Abstract
Elucidating the genetic basis of starch pasting and gelatinization properties is crucial for enhancing the quality of maize and its utility as feed and industrial raw material. In maize, ZmSBE genes encode important starch branching enzymes in the starch biosynthesis pathway. In this study, we re-sequenced the genomic sequences of ZmSBEI, ZmSBEIIa, ZmSBEIIb, and ZmSBEIII in three lines called 335 inbred lines, 68 landrace lines, and 32 teosinte lines. Analyses of nucleotide polymorphisms and haplotype diversity revealed differences in the selection patterns of ZmSBEI, ZmSBEIIa, ZmSBEIIb, and ZmSBEIII during maize domestication and improvement. A marker–trait association analysis of inbred lines detected 22 significant loci, including 18 SNPs and 4 indels significantly associated with three maize starch physicochemical properties. The allele frequencies of two variants (SNP17249C and SNP5055G) were examined in three lines. The frequency of SNP17249C in ZmSBEIIb was highest in teosinte lines, followed by landrace lines, and inbred lines, whereas there were no significant differences in the frequency of SNP5055G in ZmSBEIII among the three lines. These results suggest that ZmSBE genes play an important role in the phenotypic variations in the starch physicochemical properties in maize. The genetic variants detected in this study may be used to develop functional markers for improving maize starch quality.
1. Introduction
Maize starch is one of the highest-quality starches and accounts for 80% of the global starch yield [1]. Moreover, it is used as food and feed as well as an industrial raw material (e.g., for biodiesel production) [2]. Therefore, identifying the genes or natural variations associated with starch quality and quantity in maize kernels may help to increase the nutritional value of maize through the breeding of high-quality lines.
Starch is an important storage polysaccharide comprising amylose and amylopectin, which are structurally diverse polymers. The content ratio of these two components influences the function, quality, and utility of starch. Changing the structure of starch may enhance how it can be used. The basic methods for synthesizing starch in plants include synthesizing soluble precursors, followed by coordinated reactions: firstly, the glucan chain is connected to α-(1→4) and elongating, and then, branching, and debranching at α-(1→6) positions (i.e., hydrolysis of specific branch linkages). AGPase synthesizes the nucleotide diphosphate sugar precursor ADP-Glc through the catalysis of ATP and Glc1P. The α-(1→4)-linked glucan chains are formed by reactions catalyzed by ADP-Glc-dependent transferases, which are called starch synthases (SSs), whereas α-(1→6)-linked branch points are introduced via reactions mediated by starch branching enzymes (SBEs) and debranching is catalyzed by debranching enzymes (DBEs) [3]. Thus, the starch synthesis pathway requires diverse enzymes (e.g., GBSSI, SSs, SBEs, and DBEs). Of these enzymes, GBSSI (granule-bound starch synthase I) synthesizes amylose in the cereal endosperm, while SSs, SBEs, and DBEs coordinately synthesize amylopectin. Two non-enzymatic proteins (PTST1 and PTST2) occur in the starch synthesis pathway. Notably, SBEs are the key enzymes catalyzing the synthesis of amylopectin, which is the main component of plant starch. Therefore, the nature and activity of SBEs are the decisive factors affecting the fine structure of amylopectin [4].
As the only enzymes that catalyze the formation of amylopectin branch linkages, which are called starch branching enzymes, they modify glucan to produce branches connected by α-1,6-glycosidic bonds [5]. On the basis of amino acid sequence relationships, SBEs have been divided into the following two classes: Class I (B family) and Class II (A family). Gene expression analyses of a range of species revealed that during development, SBEI genes are expressed later than SBEII genes [6]. In cereals, there are three types of SBE-encoding genes, namely SBEI, SBEII, and SBEIII. Earlier research indicated SBEII genes can be subdivided into SBEIIa and SBEIIb genes, which differ in terms of their expression kinetic characteristics and tissue expression patterns. Specifically, SBEIIb is expressed exclusively in the endosperm tissue, whereas SBEIIa appears to be expressed ubiquitously [7]. Moreover, SBEII is important for amylopectin synthesis in the cereal endosperm. However, there are considerable variations in the relative proportions of SBEIIa and SBEIIb activities in the endosperm among diverse cereals [7,8]. In the developing wheat endosperm, SBEIIb is expressed at much lower levels than SBEIIa [9], whereas in the maize endosperm, the SBEIIb expression level is approximately 50-times greater than SBEIIa [8]. Furthermore, SBEIII, which differs from SBEI and SBEII, has been detected in many higher plants, including rice, maize, and wheat, although only TaSBEIII contributes to the production of both A and B granules in wheat grains [10,11].
Maize was domesticated from teosinte approximately 9000 years ago. Kernel starch was one of the targets of artificial selection during maize domestication [12]. The starch biosynthesis-related functions of individual SBE isozymes have been examined, but SBE gene sequence polymorphisms and natural variations in maize remain unclear. It is also unknown whether SBE genes were selected during maize domestication and improvement. In this study, we re-sequenced four ZmSBE genes in 335 inbred lines, 68 landrace lines, and 32 teosinte lines. The objectives of this research were as follows: firstly, examine the diversity in the ZmSBE sequences among three lines; secondly, identify natural variations in candidate genes associated with the kernel starch content and starch pasting and gelatinization properties; finally, examine significant associations to clarify their involvement in maize domestication and improvement.
2. Materials and Methods
2.1. Plant Materials
A total of 335 inbred lines, 68 landrace lines, and 32 teosinte lines were selected in this study (Table S1) [13]. The inbred lines were planted in the experimental field at Sanya (N 18°23′, E 109°44′), Hainan, China, in 2017 and 2018. The plants were grown by the randomized complete block design (RCBD) with two replicates. Each inbred line was grown in a single row, with a length of 3 m and a distance between each row of 0.5 m.
2.2. DNA Isolation and ZmSBEs Resequencing
The genomic DNA of each line was extracted from three mixed leaves (approximately 15 days after the emergence of maize) using the CTAB (cetyl trimethyl ammonium bromide) method [14,15]. The sequences (B73_V3 reference genome) of the ZmSBEI (GRMZM2G088753), ZmSBEIIa (GRMZM2G073054), ZmSBEIIb (GRMZM2G032628), and ZmSBEIII (GRMZM2G005298) gene in all tested lines were sequenced using the target sequence capture sequencing technology on the NimbleGen platform [16,17] by BGI Life Tech Co., Ltd (Guangdong, China). The reference sequences captured by the target sequence were the ZmSBEs gene sequences in the background of the B73 inbred line (Table S2).
2.3. Determination of Maize Kernel Starch Content and Starch Pasting and Gelatinization Properties
At the maturity stage, the ears for each inbred line were harvested, and approximately 50–60 grains were stripped from the middle of the corn ear and immediately frozen in liquid nitrogen, and preserved at −75 °C for analysis and determination of starch. After drying and weighing, the granules were crushed and passed through a sieve (100 mesh, d = 0.149 mm) to determine the starch content. The crude starch content of corn (KSC) kernel was estimated using a near-infrared analyzer (FOSS, Beijing, China).
A rapid visco analyser (RVA) (Model No. RVA-3D, Newport Scientific, Sydney, Australia) was used to determine the pasting properties. A total of 3 g of starch taken from each inbred line was dispersed into 25 mL of distilled water in the viscometer test canister. Then, the test was performed with RVA. The set speed was 160 rpm/min. Viscosity values were recorded in centipoise (cp). The pasting properties were peak viscosity (PV), trough viscosity (TV), breakdown viscosity (BD), final viscosity (FV), setback viscosity (SB), peak time (PT), and pasting temperature (Ptemp).
The gelatinization properties of maize starches were analyzed using a differential scanning calorimeter DSC 200F3 Maia (Netzsch, Germany). Firstly, 5 mg starch samples (dried starch basis) were weighed in a small crucible. Secondly, 10 mL of distilled water was added to mix it. Finally, it was sealed and stored at 4 °C. The samples were used the next day to perform DSC determination. The gelatinization properties were onset temperature (To), peak temperature (Tp), conclusion temperature (Tc), and the enthalpy of gelatinization (ΔHgel).
2.4. Analysis of Sequence Data
Multiple sequence alignments of the maize gene ZmSBEs were performed using MAFFT software [18]. MEGA7 software was used for manual proofreading. The analysis of sequences was performed using the software DNASP 5.0 [15,17]. The symbols π and θ denoted the nucleotide polymorphism of the gene, whereby π is the average number of nucleotide differences per site between any two DNA sequences, and θ was derived from the total number of segregating sites and corrected for sampling size. Tajima’s D [19] and Fu and Li’s D* and F* statistical tests [20] were used to test the neutral evolution. The levels of linkage disequilibrium (LD) between two polymorphic sites in a coding region were calculated using TASSEL 5.0 [21], and the LD level was expressed by the linkage disequilibrium parameter r2.
A total of 163,931 SNPs were obtained by genotyping using a sequencing strategy [13]. Among them, we removed the markers with a deletion rate of more than 20% and a minor allele frequency (MAF) of less than 1%. Principal component analysis (PCA) and kinship were calculated using TESSEL 5.0, and the top five PCs (Figure S1) were used to create a population structure matrix in 335 inbred lines. Gene-based variants with MAF ≥ 0.05 were identified by TASSEL 5.0. Association analysis was performed using mixed linear models (MLM) + principal components analysis (PCA) + Kinship in TASSEL 5.0. The p value threshold to control the genome-wide type 1 error rate was 2.1 × 10−3 (1/n, where n is 479, the largest marker number of 4 SBEs).
3. Results
3.1. ZmSBE Sequence Polymorphisms
The analysis of four SBE isozyme-encoding genes revealed ZmSBEI, ZmSBEIIa, ZmSBEIIb, and ZmSBEIII differ substantially in terms of length (i.e., 5694, 10,473, 17,049, and 2940 bp, respectively) (Table 1). Both ZmSBEI and ZmSBEIIb were detected on chromosome 5, whereas ZmSBEIIa and ZmSBEIII were detected on chromosomes 2 and 8, respectively. The encoded amino acid sequences consisted of similar domains, with all four proteins containing α-amylase-C, CMB-48, and α-amylase (Figure 1). To identify the ZmSBE sequence polymorphisms, such as SNPs and indels, the upstream genomic region (approximately 2000 bp), coding region, and downstream genomic region (approximately 500 bp) in three tested lines were sequenced. For ZmSBEI, 1102 polymorphisms were found, consisting of 916 SNPs and 186 indels. On average, the SNPs and indels occurred every 9.9 and 50 bp, respectively. The average length of each indel was 5.4 bp. A total of 1519 sequence variations were identified in ZmSBEIIa, including 1302 SNPs and 217 indels. On average, the SNPs and indels were detected every 9.7 and 58.8 bp, respectively. The average length of each indel was 4.9 bp. Of the 1616 sequence variations detected in ZmSBEIIb, 1353 were SNPs and 263 were indels. On average, the SNPs and indels occurred every 14.5 and 76.9 bp, respectively. The average length of each indel was 3.9 bp. In contrast, ZmSBEIII included 1044 polymorphisms, including 903 SNPs and 141 indels. On average, the SNPs and indels were detected every 7.6 and 47.6 bp, respectively. The average length of each indel was 4.7 bp (Table 1).

Table 1.
Overview of parameters of the nucleotide polymorphisms analysis of ZmSBEs.
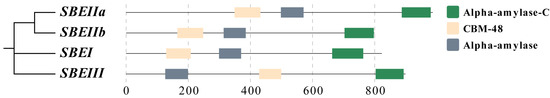
Figure 1.
Protein domain of ZmSBEs. Each of the four gene domains contains α-amylase-C, CBM-48, and α-amylase, which are distinguished by different colors.
3.2. Analysis of Nucleotide Diversity and Selection of ZmSBE Genes in Teosinte, Landraces, and Inbred Lines
To survey the variety among the four ZmSBE genes (ZmSBEI, ZmSBEIIa, ZmSBEIIb, and ZmSBEIII) in all tested lines, sequence variations were analyzed. Compared with the genes in the teosinte lines, those in the landrace lines and inbred lines were more conserved (CT < CL < CI), less diverse (πT > πL > πI), and had fewer nucleotide sequence polymorphisms (θT > θL > θI) (Table 2). The π value was lower for ZmSBEIIa than for the other three genes, implying that ZmSBEIIa had the least diverse nucleotide sequence (Table 2 and Figure 2). To further assess whether ZmSBE genes were chosen during maize evolution, neutrality tests (Tajima’s D along with Fu and Li’s D* and F*) were performed to analyze the tested sequences. Tajima’s D and Fu and Li’s F* values for ZmSBEI and ZmSBEIIb were significantly less than 0 in all test lines, showing that these two genes were chosen during evolution. For both ZmSBEIIa and ZmSBEIII, Fu and Li’s D* and F* values were significantly less than 0, but Tajima’s D value was not. Hence, these two genes may have undergone neutral evolution (Table 2).
Table 2.
Summary of nucleotide polymorphisms and neutrality test of four SBE genes.
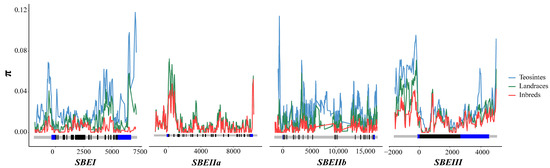
Figure 2.
The nucleotide diversity (π) of teosinte, landraces, and inbred lines. π is calculated using the method of sliding windows of 100 bp with a step of 25 bp. A schematic diagram of the ZmSBEs gene structure, which contains upstream sequence and introns (light gray), the coding region (black), and 5’ UTR and 3’ UTR (blue) are shown. The position of the start codon (ATG) is labeled as “0”, and the negative value indicates the upstream sequence of the gene.
Linkage disequilibrium (LD) decay varied among the analyzed genes. For ZmSBEI, LD was less extensive in the teosinte lines than in the landrace lines and inbred lines, implying that this gene may have been chosen during maize domestication. For ZmSBEIIa, the extent of LD in the teosinte lines was similar to that in the landrace lines, but lower than in the inbred lines. Accordingly, ZmSBEIIa may have been chosen during maize improvement. In contrast, ZmSBEIIb may have been chosen during both maize domestication and improvement, whereas the likelihood that ZmSBEIII was chosen during maize domestication and improvement was relatively low (Figure 3).
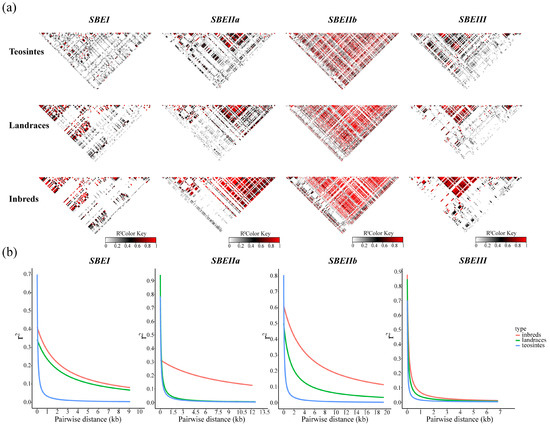
Figure 3.
Linkage disequilibrium and recombination events. (a) Linkage disequilibrium analysis of ZmSBEs for three lines. It indicates LD between pairs of ZmSBEs sequence polymorphic sites. The R2 values are indicated using the color bar. (b) LD decay of ZmSBEs for three lines determined by r2. Three lines are inbred lines (red), landrace lines (green), and teosinte lines (blue).
3.3. Analysis of the Association between Phenotypes and ZmSBEs
A total of 12 maize starch physicochemical properties were analyzed in 335 inbred lines (Table S3). The phenotypic fold of variation and coefficient of variation had a wide range of parameters. These results reflected the considerable differences in the phenotypes of the inbred lines (Table S1). To investigate whether the natural variations in ZmSBE sequences were associated with starch physicochemical properties, an association analysis was performed on the basis of the variants with a minor allele frequency ≥ 0.05. Twenty-two significant variants of ZmSBEIIa, ZmSBEIIb, and ZmSBEIII were associated with three traits (Figure 4 and Table 3). Five significant loci in ZmSBEIIa were associated with ΔHgel and SB, and the phenotypic variation explained ranged from 2.99% to 4.28%. The variation in ΔHgel was mainly explained by indel 15969 in the intron region. Four significant loci in ZmSBEIIb were associated with SB, and the phenotypic variation explained ranged from 2.83% to 3.95%. Thirteen significant loci associated with ΔHgel and KSC were detected in ZmSBEIII, and the phenotypic variation explained ranged from 2.82% to 4.88%. The phenotypic variation was mainly explained by indel 830 in the upstream region. We also observed that individual traits were significantly associated with different genes. For example, ΔHgel and SB were controlled by ZmSBEIIa and ZmSBEIII (Table 3).
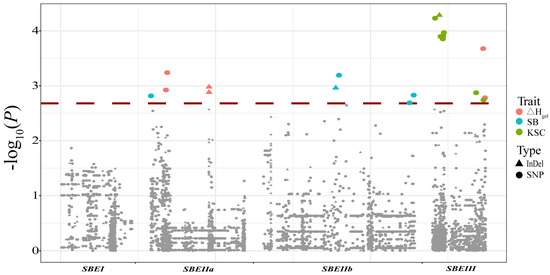
Figure 4.
Association analysis between ZmSBEs and starch quality-related traits. Indels are marked by triangles and SNPs are signed by dots. Trait abbreviations: ΔHgel: the enthalpy of gelatinization; SB: setback viscosity; KSC: crude starch content.
Table 3.
Results of candidate gene-based association analysis of SBE genes.
The four loci in ZmSBEIIa significantly associated with ΔHgel (403, 568, 5967, and 5969 bp) consisted of two SNPs and two indels. These loci were in intronic regions. The analysis of these variants detected strong LD for SNP403 and SNP568. The inbred lines were divided into five haplotypes according to the significant variants. The ANOVA results indicated that the differences in ΔHgel among the five haplotypes were significant (p = 1.1 × 10−3). We also analyzed the frequency of indel 5969 (i.e., the most significant variant) in teosinte, landraces, and inbred lines. The results indicated that the frequency of indel 5969C was lower in the teosinte lines (12.9%) than in the landrace lines (23.2%), and inbred lines (35.8%). Accordingly, indel 5969 may have been chosen during maize domestication and improvement (Figure 5).
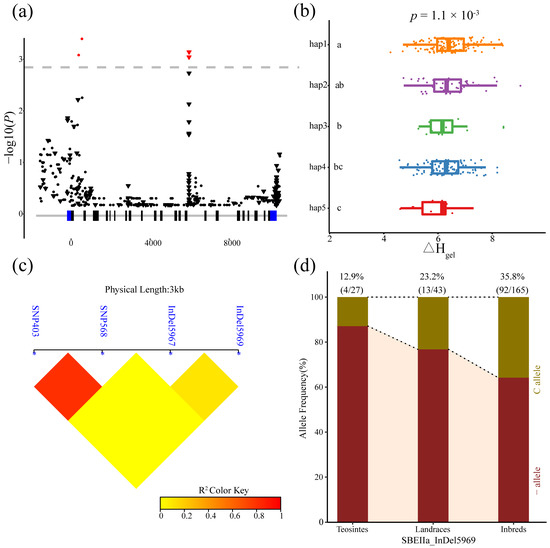
Figure 5.
Natural variability in ZmSBEIIa was significantly correlated with enthalpy (ΔHgel). (a) The Manhattan Plot of the association of ZmSBEIIa and ΔHgel. A diagrammatic sketch of the ZmSBEIIa gene structure is shown. (b) Comparisons of ΔHgel between haplotypes carrying different alleles in inbred lines. (c) Linkage disequilibrium (LD) analysis of four significant variants associated with ΔHgel. (d) The allele frequency of indel 5969 in three tested lines.
Four variants of ZmSBEIIb were associated with SB, including one indel and three SNPs (7723, 8152, 17,249, and 17,786 bp). The LD analysis showed that SNP17249 and SNP17786 are completely linked; the LD value (r2) of the remaining loci was approximately 0.8. Five major haplotypes were identified for the four loci across the 335 inbred lines. The differences in SB among these haplotypes were significant according to ANOVA (p = 1.1 × 10−6). The frequency of SNP 17249C was higher in the teosinte lines (87.5%) than in the landrace lines (66.2%), and inbred lines (46.3%), which indicated that SNP 17249C may have been gradually selected during maize domestication and improvement (Figure 6).
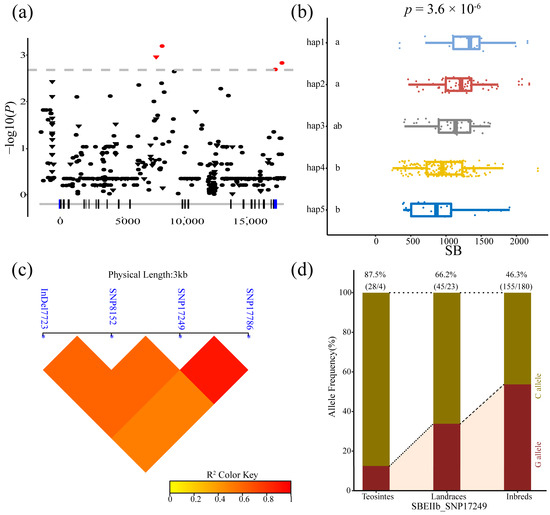
Figure 6.
Natural variations in ZmSBEIIb were significantly correlated with setback viscosity (SB). (a) The Manhattan Plot of the association of ZmSBEIIb and SB. A diagrammatic sketch of the ZmSBEIIb gene structure is shown. (b) Comparisons of SB between haplotypes carrying different alleles in inbred lines. (c) Linkage disequilibrium (LD) analysis of four significant variants associated with SB. (d) The allele frequency of SNP 17249 in three tested lines.
Two SNPs in ZmSBEIII (4817 and 5055 bp) were significantly associated with ΔHgel. Strong LD was observed between these two loci. In addition, significant differences in ΔHgel were detected among the three haplotypes for these two loci (p = 5.2 × 10−9). The frequency of SNP5055G did not differ significantly among the three populations, indicating that SNP5055 is not exposed to obvious selection during maize domestication and improvement (Figure 7).
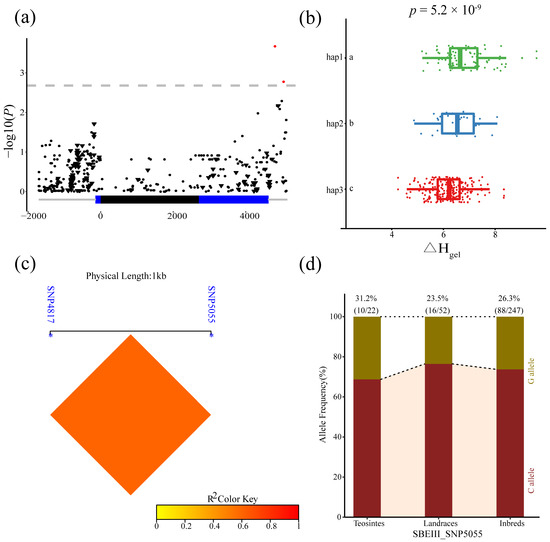
Figure 7.
Natural variations in ZmSBEIII were significantly correlated with enthalpy (ΔHgel). (a) The Manhattan Plot of the association of ZmSBEIII and ΔHgel. A diagrammatic sketch of the ZmSBEIII gene structure is shown. (b) Comparisons of SB between haplotypes carrying different alleles in inbred lines. (c) Linkage disequilibrium (LD) analysis of four significant variants associated with ΔHgel. (d) The allele frequency of SNP 5055 in three tested lines.
4. Discussion
Significant phenotypic differences and nucleotide polymorphisms are important for genetic mapping through linkage or association analyses [22,23]. Maize is a typical outcrossing crop, which has broad morphological variations, genetic diversity, and a high effective recombination rate [24,25]. The plant breeders can create novel plant gene combinations via abundant genetic variations and choose the variety of crops appropriate for diverse agricultural systems [22,23]. Analyzing the genetic diversity to screen for functional genes is critical for clarifying the genetic basis of crop phenotypic variations, thereby generating relevant information for crop improvement [23,26]. In this study, we revealed the polymorphisms in maize ZmSBE genes, which encode important enzymes in the starch biosynthesis pathway, in 335 inbred lines, 68 landrace lines, and 32 teosinte lines. The average nucleotide diversities (π) of these four genes were higher than previously reported in the whole genome [27]. The inbred lines had 52.0–82.8% less diversity than teosintes, and the selected gene ZmMADS69, which functions as a flowering activator, only retained 18.4% of the nucleotide diversity of teosinte [28]. In rice and soybean, cultivated varieties retained 70% and 36% of the nucleotide diversity of wild varieties [29,30]. Among these genes, ZmSBEIIb had the most variant sites and the highest variant frequency. Candidate gene association mapping on the basis of LD is a powerful method for identifying elite alleles for target traits [25]. This approach has been used to analyze several genes to detect variants associated with kernel composition [22,31,32,33]. In a previous study, markers were developed for the most favorable alleles of crtRB1, which is associated with the maize kernel β carotene concentration, for the provitamin A biofortification of crops [32]. In the current study, 22 significant variants of ZmSBEIIa, ZmSBEIIb, and ZmSBEIII were related to KSC, ΔHgel, and SB. The elite variations and the best haplotypes of ZmSBEIIa, ZmSBEIIb, and ZmSBEIII were identified, which may be used to generate inexpensive markers useful for enhancing the starch quality and quantity by molecular breeding in maize.
Compared with teosinte, cultivated maize has undergone considerable phenotypic changes (e.g., plant, ear, and seed morphologies) [34]. It is estimated that 2–4% of the genes were chosen during maize domestication and improvement [35]. There are also differences in kernel composition between teosinte and modern maize varieties. Starch is the main component of cereal seeds and contributes substantially to grain yield. The kernel starch content has been altered via strong selection during domestication and plant breeding [12]. The maize kernel starch content of inbred lines is reportedly 71.7%, which is higher than the corresponding content in teosinte kernels (52.92%) [12]. Previous research revealed that starch metabolism is the primary pathway in maize and rice that underwent convergent selection [36]. A total of 11 orthologous gene pairs in the starch metabolic pathway are associated with convergent selection. The ZmSBEI gene was chosen in both rice and maize [36]. The four ZmSBE genes in our study have different selection patterns during maize domestication and improvement. Specifically, ZmSBEI may have been chosen during maize domestication, while ZmSBEIIa may have been chosen during maize improvement. In contrast, it is possible that ZmSBEIIb was chosen during both maize domestication and improvement, whereas there was likely minimal selection of ZmSBEIII during maize domestication and improvement. Differences in the evolution of z2 genes, which influence the maize seed zein content, have been reported. The results revealed that teosinte and landrace lines may be crucial sources of genetic variation relevant for maize improvement [37].
In conclusion, maize ZmSBE genes were re-sequenced in 335 inbred lines, 68 landrace lines, and 32 teosinte lines. Analyses of nucleotide polymorphisms and haplotype diversity revealed differences in the selection patterns of ZmSBEI, ZmSBEIIa, ZmSBEIIb, and ZmSBEIII during maize domestication and improvement. A total of 22 significant variants of ZmSBEIIa, ZmSBEIIb, and ZmSBEIII were identified as associated with KSC, ΔHgel, and SB. The effect and selection of elite variations of ZmSBEIIa, ZmSBEIIb, and ZmSBEIII were identified. These results suggest that the genetic diversity of ZmSBE genes plays an important role in the variations of the starch physicochemical properties in maize. These variants may be applicable for increasing the maize kernel starch quality and quantity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14051068/s1, Table S1: The list of 335 inbred lines, 68 landraces, and 32 teosinte lines used in this study; Table S2: The sequences of ZmSBEs; Table S3: Statistical analysis of 12 phenotypic traits for tested inbred lines; Figure S1: The effect of each principal component; Figure S2: Natural variability in ZmSBEIIa were significantly correlated with setback viscosity; Figure S3: Natural variability in ZmSBEIII were significantly correlated with starch.
Author Contributions
C.X., P.L. and Z.Y. gave the project and financial support. H.W., P.L. and C.X. contributed ideas and designs to the subject. T.Y. and Q.L. conducted experiments. W.R., Q.L. and Y.W. analyzed the data. Q.L., C.X. and P.L. wrote and review the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31972487, 31902101, 32172009, and 32061143030), the National Key Research and Development Program of China (2022YFD1201804), the Jiangsu Province University Basic Science Research Project (21KJA210002), the Innovative Research Team of Universities in Jiangsu Province (none), the High-end Talent Project of Yangzhou University, and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), Qing Lan Project of Jiangsu Province.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data sets supporting the results of this article are included within the article (and its Supplementary Files).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, J.; Chen, J.; Yi, Q.; Hu, Y.; Liu, H.; Liu, Y.; Huang, Y. Novel role of ZmaNAC36 in co-expression of starch synthetic genes in maize endosperm. Physiol. Mol. Biol. Plants 2014, 84, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Nelson, O.; Pan, D. Starch Synthesis in Maize Endosperms. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 475–496. [Google Scholar] [CrossRef]
- Tetlow, I.J.; Emes, M.J. Starch Biosynthesis in the Developing Endosperms of Grasses and Cereals. Agronomy 2017, 7, 81. [Google Scholar] [CrossRef]
- Martin, C.; Smith, A.M. Starch Biosynthesis. Plant Cell 1995, 7, 971–985. [Google Scholar] [PubMed]
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 22. [Google Scholar] [CrossRef] [PubMed]
- Tetlow, I.J. Starch biosynthesis in developing seeds. Seed Sci. Res. 2011, 21, 5–32. [Google Scholar] [CrossRef]
- Mizuno, K.; Kawasaki, T.; Shimada, H.; Satoh, H.; Kobayashi, E.; Okumura, S.; Arai, Y.; Baba, T. Alteration of the structural properties of starch components by the lack of an isoform of starch branching enzyme in rice seeds. J. Biol. Chem. 1993, 268, 19084–19091. [Google Scholar] [CrossRef]
- Gao, M.; Fisher, D.K.; Kim, K.N.; Shannon, J.C.; Guiltinan, M.J. Independent Genetic Control of Maize Starch-Branching Enzymes IIa and IIb (Isolation and Characterization of a Sbe2a cDNA). Plant Physiol. 1997, 114, 69–78. [Google Scholar] [CrossRef]
- Regina, A.; Kosar-Hashemi, B.; Li, Z.; Pedler, A.; Mukai, Y. Starch branching enzyme IIb in wheat is expressed at low levels in the endosperm compared to other cereals and encoded at a non-syntenic locus. Planta 2005, 222, 899–909. [Google Scholar] [CrossRef]
- Han, Y.; Sun, F.J.; Rosales-Mendoza, S.; Korban, S.S. Three orthologs in rice, Arabidopsis, and Populus encoding starch branching enzymes (SBEs) are different from other SBE gene families in plants. Gene 2007, 401, 123–130. [Google Scholar] [CrossRef]
- Kang, G.; Li, S.; Zhang, M.; Peng, H.; Wang, C.; Zhu, Y.; Guo, T. Molecular cloning and expression analysis of the starch-branching enzyme III gene from common wheat (Triticum aestivum). Biochem. Genet. 2013, 51, 377–386. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Bodnar, A.L.; Scott, M.P. Wide variability in kernel composition, seed characteristics, and zein profiles among diverse maize inbreds, landraces, and teosinte. Theor. Appl. Genet. 2009, 119, 1129–1142. [Google Scholar] [CrossRef]
- Li, P.; Wei, J.; Wang, H.; Fang, Y.; Yin, S.; Xu, Y.; Liu, J.; Yang, Z.; Xu, C. Natural Variation and Domestication Selection of ZmPGP1 Affects Plant Architecture and Yield-Related Traits in Maize. Genes 2019, 10, 664. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T.M.; Chunwongse, J.; Tanksley, S.D. Microprep Protocol for Extraction of DNA from Tomato and other Herbaceous Plants. Plant Mol. Biol. Report. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Asan; Xu, Y.; Jiang, H.; Tyler-Smith, C.; Xue, Y.; Jiang, T.; Wang, J.; Wu, M.; Liu, X.; Tian, G.; et al. Comprehensive comparison of three commercial human whole-exome capture platforms. Genome Biol. 2011, 12, R95. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J. DNA sequence polymorphism analysis using DnaSP. Methods Mol. Biol. 2009, 537, 337–350. [Google Scholar] [PubMed]
- Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical Tests of Neutrality of Mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, E.; Jiang, Y.; Xu, S.; Pan, L.; Chen, Q.; Xu, C. Sequence polymorphisms in Zmisa2 gene are significantly associated with starch pasting and gelatinization properties in maize (Zea mays L.). Mol. Breed. 2014, 34, 1833–1842. [Google Scholar] [CrossRef]
- Zhang, E.; Yang, Z.; Wang, Y.; Hu, Y.; Song, X.; Xu, C. Nucleotide polymorphisms and haplotype diversity of RTCS gene in China elite maize inbred lines. PLoS ONE 2013, 8, e56495. [Google Scholar] [CrossRef]
- Li, X.; Zhu, C.; Yeh, C.T.; Wu, W.; Takacs, E.M.; Petsch, K.A.; Tian, F.; Bai, G.; Buckler, E.S.; Muehlbauer, G.J.; et al. Genic and nongenic contributions to natural variation of quantitative traits in maize. Genome Res. 2012, 22, 2436–2444. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Warburton, M.; Crouch, J. Association Mapping for Enhancing Maize (Zea mays L.) Genetic Improvement. Crop Sci. 2011, 51, 433–449. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, E.; Li, J.; Jiang, Y.; Wang, Y.; Hu, Y.; Xu, C. Analyses of sequence polymorphism and haplotype diversity of LEAFY genes revealed post-domestication selection in the Chinese elite maize inbred lines. Mol. Biol. Rep. 2014, 41, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Zhao, H.; Ren, L.; Song, W.; Zeng, B.; Guo, J.; Wang, B.; Liu, Z.; Chen, J.; Li, W.; et al. Genome-wide genetic changes during modern breeding of maize. Nat. Genet. 2012, 44, 812–815. [Google Scholar] [CrossRef]
- Liang, Y.; Liu, Q.; Wang, X.; Huang, C.; Xu, G.; Hey, S.; Lin, H.-Y.; Li, C.; Xu, D.; Wu, L.; et al. ZmMADS69 functions as a flowering activator through the ZmRap2.7-ZCN8 regulatory module and contributes to maize flowering time adaptation. New Phytol. 2019, 221, 2335–2347. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Ge, S.; Jensen, J.D.; Hu, F.; Li, X.; Dong, Y.; Gutenkunst, R.N.; Fang, L.; Huang, L.; et al. Resequencing 50 accessions of cultivated and wild rice yields markers for identifying agronomically important genes. Nat. Biotechnol. 2012, 30, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Peng, Z.; Yang, X.; Wang, W.; Fu, J.; Wang, J.; Han, Y.; Chai, Y.; Guo, T.; Yang, N. Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels. Nat. Genet. 2013, 45, 43–50. [Google Scholar] [CrossRef]
- Kandianis, C.B.; Yan, J.; Harjes, C.E.; Bai, L.; Kim, E.H.; Yang, X.; Skinner, D.J.; Fu, Z.; Mitchell, S.; Li, Q. Rare genetic variation at Zea mays crtRB1 increases beta-carotene in maize grain. Nat. Genet. 2010, 42, 322. [Google Scholar]
- Yang, Z.; Ma, S.; Hu, Y.; Zhang, E.; Xie, Z.; Xu, S.; Liu, L.; Deng, L.; Xu, C.; Huang, J. Association Analysis of the Maize Gene ZmYS1 with Kernel Mineral Concentrations. Plant Mol. Biol. Report. 2015, 33, 1327–1335. [Google Scholar] [CrossRef]
- Doebley, J. The Genetics of Maize Evolution. Ann. Rev. Genet. 2004, 38, 37–59. [Google Scholar] [CrossRef]
- Yamasaki, M.; Wright, S.I.; Mcmullen, M. Genomic screening for artificial selection during domestication and improvement in maize. Ann. Bot. 2007, 100, 967–973. [Google Scholar] [CrossRef]
- Chen, W.; Chen, L.; Zhang, X.; Yang, N.; Guo, J.; Wang, M.; Ji, S.; Zhao, X.; Yin, P.; Cai, L.; et al. Convergent selection of a WD40 protein that enhances grain yield in maize and rice. Science 2022, 375, eabg7985. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Tenaillon, M.I.; Vroh Bi, I.; Schroeder, S.G.; Sanchez-Villeda, H.; Doebley, J.F.; Gaut, B.S.; McMullen, M.D. A Large-Scale Screen for Artificial Selection in Maize Identifies Candidate Agronomic Loci for Domestication and Crop Improvement. Plant Cell 2005, 17, 2859–2872. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).