

Maternal Epigenetic Dysregulation as a Possible Risk Factor for Neurodevelopmental Disorders

 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. The Experimental Sample
2.2. Sample Collection and DNA Isolation
2.3. Bisulfite Treatment and High-Resolution Melting Analysis
2.4. Statistical Analysis
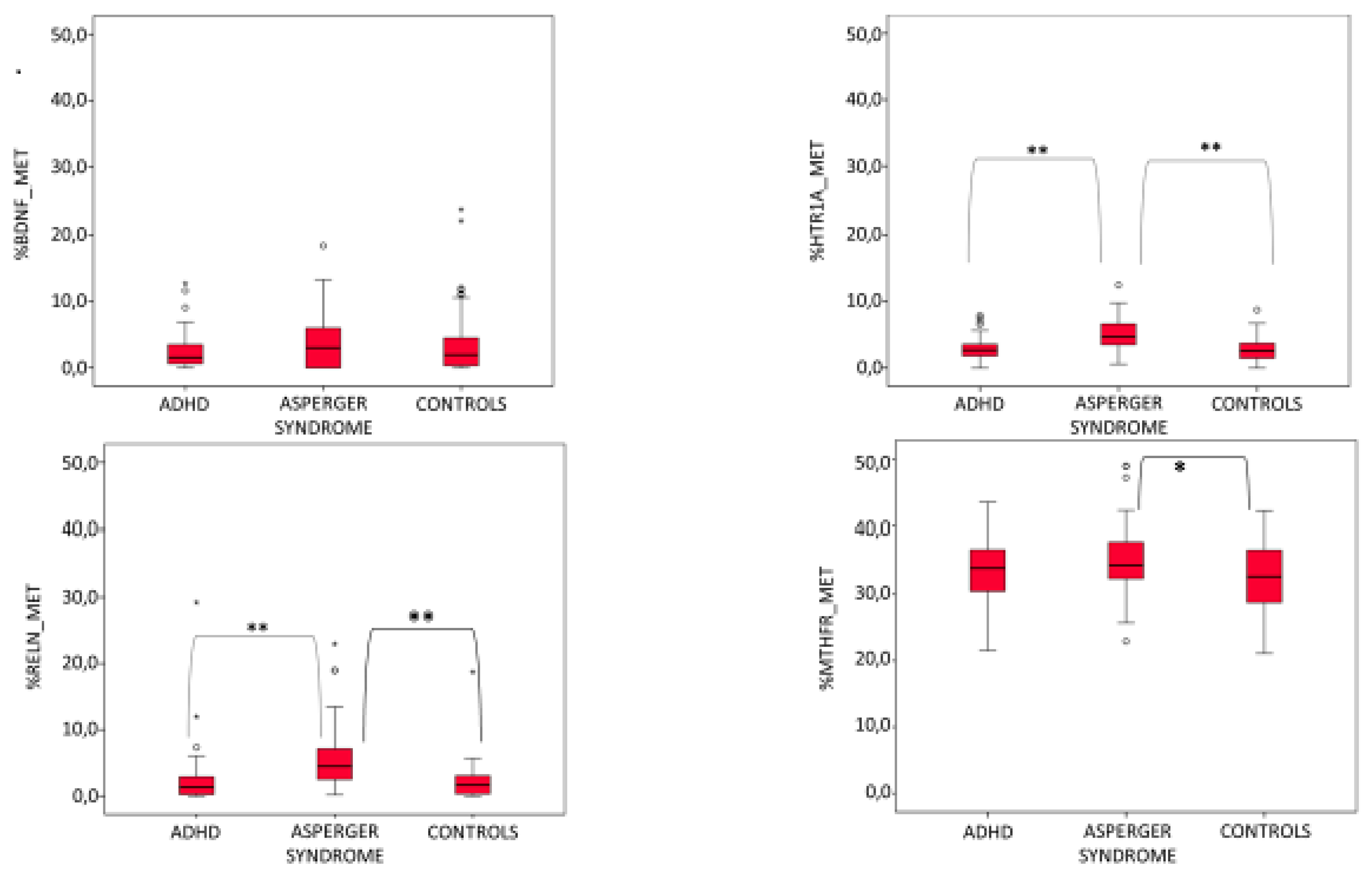
3. Results
3.1. AS Mothers Display a Higher Methylation Level Than ADHD and Healthy Control Mothers
3.2. Mean Percentage of Methylation Correlates with Some Clinical Variables
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5); American Psychiatric Publication: Washington, DC, USA, 2013. [Google Scholar]
- Hayman, V.; Fernandez, T.V. Genetic Insights Into ADHD Biology. Front. Psychiatry 2018, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.K.; Stanton, J.E.; Hans, S.; Grabrucker, A.M. Autism Spectrum Disorders: Etiology and Pathology. In Autism Spectrum Disorders [Internet]; Grabrucker, A.M., Ed.; Exon Publications: Brisbane, Australia, 2021. [Google Scholar]
- Faraone, S.V.; Perlis, R.H.; Doyle, A.E.; Smoller, J.W.; Goralnick, J.J.; Holmgren, M.A.; Sklar, P. Molecular genetics of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1313–1323. [Google Scholar] [CrossRef]
- Faraone, S.V.; Mick, E. Molecular genetics of attention deficit hyperactivity disorder. Psychiatr. Clin. North Am. 2010, 33, 159–180. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Tran, P.V.; Carlson, E.S. Atypical fetal development: Fetal alcohol syndrome, nutritional deprivation, teratogens, and risk for neurodevelopmental disorders and psychopathology. Dev. Psychopathol. 2018, 30, 1063–1086. [Google Scholar] [CrossRef]
- Rice, F.; Langley, K.; Woodford, C.; Davey Smith, G.; Thapar, A. Identifying the contribution of prenatal risk factors to offspring development and psychopathology: What designs to use and a critique of literature on maternal smoking and stress in pregnancy. Dev. Psychopathol. 2018, 30, 1107–1128. [Google Scholar] [CrossRef]
- Saez, M.; Barceló, M.A.; Farrerons, M.; López-Casasnovas, G. The association between exposure to environmental factors and the occurrence of attention-deficit/hyperactivity disorder (ADHD). A population-based retrospective cohort study. Environ. Res. 2018, 166, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Jeličić, L.; Veselinović, A.; Ćirović, M.; Jakovljević, V.; Raičević, S.; Subotić, M. Maternal Distress during Pregnancy and the Postpartum Period: Underlying Mechanisms and Child’s Developmental Outcomes-A Narrative Review. Int. J. Mol. Sci. 2022, 23, 13932. [Google Scholar] [CrossRef]
- Middeldorp, C.M.; Slof-Op’t Landt, M.C.; Medland, S.E.; van Beijsterveldt, C.E.; Bartels, M.; Willemsen, G.; Hottenga, J.J.; de Geus, E.J.; Suchiman, H.E.; Dolan, C.V.; et al. Anxiety and depression in children and adults: Influence of serotonergic and neurotrophic genes? Genes Brain Behav. 2010, 9, 808–816. [Google Scholar] [CrossRef]
- Chen, R.J.; Nabila, A.; Phalke, S.; Castro, D.F.; Toth, J.G.; Bergin, P.; Bastiaans, J.; Stuhlmann, H.; Pernis, A.B.; Toth, M. Serotonin-1A receptor, a psychiatric disease risk factor, influences offspring immunity via sex-dependent genetic nurture. iScience 2022, 25, 105595. [Google Scholar] [CrossRef] [PubMed]
- Blakeley, P.M.; Capron, L.E.; Jensen, A.B.; O’Donnell, K.J.; Glover, V. Maternal prenatal symptoms of depression and down regulation of placental monoamine oxidase A expression. J. Psychosom. Res. 2013, 75, 341–345. [Google Scholar] [CrossRef]
- Förster, E.; Bock, H.H.; Herz, J.; Chai, X.; Frotscher, M.; Zhao, S. Emerging topics in Reelin function. Eur. J. Neurosci. 2010, 31, 1511–1518. [Google Scholar] [CrossRef]
- Lintas, C.; Sacco, R.; Persico, A.M. Differential methylation at the RELN gene promoter in temporal cortex from autistic and typically developing post-puberal subjects. J. Neurodev. Disord. 2016, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Quattrocchi, C.C.; Wannenes, F.; Persico, A.M.; Ciafré, S.A.; D’Arcangelo, G.; Farace, M.G.; Keller, F. Reelin is a serine protease of the extracellular matrix. J. Biol. Chem. 2002, 277, 303–309. [Google Scholar] [CrossRef]
- Sahay, A.; Kale, A.; Joshi, S. Role of neurotrophins in pregnancy and offspring brain development. Neuropeptides 2020, 83, 102075. [Google Scholar] [CrossRef]
- D’Souza, V.; Patil, V.; Pisal, H.; Randhir, K.; Joshi, A.; Mehendale, S.; Wagh, G.; Gupte, S.; Joshi, S. Levels of brain derived neurotrophic factors across gestation in women with preeclampsia. Int. J. Dev. Neurosci. 2014, 37, 6–40. [Google Scholar] [CrossRef] [PubMed]
- Maher, G.M.; O’Keeffe, G.W.; Kearney, P.M.; Kenny, L.C.; Dinan, T.G.; Mattsson, M.; Khashan, A.S. Association of hypertensive disorders of pregnancy with risk of neurodevelopmental disorders in offspring: A systematic review and meta-analysis. JAMA Psychiatry 2018, 75, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Craciunescu, C.N.; Brown, E.C.; Mar, M.H.; Albright, C.D.; Nadeau, M.R.; Zeisel, S.H. Folic acid deficiency during late gestation decreases progenitor cell proliferation and increases apoptosis in fetal mouse brain. J. Nutr. 2004, 134, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Pulikkunnel, S.T.; Thomas, S.V. Neural tube defects: Pathogenesis and folate metabolism. J. Assoc. Physicians India 2005, 53, 127–135. [Google Scholar] [PubMed]
- Christensen, B.; Rosenblatt, D.S. Effects of folate deficiency on embryonic development. Baillieres. Clin. Haematol. 1995, 8, 617–637. [Google Scholar] [CrossRef]
- Schlotz, W.; Jones, A.; Phillips, D.I.; Gale, C.R.; Robinson, S.M.; Godfrey, K.M. Lower maternal folate status in early pregnancy is associated with childhood hyperactivity and peer problems in offspring. J. Child Psychol. Psychiatry 2010, 51, 594–602. [Google Scholar] [CrossRef]
- DeVilbiss, E.A.; Gardner, R.M.; Newschaffer, C.J.; Lee, B.K. Maternal folate status as a risk factor for autism spectrum disorders: A review of existing evidence. Br. J. Nutr. 2015, 114, 663–672. [Google Scholar] [CrossRef]
- Wagner, C. Biochemical role of folate in cellular metabolism. In Folate in Health and Disease; Bailey, L., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 23–42. [Google Scholar]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA methylation: A review of molecular mechanisms and the evidence for folate’s role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef]
- Stover, P.J. Polymorphisms in 1-carbon metabolism, epigenetics and folate-related pathologies. J. Nutr. Nutr. 2011, 4, 293–305. [Google Scholar] [CrossRef]
- Lintas, C. Linking genetics to epigenetics: The role of folate and folate-related pathways in neurodevelopmental disorders. Clin. Genet. 2019, 95, 241–252. [Google Scholar] [CrossRef]
- Hu, W.F.; Chahrour, M.H.; Walsh, C.A. The diverse genetic landscape of neurodevelopmental disorders. Annu. Rev. Genom. Hum. Genet. 2014, 15, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.; Mukamel, E.A.; Nery, J.R.; Urich, M.; Puddifoot, C.A.; Johnson, N.D.; Lucero, J.; Huang, Y.; Dwork, A.J.; Schultz, M.D.; et al. Global epigenomic reconfiguration during mammalian brain development. Science 2013, 341, 1237905. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Suetake, I.; Tada, T.; Tajima, S. Stage- and cell-specific expression of Dnmt3a and Dnmt3b during embryogenesis. Mech. Dev. 2002, 118, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R. The Griffiths Mental Development Scales from Birth to 2 Years, Manual, the 1996 Revision; Association for Research in Infant and Child Development, Test Agency: Henley, UK, 1996. [Google Scholar]
- Achenbach, T.M. Manual for Child Behavior Checklist/4-18 and Profile; University of Vermont, Department of Psychiatry: Burlington, VT, USA, 1991. [Google Scholar]
- Goyette, C.H.; Conners, C.K.; Ulrich, R.F. Normative data on revised Conners Parent and Teacher Rating Scales. J. Abnorm. Child Psychol. 1978, 6, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Rutter, M.; DiLavore, P.; Risi, S.; Gotham, K.; Bishop, S. Autism Diagnostic Observation Schedule, (ADOS-2) Modules 1-4, 2nd ed.; Western Psychological Services: Torrance, CA, USA, 2012. [Google Scholar]
- Rutter, M.; Le Couteur, A.; Lord, C. ADI-R, Autism Diagnostic Interview-Revised; Western Psychological Services: Los Angeles, CA, USA, 2003. [Google Scholar]
- Tannorella, P.; Stoccoro, A.; Tognoni, G.; Petrozzi, L.; Salluzzo, M.G.; Ragalmuto, A.; Siciliano, G.; Haslberger, A.; Bosco, P.; Bonuccelli, U.; et al. Methylation analysis of multiple genes in blood DNA of Alzheimer’s disease and healthy individuals. Neurosci. Lett. 2015, 600, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Stenz, L.; Zewdie, S.; Laforge-Escarra, T.; Prados, J.; La Harpe, R.; Dayer, A.; Paoloni-Giacobino, A.; Perroud, N.; Aubry, J.-M. BDNF promoter I methylation correlates between post-mortem human peripheral and brain tissues. Neurosci. Res. 2015, 91, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.; Stoccoro, A.; Cagiano, R.; Nicolì, V.; Ricciardi, R.; Tancredi, R.; Trovato, R.; Santorelli, F.M.; Calderoni, S.; Muratori, F.; et al. Correlation among maternal risk factors, gene methylation and disease severity in females with autism spectrum disorder. Epigenomics 2022, 14, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Migheli, F.; Stoccoro, A.; Coppedè, F.; Omar, W.A.W.; Failli, A.; Consolini, R.; Seccia, M.; Spisni, R.; Miccoli, P.; Mathers, J.C.; et al. Comparison study of MS-HRM and pyrosequencing techniques for quantification of APC and CDKN2A gene methylation. PLoS ONE 2013, 8, e52501. [Google Scholar] [CrossRef]
- Sivasangari, K.; Rajan, E.K. Prenatal exposure to valproic acid alters Reelin, NGF expressing neuron architecture and impairs social interaction in their autistic-like phenotype male offspring. Exp. Brain Res. 2022, 240, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Pathak, A.; Sinha, R.A.; Mohan, V.; Mitra, K.; Godbole, M.M. Maternal thyroid hormone before the onset of fetal thyroid function regulates reelin and downstream signaling cascade affecting neocortical neuronal migration. Cereb. Cortex. 2011, 21, 11–21. [Google Scholar] [CrossRef]
- Su, Y.; Yang, X.; Yang, L.; Liu, X.; She, Z.; Zhang, Y.; Dong, Z. Thyroid hormones regulate reelin expression in neuropsychiatric disorders. Can. J. Physiol. Pharmacol. 2022, 100, 1033–1044. [Google Scholar] [CrossRef]
- Dutra, M.L.; Dias, P.; Freiberger, V.; Ventura, L.; Comim, C.M.; Martins, D.F.; Bobinski, F. Maternal immune activation induces autism-like behavior and reduces brain-derived neurotrophic factor levels in the hippocampus and offspring cortex of C57BL/6 mice. Neurosci. Lett. 2022, 793, 136974. [Google Scholar] [CrossRef] [PubMed]
- Kouter, K.; Zupanc, T.; Videtič Paska, A. Targeted sequencing approach: Comprehensive analysis of DNA methylation and gene expression across blood and brain regions in suicide victims. World J. Biol. Psychiatry 2022, 9, 12–23. [Google Scholar] [CrossRef]
- Iurato, S.; Carrillo-Roa, T.; Arloth, J.; Czamara, D.; Diener-Hölzl, L.; Lange, J.; Müller-Myhsok, B.; Binder, E.B.; Erhardt, A. DNA Methylation signatures in panic disorder. Transl. Psychiatry 2017, 7, 1287. [Google Scholar] [CrossRef]
- Schneider, I.; Kugel, H.; Redlich, R.; Grotegerd, D.; Bürger, C.; Bürkner, P.-C.; Opel, N.; Dohm, K.; Zaremba, D.; Meinert, S.; et al. Association of Serotonin Transporter Gene AluJb Methylation with Major Depression, Amygdala Responsiveness, 5-HTTLPR/rsPolymorphism, and Stress. Neuropsychopharmacology 2018, 6, 1308–1316. [Google Scholar] [CrossRef]
- Folsom, T.D.; Fatemi, S.H. The involvement of Reelin in neurodevelopmental disorder. Neuropharmacology 2013, 68, 122–135. [Google Scholar] [CrossRef]
- Stoccoro, A.; Gallo, R.; Calderoni, S.; Cagiano, R.; Muratori, F.; Migliore, L.; Grossi, E.; Coppedè, F. Artificial neural networks reveal sex differences in gene methylation, and connections between maternal risk factors and symptom severity in autism spectrum disorder. Epigenomics 2022, 14, 1181–1195. [Google Scholar] [CrossRef]
- Costa-Júnior, J.M.; Ferreira, S.M.; Kurauti, M.A.; Bernstein, D.L.; Ruano, E.G.; Kameswaran, V.; Schug, J.; Freitas-Dias, R.; Zoppi, C.C.; Boschero, A.C.; et al. Paternal Exercise Improves the Metabolic Health of Offspring via Epigenetic Modulation of the Germline. Int. J. Mol. Sci. 2021, 23, 1. [Google Scholar] [CrossRef]
- Fitz-James, M.H.; Cavalli, G. Molecular mechanisms of transgenerational epigenetic inheritance. Nat. Rev. Genet. 2022, 23, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.J.; Lage, S.; Caruncho, H.J.; Cadavid, M.I.; Loza, M.I.; Brea, J. Reelin influences the expression and function of dopamine D2 and serotonin 5-HT2A receptors: A comparative study. Neuroscience 2015, 290, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Piras, I.S.; Costa, A.; Tirindelli, M.C.; Stoccoro, A.; Huentelman, M.J.; Sacco, R.; Coppedè, F.; Lintas, C. Genetic and epigenetic MTHFR gene variants in the mothers of attention-deficit/hyperactivity disorder affected children as possible risk factors for neurodevelopmental disorders. Epigenomics 2020, 12, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Qiu, Y.; Li, Y.; Cong, X. Genetics of autism spectrum disorder: An umbrella review of systematic reviews and meta-analyses. Transl. Psychiatry 2022, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Mpoulimari, I.; Zintzaras, E. Synthesis of genetic association studies on autism spectrum disorders using a genetic model-free approach. Psychiatr. Genet. 2022, 32, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Wang, J.; Zhang, F.; Diao, B.; Song, Z.F.; Shan, L.L.; Wang, W.; Cao, H.J.; Li, X.Q. Correlation between MTHFR gene methylation and pre-eclampsia, and its clinical significance. Genet. Mol. Res. 2015, 14, 8021–8028. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Bosi, S.; Bazzan, E.; Di Domenico, M.; De Mattei, M.; Selvatici, R.; Patella, A.; Marci, R.; Tognon, M.; Martini, F. Methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples of infertile couples correlates with recurrent spontaneous abortion. Hum. Reprod. 2012, 27, 3632–3638. [Google Scholar] [CrossRef]
- Saraswathy, K.N.; Kaur, L.; Talwar, S.; Mishra, J.; Huidrom, S.; Sachdeva, M.P.; Puri, M. Methylenetetrahydrofolate Reductase Gene-specific Methylation and Recurrent Miscarriages: A Case-Control Study from North India. J. Hum. Reprod. Sci. 2018, 11, 142–147. [Google Scholar] [CrossRef]
- Coppedè, F.; Denaro, M.; Tannorella, P.; Migliore, L. Increased MTHFR promoter methylation in mothers of Down syndrome individuals. Mutat. Res. 2016, 787, 1–6. [Google Scholar] [CrossRef]
- Asim, A.; Agarwal, S.; Panigrahi, I.; Saiyed, N.; Bakshi, S. MTHFR promoter hypermethylation may lead to congenital heart defects in Down syndrome. Intractable Rare Dis. Res. 2017, 6, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Khazamipour, N.; Noruzinia, M.; Fatehmanesh, P.; Keyhanee, M.; Pujol, P. MTHFR promoter hypermethylation in testicular biopsies of patients with non-obstructive azoospermia: The role of epigenetics in male infertility. Hum. Reprod. 2009, 24, 2361–2364. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Selvatici, R.; Di Domenico, M.; Marci, R.; Vesce, F.; Tognon, M. Methylation loss at H19 imprinted gene correlates with methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples from infertile males. Epigenetics 2013, 8, 990–997. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Genes | Primer Sequences | Number of CpGs in the Amplified Region | Anealling Temperature (°C) | PCR Product Size (bp) |
|---|---|---|---|---|
| MTHFR | 5′-TTTTAATTTTTGTTTGGAGGGTAGT-3′ (for) | 7 | 55 | 155 |
| 5′-AAAAAAACCACTTATCACCAAATTC-3′ (rev) | ||||
| BDNF | 5′-GGGTTGTTAATTTATATTTGGGAAGT-3′ (for) | 4 | 58 | 119 |
| 5′-AACCACTAATTACCCACAAAAACC-3′ (rev) | ||||
| HTR1A | 5′-TGTTTGTTAGTGGGGAGATTTTAGT-3′ (for) | 15 | 52 | 251 |
| 5′-CAAAAACCCAAACAAAAAATTCTTA-3′ (rev) | ||||
| RELN | 5′-TTGAAGAGTTTAGAAGTAATGAATAATAGA-3′ (for) | 7 | 56 | 192 |
| 5′-ACCTCATCTATAAAAAATTTTAAAATAAAA-3′ (rev) |
| Status | N. | Mean ± SD | df | F | Sig. | |
|---|---|---|---|---|---|---|
| %BDNF_MET | ADHD | 45 | 2.5 ± 3.0 | 2 | 1.608 | 0.204 |
| ASD | 43 | 4.1 ± 4.7 | ||||
| CTRL | 45 | 3.9 ± 5.6 | ||||
| Tot. | 133 | 3.5 ± 4.6 | ||||
| %HTR1A_MET | ADHD | 45 | 3.0 ± 1.9 | 2 | 14.080 | 0.000 |
| ASD | 45 | 5.0 ± 2.7 | ||||
| CTRL | 45 | 2.7 ± 1.8 | ||||
| Tot. | 135 | 3.6 ± 2.4 | ||||
| %RELN_MET | ADHD | 45 | 2.6 ± 4.7 | 2 | 8.240 | 0.000 |
| ASD | 44 | 5.7 ± 5.0 | ||||
| CTRL | 45 | 2.3 ± 3.1 | ||||
| Tot. | 134 | 3.5 ± 4.6 | ||||
| %MTHFR_MET | ADHD | 45 | 33.4 ± 4.8 | 2 | 2.978 | 0.054 |
| ASD | 44 | 35.4 ± 5.6 | ||||
| CTRL | 45 | 32.7 ± 5.6 | ||||
| Tot. | 134 | 33.8 ± 5.4 |
| Newborn Birth Weight | Active Smoking | Passive Smoking | Alcohol Use | Drug Use | Threatened Miscarriage | Obstetric Complications- Mother | Obstetric Complications- Newborn | Pregnancy Duration | Exposure to Toxic/Polluting Agents | Family History of Neuropsychiatric Disorders | Autoimmune/ Inflammatory Diseases | Infections | Psychological Trauma | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| %BDNF_MET | ADHD | Tau | −0.387 | 0.017 | 0.260 | −0.154 | 0.244 | 0.409 | 0.205 | 0.204 | 0.191 | 0.007 | −0.033 | 0.035 | 0.356 | 0.218 |
| Sig. | 0.008 | 0.920 | 0.128 | 0.367 | 0.154 | 0.017 | 0.232 | 0.234 | 0.266 | 0.967 | 0.849 | 0.838 | 0.038 | 0.203 | ||
| N. | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | ||
| ASD | Tau | −0.019 | 0.225 | −0.073 | 0.000 | −0.063 | −0.507 | −0.133 | −0.034 | 0.364 | −0.090 | 0.418 | ||||
| Sig. | 0.873 | 0.387 | 0.780 | 1.000 | 0.794 | 0.052 | 0.390 | 0.805 | 0.163 | 0.550 | 0.109 | |||||
| N. | 39 | 12 | 12 | 12 | 12 | 14 | 12 | 31 | 36 | 12 | 12 | 33 | 11 | 12 | ||
| CTRL | Tau | −0.109 | −0.249 | 0.147 | −0.206 | −0.156 | −0.031 | 0.052 | −0.217 | −0.180 | −0.166 | −0.091 | −0.156 | −0.249 | −0.142 | |
| Sig. | 0.401 | 0.100 | 0.332 | 0.174 | 0.303 | 0.837 | 0.731 | 0.152 | 0.230 | 0.273 | 0.549 | 0.303 | 0.100 | 0.348 | ||
| N. | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | ||
| %HTR1A_MET | ADHD | Tau | 0.000 | 0.102 | −0.025 | −0.236 | −0.309 | 0.198 | −0.069 | −0.087 | −0.139 | −0.213 | 0.055 | −0.030 | 0.025 | −0.195 |
| Sig. | 1.000 | 0.548 | 0.882 | 0.166 | 0.069 | 0.244 | 0.683 | 0.610 | 0.415 | 0.210 | 0.744 | 0.861 | 0.882 | 0.252 | ||
| N. | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | ||
| ASD | Tau | −0.274 | 0.145 | −0.372 | 0.189 | 0.505 | −0.097 | 0.216 | −0.153 | −0.261 | −0.041 | 0.042 | −0.334 | 0.261 | ||
| Sig. | 0.013 | 0.554 | 0.128 | 0.440 | 0.025 | 0.693 | 0.141 | 0.245 | 0.285 | 0.866 | 0.768 | 0.192 | 0.285 | |||
| N. | 41 | 13 | 13 | 13 | 13 | 15 | 13 | 33 | 38 | 13 | 13 | 35 | 12 | 13 | ||
| CTRL | Tau | −0.073 | 0.151 | 0.133 | 0.040 | −0.289 | −0.027 | 0.187 | −0.327 | −0.033 | −0.081 | 0.035 | 0.178 | −0.052 | 0.058 | |
| Sig. | 0.567 | 0.311 | 0.372 | 0.787 | 0.053 | 0.855 | 0.210 | 0.028 | 0.821 | 0.586 | 0.815 | 0.231 | 0.726 | 0.697 | ||
| N. | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | ||
| %RELN_MET | ADHD | Tau | −0.054 | −0.224 | −0.019 | −0.250 | −0.156 | 0.005 | 0.005 | −0.078 | 0.178 | −0.144 | −0.033 | −0.035 | 0.019 | 0.131 |
| Sig. | 0.707 | 0.192 | 0.911 | 0.145 | 0.363 | 0977 | 0.977 | 0.650 | 0.298 | 0.402 | 0.849 | 0.838 | 0.911 | 0.444 | ||
| N. | 25 | 25 | 25 | 25 | 29 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | ||
| ASD | Tau | 0.036 | 0.097 | 0.083 | 0.075 | −0.345 | 0.048 | −0.092 | −0.187 | −0.065 | 0.207 | 0.156 | −0.111 | −0.261 | ||
| Sig. | 0.744 | 0.693 | 0.735 | 0.758 | 0.126 | 0.844 | 0.531 | 0.161 | 0.789 | 0.398 | 0.280 | 0.664 | 0.285 | |||
| N. | 40 | 13 | 13 | 13 | 13 | 15 | 13 | 33 | 37 | 13 | 13 | 34 | 12 | 13 | ||
| CTRL | Tau | −0.237 | −0.230 | −0.048 | −0.213 | −0.263 | −0.035 | 0.147 | −0.073 | −0.179 | −0.082 | 0.145 | −0.013 | −0.230 | 0.035 | |
| Sig. | 0.066 | 0.128 | 0.753 | 0.158 | 0.081 | 0.819 | 0.331 | 0.627 | 0.231 | 0.585 | 0.335 | 0.932 | 0.128 | 0.815 | ||
| N. | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | ||
| %MTHFR_MET | ADHD | Tau | −0.094 | −0.238 | 0.063 | −0.189 | 0.000 | 0.248 | 0.208 | −0.173 | −0.050 | −0.114 | 0.065 | 0.050 | 0.025 | −0.119 |
| Sig. | 0.512 | 0.161 | 0.711 | 0.267 | 1.000 | 0.145 | 0.221 | 0.308 | 0.767 | 0.503 | 0.703 | 0.771 | 0.882 | 0.484 | ||
| N. | 25 | 25 | 25 | 29 | 29 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | ||
| ASD | Tau | 0.003 | −0.055 | −0.143 | 0.477 | 0.091 | 0.000 | −0.071 | −0.172 | −0.112 | 0.072 | 0.143 | −0.172 | −0.187 | ||
| Sig. | 0.981 | 0.830 | 0.578 | 0.064 | 0.698 | 1.000 | 0.631 | 0.200 | 0.663 | 0.781 | 0.322 | 0.526 | 0.468 | |||
| N. | 40 | 12 | 12 | 12 | 12 | 14 | 12 | 32 | 37 | 12 | 12 | 34 | 11 | 12 | ||
| CTRL | Tau | −0.063 | −0.174 | 0.137 | −0.234 | −0.076 | −0.010 | −0.042 | 0.051 | 0.129 | 0.290 | −0.058 | 0.093 | 0.151 | 0.162 | |
| Sig. | 0.623 | 0.243 | 0.359 | 0.116 | 0.608 | 0.945 | 0.776 | 0.732 | 0.383 | 0.052 | 0.697 | 0.531 | 0.312 | 0.276 | ||
| N. | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lintas, C.; Cassano, I.; Azzarà, A.; Stigliano, M.G.; Gregorj, C.; Sacco, R.; Stoccoro, A.; Coppedè, F.; Gurrieri, F. Maternal Epigenetic Dysregulation as a Possible Risk Factor for Neurodevelopmental Disorders. Genes 2023, 14, 585. https://doi.org/10.3390/genes14030585
Lintas C, Cassano I, Azzarà A, Stigliano MG, Gregorj C, Sacco R, Stoccoro A, Coppedè F, Gurrieri F. Maternal Epigenetic Dysregulation as a Possible Risk Factor for Neurodevelopmental Disorders. Genes. 2023; 14(3):585. https://doi.org/10.3390/genes14030585
Chicago/Turabian StyleLintas, Carla, Ilaria Cassano, Alessia Azzarà, Maria Grazia Stigliano, Chiara Gregorj, Roberto Sacco, Andrea Stoccoro, Fabio Coppedè, and Fiorella Gurrieri. 2023. "Maternal Epigenetic Dysregulation as a Possible Risk Factor for Neurodevelopmental Disorders" Genes 14, no. 3: 585. https://doi.org/10.3390/genes14030585
APA StyleLintas, C., Cassano, I., Azzarà, A., Stigliano, M. G., Gregorj, C., Sacco, R., Stoccoro, A., Coppedè, F., & Gurrieri, F. (2023). Maternal Epigenetic Dysregulation as a Possible Risk Factor for Neurodevelopmental Disorders. Genes, 14(3), 585. https://doi.org/10.3390/genes14030585