
Mitochondrial Genomes Assembled from Non-Invasive eDNA Metagenomic Scat Samples in Critically Endangered Mammals

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequencing of Mitochondrial Genomes
2.2. Mitochondrial Genome Assembly
2.3. Mitochondrial Genome Annotation and Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dinerstein, E. Global and Local Conservation Priorities. Science 2007, 318, 1377–1382. [Google Scholar] [CrossRef]
- Joshi, A.R.; Dinerstein, E.; Wikramanayake, E.; Anderson, M.L.; Olson, D.; Jones, B.S.; Seidensticker, J.; Lumpkin, S.; Hansen, M.C.; Sizer, N.C.; et al. Tracking changes and preventing loss in critical tiger habitat. Sci. Adv. 2016, 2, e1501675. [Google Scholar] [CrossRef] [Green Version]
- Stephen, C.; Duff, J.; Gavier-Widen, D.; Ryser-Degiorgis, M.; Uhart, M.; Sleeman, J.; Nguyen, N.; Zimmer, P.; Grillo, T.; Lee, H.; et al. Proposed attributes of national wildlife health programmes. Rev. Sci. Tech. Off. Int. Epiz. 2018, 37, 925–936. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Ehrlich, P.R.; Beattie, A.; Ceballos, G.; Crist, E.; Diamond, J.; Dirzo, R.; Ehrlich, A.H.; Harte, J.; Harte, M.E.; et al. Underestimating the Challenges of Avoiding a Ghastly Future. Front. Conserv. Sci. 2021, 1, 9. [Google Scholar] [CrossRef]
- Le Breton, T.D.; Zimmer, H.C.; Gallagher, R.V.; Cox, M.; Allen, S.; Auld, T.D. Using IUCN criteria to perform rapid assessments of at-risk taxa. Biodivers. Conserv. 2019, 28, 863–883. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, K.M.; Kline, R.J.; Rahman, S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Carøe, C.; Gopalakrishnan, S.; Vinner, L.; Mak, S.S.T.; Sinding, M.H.S.; Samaniego, J.A.; Wales, N.; Sicheritz-Pontén, T.; Gilbert, M.T.P. Single-tube library preparation for degraded DNA. Methods Ecol. Evol. 2017, 9, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Carroll, E.L.; Bruford, M.W.; DeWoody, J.A.; Leroy, G.; Strand, A.; Waits, L.; Wang, J. Genetic and genomic monitoring with minimally invasive sampling methods. Evol. Appl. 2018, 11, 1094–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozarth, C.A.; Alva-Campbell, Y.R.; Ralls, K.; Henry, T.R.; Smith, D.A.; Westphal, M.F.; Maldonado, J.E. An efficient noninvasive method for discriminating among faeces of sympatric North American canids. Conserv. Genet. Resour. 2010, 2, 173–175. [Google Scholar] [CrossRef]
- Wilbert, T.R.; Woollett, D.A.S.; Westphal, M.F.; Whitelaw, A.; Ralls, K.; Maldonado, J.E. Distribution, fine-scale subdivision, and population size of San Joaquin kit foxes in the Ciervo-Panoche Natural Area, California. Conserv. Genet. 2019, 20, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, S.; Maldonado, J.; Ortega, J.; Talarico, A.C.; Bidegaray-Batista, L.; Garcia, J.E.; Duarte, J.M.B. Identification of the endangered small red brocket deer (Mazama bororo) using noninvasive genetic techniques (Mammalia; Cervidae). Mol. Ecol. Resour. 2009, 9, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Grattarola, F.; Gonzalez, S.; Cosse, M. A novel primer set for mammal species identification from feces samples. Conserv. Genet. Resour. 2014, 7, 57–59. [Google Scholar] [CrossRef]
- Tian, J.; Bao, X.; Du, J.; Lu, Z.; Li, Y.; Fu, Z.; Liu, W. Sex identification based on the CHD gene from Gentoo penguin (Pygoscelis papua) fecal DNA samples. Conserv. Genet. Resour. 2021, 13, 249–253. [Google Scholar] [CrossRef]
- Matsui, A.; Rakotondraparany, F.; Hasegawa, M.; Horai, S. Determination of a complete lemur mitochondrial ge-nome from feces. Mammal Study 2007, 32, 7–16. [Google Scholar] [CrossRef]
- van der Valk, T.; Durazo, F.L.; Dalén, L.; Guschanski, K. Whole mitochondrial genome capture from faecal samples and museum-preserved specimens. Mol. Ecol. Resour. 2017, 17, e111–e121. [Google Scholar] [CrossRef] [Green Version]
- Moss, E.L.; Maghini, D.G.; Bhatt, A.S. Complete, closed bacterial genomes from microbiomes using nanopore sequencing. Nat. Biotechnol. 2020, 38, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Yiming, L.; Zhongwei, G.; Qisen, Y.; Yushan, W.; Niemelä, J. The implications of poaching for giant panda conservation. Biol. Conserv. 2003, 111, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Xu, M.; Wong, M.H.G.; Qiu, S.; Sheng, Q.; Li, X.; Song, Z. Climate change-induced decline in bamboo habitats and species diversity: Implications for giant panda conservation. Divers. Distrib. 2015, 21, 379–391. [Google Scholar] [CrossRef]
- Swaisgood, R.; Wang, D.; Wei, F. errata version of 2016 assessment. Ailuropoda melanoleuca. In IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural: Cambridge, UK, 2017; p. e.T712A121745669, Retrieved 15 January 2022. [Google Scholar]
- Ullah, Z.; Ullah, I.; Ullah, I.; Mahmood, S.; Iqbal, Z. Poaching of Asiatic black bear: Evidence from Siran and Kaghan valleys, Pakistan. Glob. Ecol. Conserv. 2020, 24, e01351. [Google Scholar] [CrossRef]
- Pires, S.F.; Moreto, W.D. The Illegal Wildlife Trade. Oxford Handbooks Online. 2016. Available online: https://academic.oup.com/edited-volume/41333/chapter/352362392. [CrossRef] [Green Version]
- Garshelis, D.; Steinmetz, R. Ursus thibetanus. In IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural: Cambridge, UK, 2020; p. e.T22824A166528664. [Google Scholar] [CrossRef]
- Pietersen, D.W.; Challender, D.W. Research needs for pangolins. In Pangolins: Science, Society and Conservation; Challender, D., Nash, H., Waterman, C., Eds.; Academic Press: London, UK, 2020; pp. 537–543. [Google Scholar] [CrossRef]
- Heighton, S.P.; Gaubert, P. A timely systematic review on pangolin research, commercialization, and popularization to identify knowledge gaps and produce conservation guidelines. Biol. Conserv. 2021, 256, 109042. [Google Scholar] [CrossRef]
- Challender, D.W.; Heinrich, S.; Shepherd, C.R.; Katsis, L.K. International trade and trafficking in pangolins, 1900–2019. In Pangolins; Academic Press: London, UK, 2020; pp. 259–276. [Google Scholar] [CrossRef]
- Winn, H.E.; Price, C.A.; Sorensen, P.W. The distributional biology of the right whale (Eubalaena glacialis) in the western North Atlantic. In Right Whales: Past and Present Status; IWC: Cambridge, UK, 1986; pp. 129–138. [Google Scholar]
- Reilly, S.B.; Bannister, J.L.; Best, P.B.; Brown, M.; Brownell, R.L., Jr.; Butterworth, D.S.; Clapham, P.J.; Cooke, J.; Donovan, G.P.; Urbán, J.; et al. Eubalaena japonica. IUCN Red List of Threatened Species. 2008. Version 2012.2. Available online: www.iucnredlist.org (accessed on 28 August 2022).
- Kraus, S.D.; Brown, M.W.; Caswell, H.; Clark, C.W.; Fujiwara, M.; Hamilton, P.K.; Kenney, R.D.; Knowlton, A.R.; Landry, S.; Mayo, C.A.; et al. North Atlantic Right Whales in Crisis. Science 2005, 309, 561–562. [Google Scholar] [CrossRef]
- Knowlton, A.R.; Brown, M.W. Running the gauntlet: Right whales and vessel strikes. In The Urban Whale: North Atlantic Right Whales at the Crossroads; Kraus, S.D., Rolland, R.M., Eds.; Harvard University Press: Cambridge, MA, USA, 2007; pp. 409–435. [Google Scholar]
- Sanders, J.G.; Beichman, A.C.; Roman, J.; Scott, J.J.; Emerson, D.; McCarthy, J.J.; Girguis, P.R. Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores. Nat. Commun. 2015, 6, 8285. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Mishra, S.; Zhao, J.; Tang, J.; Zeng, B.; Kong, F.; Ning, R.; Li, M.; Zhang, H.; Zeng, Y.; et al. Metagenomic Study Suggests That the Gut Microbiota of the Giant Panda (Ailuropoda melanoleuca) May Not Be Specialized for Fiber Fermentation. Front. Microbiol. 2018, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-E.; Jiang, H.-Y.; Li, L.-M.; Zhang, X.-J.; Li, G.-Y.; Li, H.-M.; Jin, X.-J.; Chen, J.-P. The Fecal Metagenomics of Malayan Pangolins Identifies an Extensive Adaptation to Myrmecophagy. Front. Microbiol. 2018, 9, 2793. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; Depamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 1–31. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernt, M.; Braband, A.; Schierwater, B.; Stadler, P.F. Genetic aspects of mitochondrial genome evolution. Mol. Phylogenetics Evol. 2013, 69, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conant, G.C.; Wolfe, K.H. GenomeVx: Simple web-based creation of editable circular chromosome maps. Bioinformatics 2008, 24, 861–862. [Google Scholar] [CrossRef] [Green Version]
- Jühling, F.; Pütz, J.; Florentz, C.; Stadler, P.F. Armless mitochondrial tRNAs in Enoplea (Nematoda). RNA Biol. 2012, 9, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Kerpedjiev, P.; Hammer, S.; Hofacker, I.L. Forna (force-directed RNA): Simple and effective online RNA secondary structure diagrams. Bioinformatics 2015, 31, 3377–3379. [Google Scholar] [CrossRef] [Green Version]
- Gruber, A.; Lorenz, R.; Bernhart, S.H.F.; Neuböck, R.; Hofacker, I.L. The Vienna RNA Websuite. Nucleic Acids Res. 2008, 36 (Suppl. S2), W70–W74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sbisà, E.; Tanzariello, F.; Reyes, A.; Pesole, G.; Saccone, C. Mammalian mitochondrial D-loop region structural analysis: Identification of new conserved sequences and their functional and evolutionary implications. Gene 1997, 205, 125–140. [Google Scholar] [CrossRef]
- Hwang, D.-S.; Ki, J.-S.; Jeong, D.-H.; Kim, B.-H.; Lee, B.-K.; Han, S.-H.; Lee, J.-S. A comprehensive analysis of three Asiatic black bear mitochondrial genomes (subspecies ussuricus, formosanus and mupinensis), with emphasis on the complete mtDNA sequence of Ursus thibetanus ussuricus(Ursidae). DNA Seq. 2008, 19, 418–429. [Google Scholar] [CrossRef]
- Hari, R.; Paterson, I.C.; Choo, S.W. A new complete mitogenome of the critically endangered Chinese pangolin Manis pentadactyla. Conserv. Genet. Resour. 2016, 8, 423–426. [Google Scholar] [CrossRef]
- du Toit, Z.; du Plessis, M.; Dalton, D.L.; Jansen, R.; Grobler, J.P.; Kotzé, A. Mitochondrial genomes of African pangolins and insights into evolutionary patterns and phylogeny of the family Manidae. BMC Genom. 2017, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Allwood, J.S.; Scheible, M.K.; Faith, S.A. Demonstration of a basic mitochondrial enrichment method to produce the complete mitochondrial genome sequence of the endangered North Atlantic right whale (Eubalaena glacialis). Conserv. Genet. Resour. 2017, 10, 483–486. [Google Scholar] [CrossRef]
- Sasaki, T.; Nikaido, M.; Hamilton, H.; Goto, M.; Kato, H.; Kanda, N.; Pastene, L.A.; Cao, Y.; Fordyce, E.; Hasegawa, M.; et al. Mitochondrial Phylogenetics and Evolution of Mysticete Whales. Syst. Biol. 2005, 54, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Arnason, U.; Gullberg, A.; Janke, A. Mitogenomic analyses provide new insights into cetacean origin and evolution. Gene 2004, 333, 27–34. [Google Scholar] [CrossRef]
- Xiufeng, X.; Árnason, Ú. The complete mitochondrial DNA sequence of the horse, Equus caballus: Extensive heteroplasmy of the control region. Gene 1994, 148, 357–362. [Google Scholar] [CrossRef]
- Kumar, A.; Yadav, P.; Usmani, A.; Hussian, S.A.; Gupta, S.K. Complete mitogenome of Ganges river dolphin, Platanista gangetica gangetica and its phylogenetic relationship with other cetaceans. Mol. Biol. Rep. 2021, 48, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Zeng, B.; Meng, X.; Yue, B.; Zhang, Z.; Zou, F. The complete mitochondrial genome and phylogenetic analysis of the giant panda (Ailuropoda melanoleuca). Gene 2007, 397, 76–83. [Google Scholar] [CrossRef]
- Beckenbach, A.T. Numts and mitochondrial pseudogenes. Myrmecol. News 2009, 12, 217–218. [Google Scholar]
- Bit, A.; Thakur, M.; Singh, S.K.; Joshi, B.D.; Singh, V.K.; Sharma, L.K.; Tripathy, B.; Chandra, K. Assembling mitogenome of Himalayan Black Bear (U. t. laniger) from low depth reads and its application in drawing phylogenetic inferences. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Montelli, S.; Peruffo, A.; Patarnello, T.; Cozzi, B.; Negrisolo, E. Back to Water: Signature of Adaptive Evolution in Cetacean Mitochondrial tRNAs. PLoS ONE 2016, 11, e0158129. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, S.R.; Sussman, J.; Warrant, R.; Kim, S.-H. Crystal structure of yeast phenylalanine transfer RNA: II. Structural features and functional implications. J. Mol. Biol. 1978, 123, 631–660. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Huang, C.-C.; Tseng, Y.-W.; Suwal, T.L.; Chi, M.-J.; Jang-Liaw, N.-H.; Hung, K.-H. Complete mitochondrial genome of Manis pentadactyla pentadactyla (Mammalia: Pholidota), an endemic subspecies of Chinese pangolin: Mitogenome characterisation and phylogenetic implications. Biodivers. Data J. 2021, 9, e77961. [Google Scholar] [CrossRef]
- Hou, W.-R.; Chen, Y.; Wu, X.; Hu, J.-C.; Peng, Z.-S.; Yang, J.; Tang, Z.-X.; Zhou, C.-Q.; Li, Y.-M.; Yang, S.-K.; et al. A complete mitochondrial genome sequence of Asian black bear Sichuan subspecies (Ursus thibetanus mupinensis). Int. J. Biol. Sci. 2007, 3, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldick, R.C.; Kraus, S.; Brown, M.; White, B.N. Evaluating the effects of historic bottleneck events: An assessment of microsatellite variability in the endangered, North Atlantic right whale. Mol. Ecol. 2002, 11, 2241–2249. [Google Scholar] [CrossRef] [PubMed]
- Hoelzel, A.R.; Hancock, J.M.; A Dover, G. Evolution of the cetacean mitochondrial D-loop region. Mol. Biol. Evol. 1991, 8, 475–493. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, A.; Hugot, J.-P.; van Vuuren, B.J. Comparison of mitochondrial genome sequences of pangolins (Mammalia, Pholidota). Comptes Rendus Biol. 2015, 338, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Gemmell, N.J.; Western, P.; Watson, J.M.; Graves, J. Evolution of the mammalian mitochondrial control region--comparisons of control region sequences between monotreme and therian mammals. Mol. Biol. Evol. 1996, 13, 798–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas-Toro, I.; Ortega, J.; Baeza, J.A. The complete mitochondrial genome of the Honduran white bat Ectophylla alba (Allen 1982) (Chiroptera: Phyllostomidae). Gene 2021, 802, 145868. [Google Scholar] [CrossRef]
- Ennis, C.C.; Ortega, J.; Baeza, J.A. First genomic resource for an endangered neotropical mega-herbivore: The complete mitochondrial genome of the forest-dweller (Baird’s) tapir (Tapirus bairdii). PeerJ 2022, 10, e13440. [Google Scholar] [CrossRef]


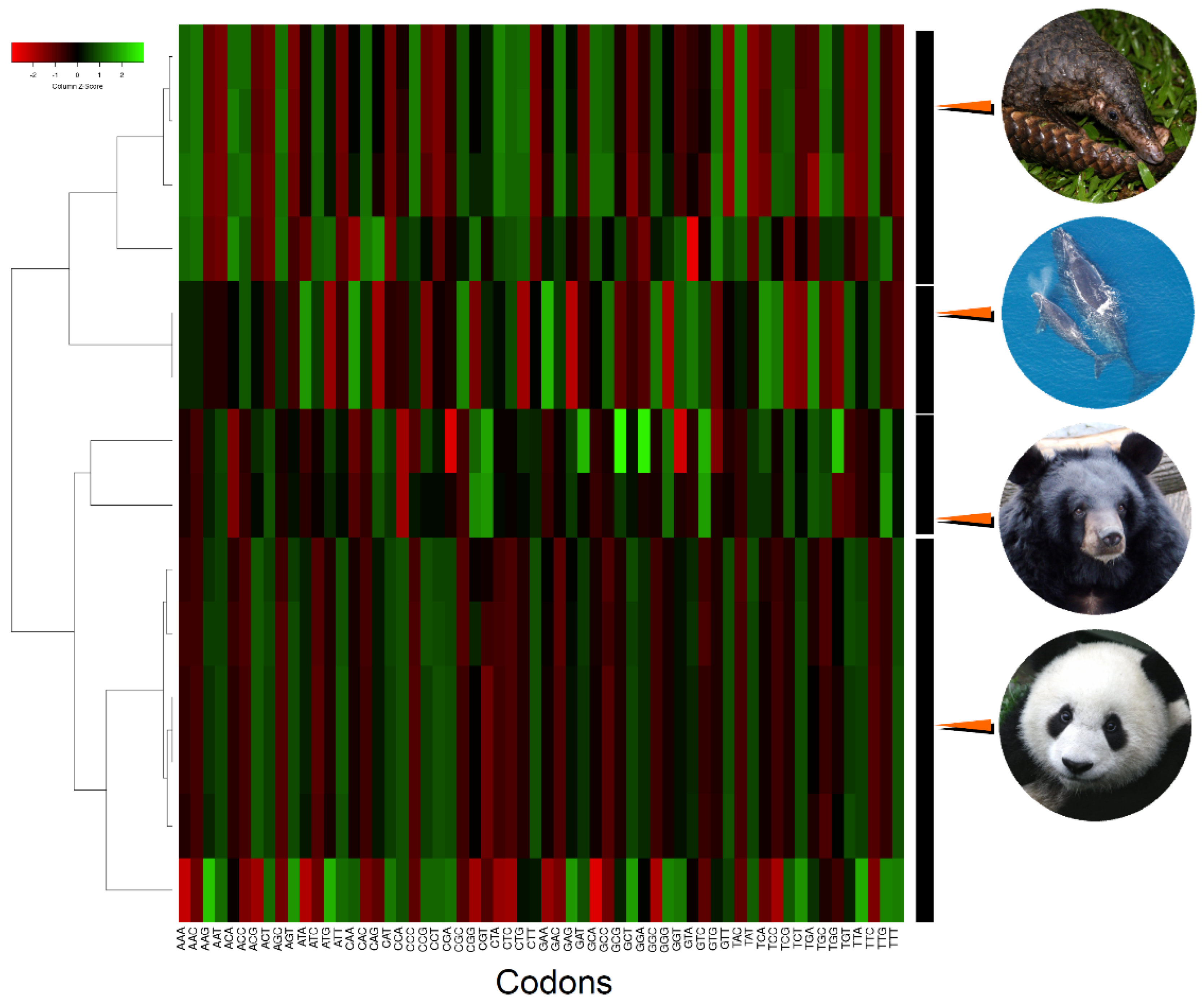
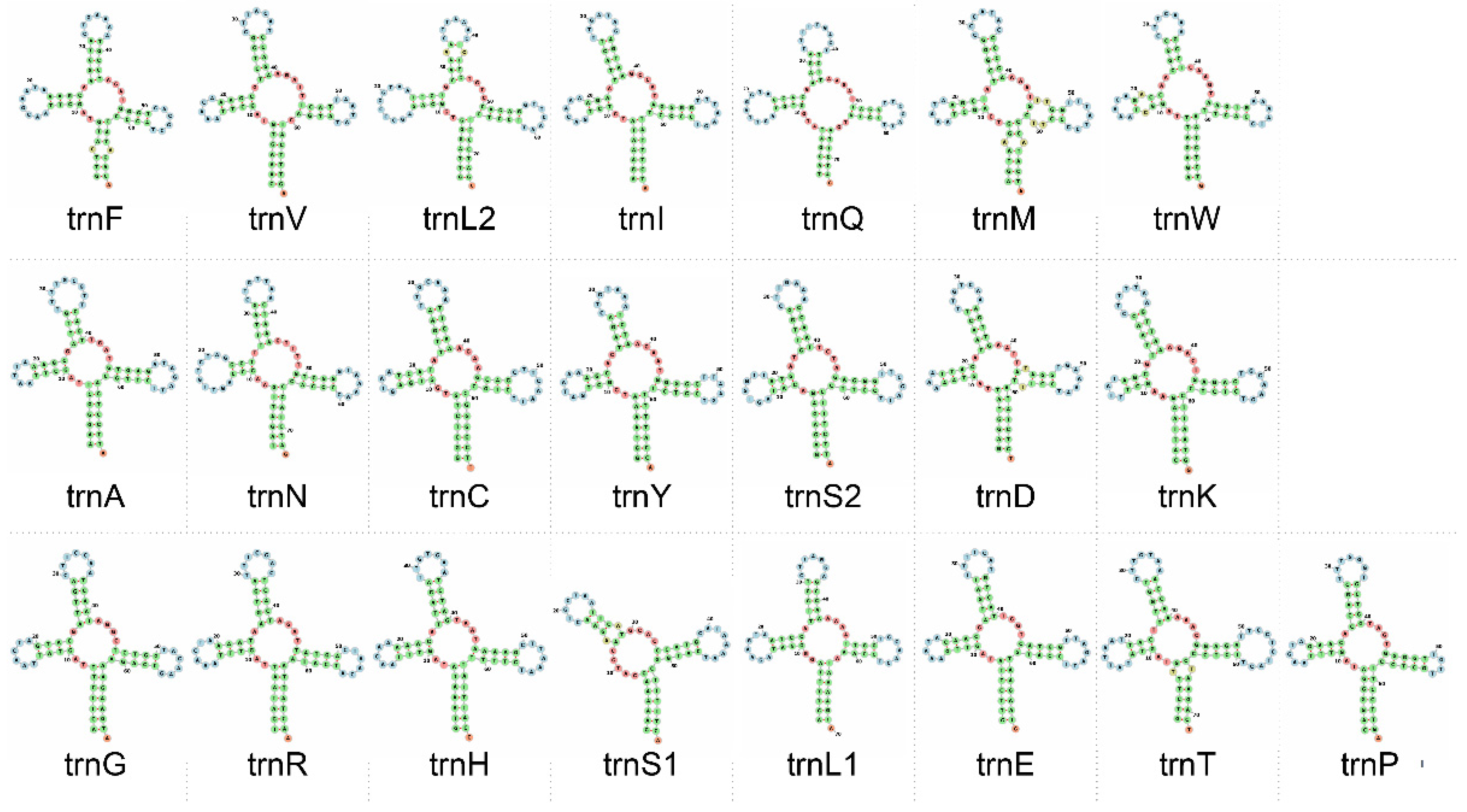



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | NCBI SRA | Read N | Assembly | Coverage | Length (bp) | A |
|---|---|---|---|---|---|---|
| Ursus thibetanus | SRR6109212 | 10,906,235 | Circular | 122x | 16,634 | 1 |
| Ursus thibetanus | SRR6109213 | 20,539,115 | Partial (1) | 717x | 4.290 * | – |
| Ursus thibetanus | SRR6109214 | 8,678,339 | Circular | 79x | 16,725 | 2 |
| Ursus thibetanus | SRR6109215 | 8,052,349 | None | – | – | – |
| Ailuropoda melanoleuca | SRR6109207 | 12,404,708 | Circular | 40x | 16,615 | 3 |
| Ailuropoda melanoleuca | SRR6109208 | 11,477,287 | Circular | 246x | 16,786 | 4 |
| Ailuropoda melanoleuca | SRR6109209 | 11,245,869 | Circular | 34x | 16,677 | 5 |
| Ailuropoda melanoleuca | SRR6109210 | 11,944,071 | Circular | 224x | 16,852 | 6 |
| Ailuropoda melanoleuca | SRR6109211 | 12,489,755 | Circular | 56x | 16,617 | 7 |
| Ailuropoda melanoleuca | SRR6109216 | 12,611,226 | Circular | 127x | 16,767 | 8 |
| Manis javanica | SRR7477311 | 23,833,983 | Circular | 75x | 16,576 | 9 |
| Manis javanica | SRR7498027 | 23,619,748 | Circular | 183x | 16,576 | 10 |
| Manis javanica | SRR7507293 | 23,879,794 | Circular | 480x | 16,576 | 11 |
| Manis javanica | SRR7524043 | 24,216,055 | Circular | 294x | 16,576 | 12 |
| Eubalena glacialis | ERR4056911 | 7,013,164 | Partial (7) | 6x | 15,573 * | – |
| Eubalena glacialis | ERR4056912 | 9,384,989 | Circular | 43x | 16,387 | 13 |
| Eubalena glacialis | ERR4056913 | 10,423,814 | Partial (4) | 11x | 16,291 * | – |
| Eubalena glacialis | ERR4056914 | 7,194,209 | Circular | 12x | 16,387 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baeza, J.A.; Barata, R.; Rajapakse, D.; Penaloza, J.; Harrison, P.; Haberski, A. Mitochondrial Genomes Assembled from Non-Invasive eDNA Metagenomic Scat Samples in Critically Endangered Mammals. Genes 2023, 14, 657. https://doi.org/10.3390/genes14030657
Baeza JA, Barata R, Rajapakse D, Penaloza J, Harrison P, Haberski A. Mitochondrial Genomes Assembled from Non-Invasive eDNA Metagenomic Scat Samples in Critically Endangered Mammals. Genes. 2023; 14(3):657. https://doi.org/10.3390/genes14030657
Chicago/Turabian StyleBaeza, J. Antonio, Ryan Barata, Dilani Rajapakse, Jayra Penaloza, Preston Harrison, and Adam Haberski. 2023. "Mitochondrial Genomes Assembled from Non-Invasive eDNA Metagenomic Scat Samples in Critically Endangered Mammals" Genes 14, no. 3: 657. https://doi.org/10.3390/genes14030657