
- Overman, J.J.; Clarkson, A.N.; Wanner, I.B.; Overman, W.T.; Eckstein, I.; Maguire, J.L.; Dinov, I.D.; Toga, A.W.; Carmichael, S.T. A role for ephrin-A5 in axonal sprouting, recovery, and activity-dependent plasticity after stroke. Proc. Natl. Acad. Sci. USA 2012, 109, E2230–E2239. [Google Scholar] [CrossRef]
- Edwards, L.L.; King, E.M.; Buetefisch, C.M.; Borich, M.R. Putting the “Sensory” Into Sensorimotor Control: The Role of Sensorimotor Integration in Goal-Directed Hand Movements After Stroke. Front. Integr. Neurosci. 2019, 13, 16. [Google Scholar] [CrossRef]
- Jones, T.A.; Adkins, D.L. Motor System Reorganization After Stroke: Stimulating and Training Toward Perfection. Physiology 2015, 30, 358–370. [Google Scholar] [CrossRef]
- Ueno, M.; Yamashita, T. Kinematic analyses reveal impaired locomotion following injury of the motor cortex in mice. Exp. Neurol. 2011, 230, 280–290. [Google Scholar] [CrossRef]
- Minassian, A.; Radmilovic, M.D.; Vogel, S.; Diedenhofen, M.; Nelles, M.; Stoeber, M.; Wiedermann, D.; Hoehn, M. Cortical tissue loss and major structural reorganization as result of distal middle cerebral artery occlusion in the chronic phase of nude mice. Sci. Rep. 2019, 9, 6823. [Google Scholar] [CrossRef]
- Wagstaff, K. Clustering with Missing Values: No Imputation Required. In Classification, Clustering, and Data Mining Applications; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Van der Vliet, R.; Selles, R.W.; Andrinopoulou, E.R.; Nijland, R.; Ribbers, G.M.; Frens, M.A.; Meskers, C.; Kwakkel, G. Predicting Upper Limb Motor Impairment Recovery after Stroke: A Mixture Model. Ann. Neurol. 2020, 87, 383–393. [Google Scholar] [CrossRef]
- Schaar, K.L.; Brenneman, M.M.; Savitz, S.I. Functional assessments in the rodent stroke model. Exp. Transl. Stroke Med. 2010, 2, 13. [Google Scholar] [CrossRef]
- Balbinot, G.; Schuch, C.P.; Jeffers, M.S.; McDonald, M.W.; Livingston-Thomas, J.M.; Corbett, D. Post-stroke kinematic analysis in rats reveals similar reaching abnormalities as humans. Sci. Rep. 2018, 8, 8738. [Google Scholar] [CrossRef]
- Weber, R.Z.; Mulders, G.; Kaiser, J.; Tackenberg, C.; Rust, R. Deep learning-based behavioral profiling of rodent stroke recovery. BMC Biol. 2022, 20, 232. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U. Thomas Willis Lecture: Is Translational Stroke Research Broken, and if So, How Can We Fix It? Stroke 2016, 47, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, J.W.; Carmichael, S.T.; Corbett, D.; Wittenberg, G.F. Getting neurorehabilitation right: What can be learned from animal models? Neurorehabil Neural Repair 2012, 26, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Corbett, D.; Carmichael, S.T.; Murphy, T.H.; Jones, T.A.; Schwab, M.E.; Jolkkonen, J.; Clarkson, A.N.; Dancause, N.; Weiloch, T.; Johansen-Berg, H.; et al. Enhancing the alignment of the preclinical and clinical stroke recovery research pipeline: Consensus-based core recommendations from the Stroke Recovery and Rehabilitation Roundtable translational working group. Int. J. Stroke 2017, 12, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Xerri, C.; Zennou-Azogui, Y.; Sadlaoud, K.; Sauvajon, D. Interplay between intra- and interhemispheric remodeling of neural networks as a substrate of functional recovery after stroke: Adaptive versus maladaptive reorganization. Neuroscience 2014, 283, 178–201. [Google Scholar] [CrossRef]
- Brooks, V.B. Motor control. How posture and movements are governed. Phys. Ther. 1983, 63, 664–673. [Google Scholar] [CrossRef]
- Biernaskie, J.; Szymanska, A.; Windle, V.; Corbett, D. Bi-hemispheric contribution to functional motor recovery of the affected forelimb following focal ischemic brain injury in rats. Eur. J. Neurosci. 2005, 21, 989–999. [Google Scholar] [CrossRef]
- Di Pino, G.; Pellegrino, G.; Assenza, G.; Capone, F.; Ferreri, F.; Formica, D.; Ranieri, F.; Tombini, M.; Ziemann, U.; Rothwell, J.C.; et al. Modulation of brain plasticity in stroke: A novel model for neurorehabilitation. Nat. Rev. Neurol. 2014, 10, 597–608. [Google Scholar] [CrossRef]
- Jäger, R.; Russwurm, C.; Schwede, F.; Genieser, H.-G.; Koesling, D.; Russwurm, M. Activation of PDE10 and PDE11 Phosphodiesterases. J. Biol. Chem. 2012, 287, 1210–1219. [Google Scholar] [CrossRef]
- Fujishige, K.; Kotera, J.; Omori, K. Striatum- and testis-specific phosphodiesterase PDE10A isolation and characterization of a rat PDE10A. JBIC J. Biol. Inorg. Chem. 1999, 266, 1118–1127. [Google Scholar]
- Beker, M.C.; Pence, M.E.; Yagmur, S.; Caglayan, B.; Caglayan, A.; Kilic, U.; Yelkenci, H.E.; Altintas, M.O.; Caglayan, A.B.; Doeppner, T.R.; et al. Phosphodiesterase 10A deactivation induces long-term neurological recovery, Peri-infarct remodeling and pyramidal tract plasticity after transient focal cerebral ischemia in mice. Exp. Neurol. 2022, 358, 114221. [Google Scholar] [CrossRef]
- Elder, J.; Cortes, M.; Rykman, A.; Hill, J.; Karuppagounder, S.; Edwards, D.; Ratan, R.R. The Epigenetics of Stroke Recovery and Rehabilitation: From Polycomb to Histone Deacetylases. Neurotherapeutics 2013, 10, 808–816. [Google Scholar] [CrossRef]
- Lu, C.; Wu, X.; Ma, H.; Wang, Q.; Wang, Y.; Luo, Y.; Li, C.; Xu, H. Optogenetic Stimulation Enhanced Neuronal Plasticities in Motor Recovery after Ischemic Stroke. Neural Plast. 2019, 2019, 5271573. [Google Scholar] [CrossRef]
- Lee, X.; Yang, Z.; Shao, Z.; Rosenberg, S.S.; Levesque, M.; Pepinsky, R.B.; Qiu, M.; Miller, R.H.; Chan, J.R.; Mi, S. NGF Regulates the Expression of Axonal LINGO-1 to Inhibit Oligodendrocyte Differentiation and Myelination. J. Neurosci. 2007, 27, 220–225. [Google Scholar] [CrossRef]
- Benowitz, L.I.; Carmichael, S.T. Promoting axonal rewiring to improve outcome after stroke. Neurobiol. Dis. 2010, 37, 259–266. [Google Scholar] [CrossRef]
- Lindau, N.T.; Bänninger, B.J.; Gullo, M.; Good, N.A.; Bachmann, L.C.; Starkey, M.L.; Schwab, M.E. Rewiring of the corticospinal tract in the adult rat after unilateral stroke and anti-Nogo-A therapy. Brain 2014, 137 Pt 3, 739–756. [Google Scholar] [CrossRef]
- Wahl, A.S.; Omlor, W.; Rubio, J.C.; Chen, J.L.; Zheng, H.; Schröter, A.; Gullo, M.; Weinmann, O.; Kobayashi, K.; Helmchen, F.; et al. Neuronal repair. Asynchronous therapy restores motor control by rewiring of the rat corticospinal tract after stroke. Science 2014, 344, 1250–1255. [Google Scholar] [CrossRef]
- Carmichael, S.T.; Kathirvelu, B.; Schweppe, C.A.; Nie, E.H. Molecular, cellular and functional events in axonal sprouting after stroke. Exp. Neurol. 2017, 287 Pt 3, 384–394. [Google Scholar] [CrossRef]
- Fourcaudot, E.; Gambino, F.; Humeau, Y.; Casassus, G.; Shaban, H.; Poulain, B.; Lüthi, A. cAMP/PKA signaling and RIM1α mediate presynaptic LTP in the lateral amygdala. Proc. Natl. Acad. Sci. USA 2008, 105, 15130–15135. [Google Scholar] [CrossRef]
- Qu, H.; Zhao, M.; Zhao, S.; Xiao, T.; Tang, X.; Zhao, D.; Jolkkonen, J.; Zhao, C. Forced limb-use enhances brain plasticity through the cAMP/PKA/CREB signal transduction pathway after stroke in adult rats. Restor. Neurol. Neurosci. 2014, 32, 597–609. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Wu, J.; Manaenko, A.; Yang, P.; Tang, J.; Fu, W.; Zhang, J. Activation of Dopamine D2 Receptor Suppresses Neuroinflammation Through alphaB-Crystalline by Inhibition of NF-kappaB Nuclear Translocation in Experimental ICH Mice Model. Stroke 2015, 46, 2637–2646. [Google Scholar] [CrossRef] [PubMed]
- Huck, J.H.; Freyer, D.; Böttcher, C.; Mladinov, M.; Muselmann-Genschow, C.; Thielke, M.; Gladow, N.; Bloomquist, D.; Mergenthaler, P.; Priller, J. De Novo Expression of Dopamine D2 Receptors on Microglia after Stroke. J. Cereb. Blood Flow Metab. 2015, 35, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Du, R.H.; Zhou, Y.; Xia, M.-L.; Lu, M.; Ding, J.-H.; Hu, G. α-Synuclein disrupts the anti-inflammatory role of Drd2 via interfering β-arrestin2-TAB1 interaction in astrocytes. J. Neuroinflamm. 2018, 15, 258. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Sun, T.; Zhang, J.; Liu, Y.; Wang, D.; Zhu, H.; Yao, H.; Ding, J.; Hu, G.; Lu, M. Drd2 biased agonist prevents neurodegeneration against NLRP3 inflammasome in Parkinson’s disease model via a β-arrestin2-biased mechanism. Brain Behav. Immun. 2020, 90, 259–271. [Google Scholar] [CrossRef]
- Graham, T.A.; Sottoriva, A. Measuring cancer evolution from the genome. J. Pathol. 2017, 241, 183–191. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Remizova, J.A.; Denisova, A.E.; Stavchansky, V.V.; Golovina, K.D.; Gubsky, L.V.; Limborska, S.A.; Dergunova, L.V. Differential gene expression in the contralateral hemisphere of the rat brain after focal ischemia. Sci. Rep. 2023, 13, 573. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Götz, J.; Wieters, F.; Fritz, V.J.; Käsgen, O.; Kalantari, A.; Fink, G.R.; Aswendt, M. Temporal and Spatial Gene Expression Profile of Stroke Recovery Genes in Mice. Genes 2023, 14, 454. https://doi.org/10.3390/genes14020454
Götz J, Wieters F, Fritz VJ, Käsgen O, Kalantari A, Fink GR, Aswendt M. Temporal and Spatial Gene Expression Profile of Stroke Recovery Genes in Mice. Genes. 2023; 14(2):454. https://doi.org/10.3390/genes14020454
Chicago/Turabian StyleGötz, Jan, Frederique Wieters, Veronika J. Fritz, Olivia Käsgen, Aref Kalantari, Gereon R. Fink, and Markus Aswendt. 2023. "Temporal and Spatial Gene Expression Profile of Stroke Recovery Genes in Mice" Genes 14, no. 2: 454. https://doi.org/10.3390/genes14020454
APA StyleGötz, J., Wieters, F., Fritz, V. J., Käsgen, O., Kalantari, A., Fink, G. R., & Aswendt, M. (2023). Temporal and Spatial Gene Expression Profile of Stroke Recovery Genes in Mice. Genes, 14(2), 454. https://doi.org/10.3390/genes14020454