
Phylogeny of the Diploid Species of Rubus (Rosaceae)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Genotyping by sequencing (GBS) Dataset
2.3. Phylogenetic Analysis
3. Results
3.1. GBS Data Analysis Summary
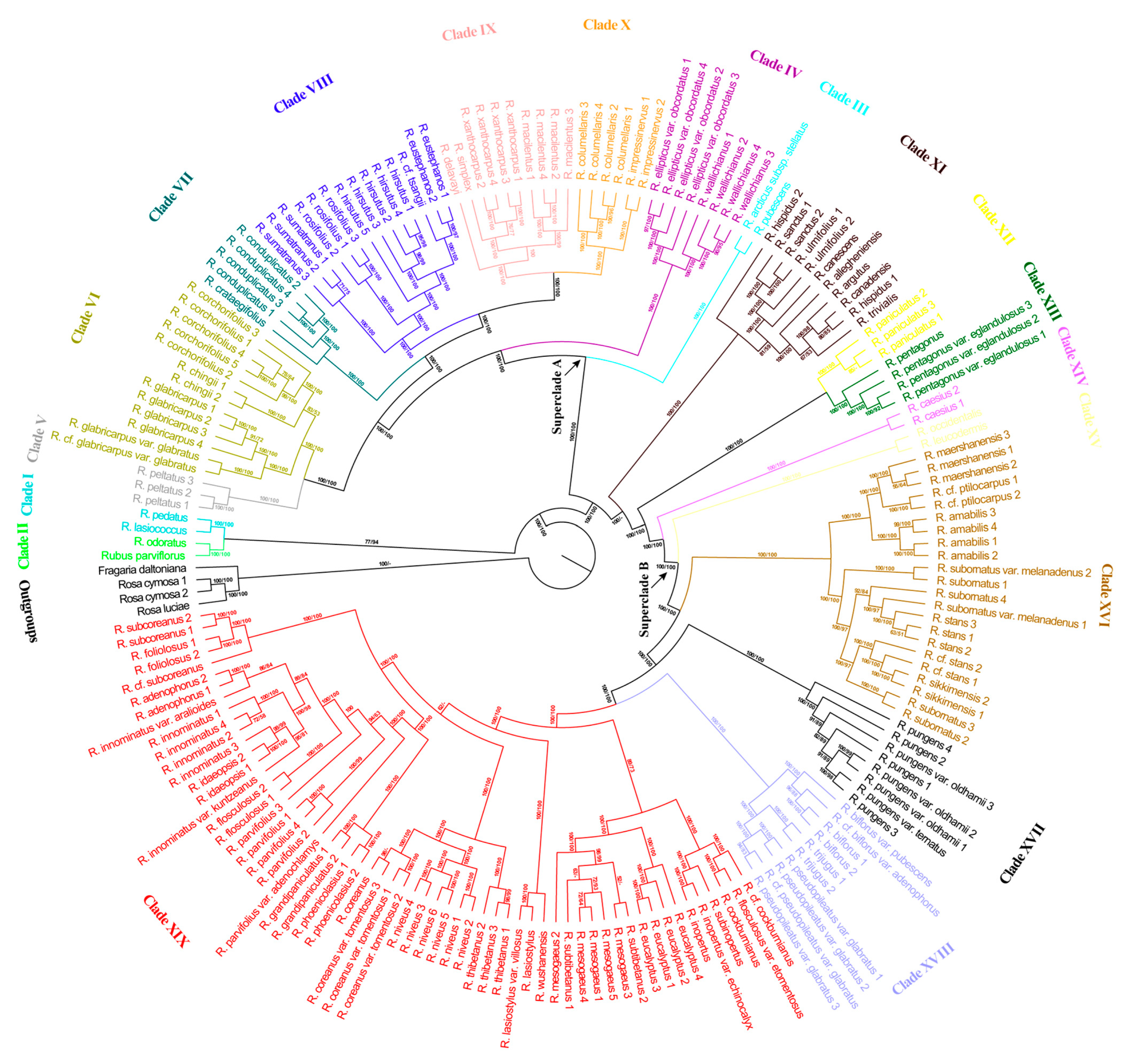
3.2. Phylogenetic Tree Based on SNPs of Nuclear Genome
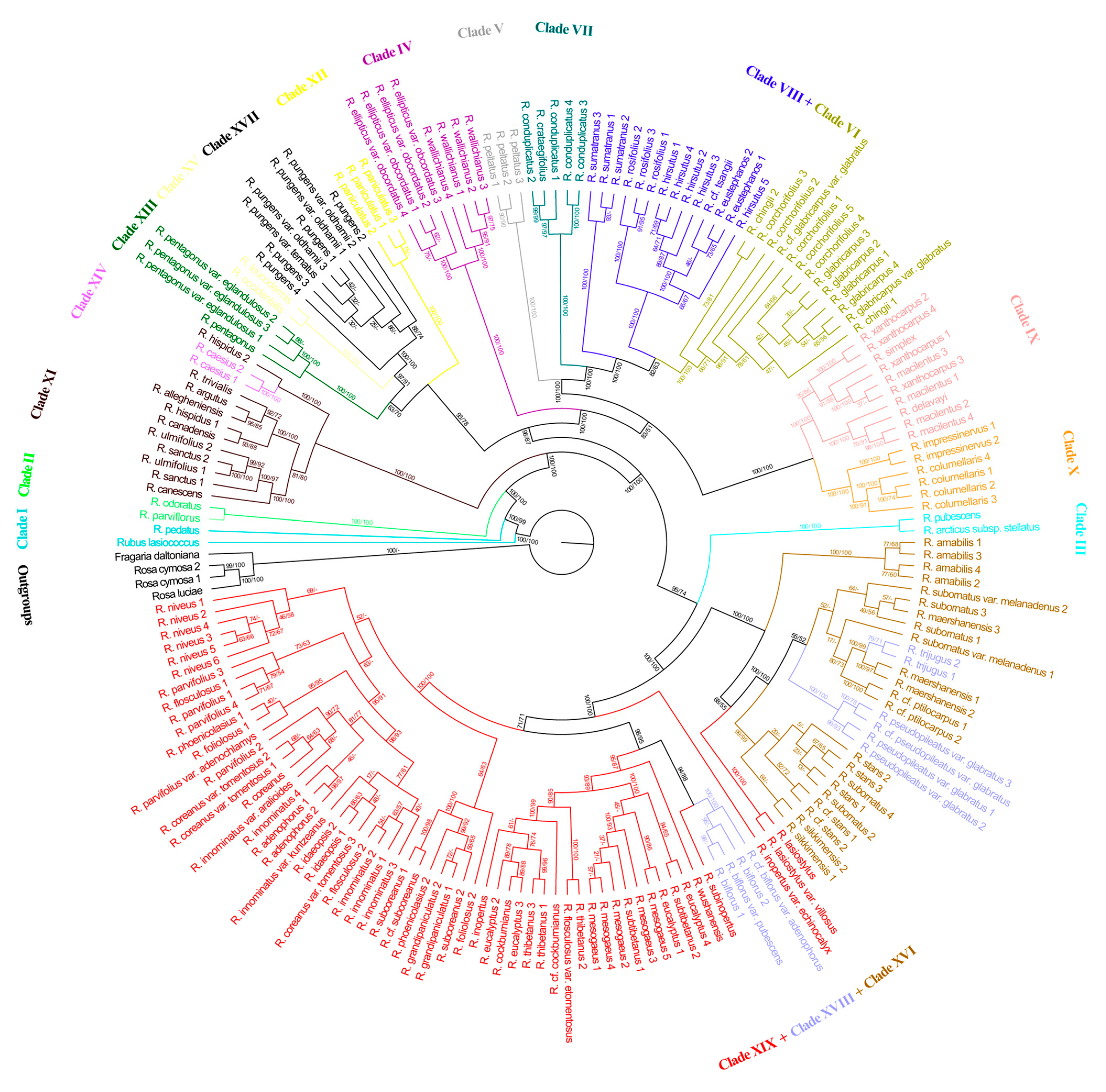
3.3. Phylogenetic Tree Based on SNPs of Chloroplast Genome
4. Discussion
4.1. Phylogenetic Incongruence between the Chloroplast and Nuclear Phylogenies
4.2. Major Evolutionary Lineages within Rubus Revealed in This Study
4.3. A Preliminary Classification Scheme of the Diploid Species of Rubus
- Group 1
- Group 2
- Group 3
- Group 4
- Group 5
- Group 6
- Group 7
- Group 8
- Group 9
- Group 10
- Group 11
- Group 12
- Group 13
- Group 14
- Group 15
- Group 16
- Group 17
- Group 18
- Group 19
- Group 20
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Taxon | Subgeneric Classification of Focke [9, 10,11] | Sectional Classification of Yu and Lu [83], Lu and Yu [6], and Lu and Boufford [7] | Ploidy Level | Voucher |
|---|---|---|---|---|
| Rubus odoratus L. | R. subg. Anoplobatus | — | 2x | USA: Massachusetts, Northampton, Ano. s. n. (BR, Living Plant Collections Accession 19941489-35) |
| R. parviflorus Nutt. | R. subg. Anoplobatus | — | 2x | USA: W. R. Smith 29200 (MO) |
| R. arcticus L. subsp. stellatus (Sm.) B. Boivin | R. subg. Cylactis | — | 2x | USA: Alaska, Wasilla, K. Hummer et C. I. Wright s. n. (NPGS, Accession PI 606535) |
| R. pubescens Raf. | — | — | 2x | USA: D. Atha 7040 (MO) |
| R. simplex Focke | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Leibo, X. H. Xiong 2036 (CDBI) |
| R. xanthocarpus Bureau et Franch. 1 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1159 (CDBI) |
| R. xanthocarpus Bureau et Franch. 2 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1316 (CDBI) |
| R. xanthocarpus Bureau et Franch. 3 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2002 (CDBI) |
| R. xanthocarpus Bureau et Franch. 4 | R. subg. Cylactis | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 2020 (CDBI) |
| R. lasiococcus A. Gray | R. subg. Dalibarda | — | 2x | USA: Oregon, Deschutes County, O. L. Jahn s. n. (NPGS, Accession PI 553659) |
| R. pedatus Sm. | R. subg. Dalibarda | — | 2x | USA: Alaska, Sitka, H. Chambers s. n. (NPGS, Accession PI618503) |
| R. adenophorus Rolfe 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1967 (CDBI) |
| R. adenophorus Rolfe 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1968 (CDBI) |
| R. amabilis Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1146-3 (CDBI) |
| R. amabilis Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1146-7 (CDBI) |
| R. amabilis Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1863 (CDBI) |
| R. amabilis Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Wenchuan, X. H. Xiong 1870 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Xiaojin, X. H. Xiong 1860 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Yajiang, X. H. Xiong 2156 (CDBI) |
| R. biflorus Buch.-Ham. ex Sm. var. pubescens T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Muli, X. H. Xiong 1365 (CDBI) |
| R. cf. biflorus Buch.-Ham. ex Sm. var. adenophorus Franch. | — | R. sect. Idaeobatus | NA | China: Xizang, Linzhi, X. H. Xiong 1054 (CDBI) |
| R. chingii Hu 1 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1966 (CDBI) |
| R. chingii Hu 2 | — | R. sect. Idaeobatus | 2x | China: Anhui, Huangshan, X. H. Xiong 831 (CDBI) |
| R. cockburnianus Hemsl. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Maoxian, X. F. Gao et al. 4983 (CDBI) |
| R. cf. cockburnianus Hemsl. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Bomi, Y. D. Gao et M. Li YLZB0219 (CDBI) |
| R. columellaris Tutcher 1 | — | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1389 (CDBI) |
| R. columellaris Tutcher 2 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1735 (CDBI) |
| R. columellaris Tutcher 3 | — | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1789 (CDBI) |
| R. columellaris Tutcher 4 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1970 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 1 (R. trianthus Focke) | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1648 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1836-3 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1912 (CDBI) |
| R. conduplicatus Duthie ex J. H. Veitch 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1964 (CDBI) |
| R. corchorifolius L. f. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1634 (CDBI) |
| R. corchorifolius L. f. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1733 (CDBI) |
| R. corchorifolius L. f. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1742 (CDBI) |
| R. corchorifolius L. f. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1828-2 (CDBI) |
| R. corchorifolius L. f. 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1918 (CDBI) |
| R. coreanus Miq. | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Xiong s. n. (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 1 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1173 (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 2 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1176 (CDBI) |
| R. coreanus Miq. var. tomentosus Cardot 3 | — | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1771 (CDBI) |
| R. crataegifolius Bunge | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Beijing, X. H. Xiong 1991 (CDBI) |
| R. delavayi Franch. | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Yunnan, Shizong, X. H. Xiong 1824 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1427 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Maguan, X. H. Xiong 1709 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1737 (CDBI) |
| R. ellipticus Sm. var. obcordatus (Franch.) Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1758 (CDBI) |
| R. eucalyptus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1304 (CDBI) |
| R. eucalyptus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1839 (CDBI) |
| R. eucalyptus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Dayi, X. H. Xiong 1856 (CDBI) |
| R. eucalyptus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1993 (CDBI) |
| R. eustephanos Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1534 (CDBI) |
| R. eustephanos Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1784 (CDBI) |
| R. flosculosus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Xianju, S. Q. Xu 1299 (CDBI) |
| R. flosculosus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Suichang, S. Q. Xu 1344 (CDBI) |
| R. flosculosus Focke var. etomentosus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2014 (CDBI) |
| R. foliolosus D. Don 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 4x | China: Yunnan, Malipo, X. H. Xiong 1712 (CDBI) |
| R. foliolosus D. Don 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 4x | China: Yunnan, X. H. Xiong 1744 (CDBI) |
| R. glabricarpus Cheng 1 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Wenzhou, X. H. Xiong 1838-1 (CDBI) |
| R. glabricarpus Cheng 2 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Wenzhou, X. H. Xiong 1838-3 (CDBI) |
| R. glabricarpus Cheng 3 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1947 (CDBI) |
| R. glabricarpus Cheng 4 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1949 (CDBI) |
| R. glabricarpus Cheng var. glabratus C. Z. Zheng et Y. Y. Fang | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1933 (CDBI) |
| R. cf. glabricarpus Cheng var. glabratus C. Z. Zheng et Y. Y. Fang | — | R. sect. Idaeobatus | NA | China: Fujian, Wuyishan, M. Li LMWYS-45 (CDBI) |
| R. grandipaniculatus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wuxi, X. H. Xiong 2080 (CDBI) |
| R. grandipaniculatus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Chongqing, Wuxi, X. H. Xiong 2087 (CDBI) |
| R. hirsutus Thunb. 1 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Hangzhou, X. H. Xiong 1594 (CDBI) |
| R. hirsutus Thunb. 2 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Lushan, X. H. Xiong 1635 (CDBI) |
| R. hirsutus Thunb. 3 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1734 (CDBI) |
| R. hirsutus Thunb. 4 | — | R. sect. Idaeobatus | 2x | China: Zhejiang, Wenzhou, X. H. Xiong 1837-1 (CDBI) |
| R. hirsutus Thunb. 5 | — | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1983 (CDBI) |
| R. idaeopsis Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1627 (CDBI) |
| R. idaeopsis Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1628 (CDBI) |
| R. impressinervus F. P. Metcalf 1 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1930 (CDBI) |
| R. impressinervus F. P. Metcalf 2 | — | R. sect. Idaeobatus | NA | China: Zhejiang, Qingyuan, X. H. Xiong 1935 (CDBI) |
| R. innominatus S. Moore 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Wuxi, Ano. THP-WX-1601 (CDBI) |
| R. innominatus S. Moore 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Chengkou, X. H. Xiong 1220 (CDBI) |
| R. innominatus S. Moore 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1300 (CDBI) |
| R. innominatus S. Moore 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1788 (CDBI) |
| R. innominatus S. Moore var. aralioides (Hance) T. T. Yu et L. T. Lu | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Jiangxi, Lushan, X. H. Xiong 1645 (CDBI) |
| R. innominatus S. Moore var. kuntzeanus (Hemsl.) L. H. Bailey | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Xuyong, X. H. Xiong 1551 (CDBI) |
| R. inopertus (Focke) Focke | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1985 (CDBI) |
| R. inopertus (Focke) Focke var. echinocalyx Cardot | — | R. sect. Idaeobatus | NA | China: Sichuan, Xuyong, X. H. Xiong 1562 (CDBI) |
| R. lasiostylus Focke | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Xian, C. Zhang et S. Z. Yang zc12087 (CDBI) |
| R. lasiostylus Focke var. villosus Cardot | — | R. sect. Idaeobatus | NA | China: Chongqing, Chengkou, X. H. Xiong 1209 (CDBI) |
| R. leucodermis Douglas ex Hook. | R. subg. Idaeobatus | — | 2x | Canada: British Columbia, H. A. Daubeny s. n. (NPGS, Accession PI 618562) |
| R. macilentus Jacquem. ex Cambess. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1418 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1430 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1535 (CDBI) |
| R. macilentus Jacquem. ex Cambess. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1763 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 1 (R. lutescens Franch.) | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1259 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Muli, X. H. Xiong 1362 (CDBI) |
| R. maershanensis Huan C. Wang et H. Sun 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1786 (CDBI) |
| R. mesogaeus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1080 (CDBI) |
| R. mesogaeus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Cuona, X. H. Xiong 1575 (CDBI) |
| R. mesogaeus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Shimian, X. H. Xiong 1861 (CDBI) |
| R. mesogaeus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2007 (CDBI) |
| R. mesogaeus Focke 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Jinyang, X. H. Xiong 2030 (CDBI) |
| R. niveus Thunb. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Kangding, X. H. Xiong 1268 (CDBI) |
| R. niveus Thunb. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Yajiang, X. H. Xiong 1345 (CDBI) |
| R. niveus Thunb. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Malipo, X. H. Xiong 1718 (CDBI) |
| R. niveus Thunb. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1741 (CDBI) |
| R. niveus Thunb. 5 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1787 (CDBI) |
| R. niveus Thunb. 6 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luding, X. H. Xiong 1862 (CDBI) |
| R. occidentalis L. | R. subg. Idaeobatus | — | 2x | USA: Oregon, Ano. s. n. (NPGS, Accession PI 672663) |
| R. parvifolius L. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Sichuan, Chengdu, X. H. Xiong 1528 (CDBI) |
| R. parvifolius L. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Zhejiang, Hangzhou, X. H. Xiong 1591 (CDBI) |
| R. parvifolius L. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Anhui, Susong, X. H. Xiong 1703 (CDBI) |
| R. parvifolius L. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x, 3x, 4x | China: Sichuan, Luzhou, X. H. Xiong 1804 (CDBI) |
| R. parvifolius L. var. adenochlamys (Focke) Migo | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Shaanxi, Taibai, X. H. Xiong 1333 (CDBI) |
| R. peltatus Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1938 (CDBI) |
| R. peltatus Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1942 (CDBI) |
| R. peltatus Maxim. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1987 (CDBI) |
| R. pentagonus Wall. | R. subg. Idaeobatus | R. sect. Idaeobatus | 4x | China: Xizang, Motuo, X. H. Xiong 1409 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, B. Xu et X. H. Xiong YLZB1151 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, B. Xu et X. H. Xiong YLZB1174 (CDBI) |
| R. pentagonus Wall. var. eglandulosus T. T. Yu et L. T. Lu 3 | — | R. sect. Idaeobatus | NA | China: Xizang, Motuo, X. H. Xiong 1406 (CDBI) |
| R. phoenicolasius Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Hanzhong, X. H. Xiong 1322 (CDBI) |
| R. phoenicolasius Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Taibai, X. H. Xiong 1334 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-1 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-27 (CDBI) |
| R. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu 3 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 1147-8 (CDBI) |
| R. cf. pseudopileatus Cardot var. glabratus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Sichuan, Luding, X. H. Xiong 1868 (CDBI) |
| R. cf. ptilocarpus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2003 (CDBI) |
| R. cf. ptilocarpus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2005 (CDBI) |
| R. pungens Cambess. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 1162 (CDBI) |
| R. pungens Cambess. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Wushan, X. H. Xiong 1186 (CDBI) |
| R. pungens Cambess. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Chongqing, Chengkou, X. H. Xiong 1200 (CDBI) |
| R. pungens Cambess. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Milin, B. Xu et X. M. Zhou YLZB1613 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Shaanxi, Taibai, X. H. Xiong 1332 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Gansu, Longnan, X. H. Xiong 1337 (CDBI) |
| R. pungens Cambess. var. oldhamii (Miq.) Maxim. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Baoxing, X. H. Xiong 2008 (CDBI) |
| R. pungens Cambess. var. ternatus Cardot | — | R. sect. Idaeobatus | NA | China: Chongqing, Chengkou, X. H. Xiong 1197 (CDBI) |
| R. rosifolius Sm. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Ganzhou, X. H. Xiong 1732 (CDBI) |
| R. rosifolius Sm. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Zhejiang, Qingyuan, X. H. Xiong 1913 (CDBI) |
| R. rosifolius Sm. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1975 (CDBI) |
| R. sikkimensis Hook. f. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Motuo, X. H. Xiong 984 (CDBI) |
| R. sikkimensis Hook. f. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Motuo, W. B. Ju et H. N. Deng YLZB0833 (CDBI) |
| R. stans Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, Shangri-la, X. H. Xiong 1996 (CDBI) |
| R. stans Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Jinyang, X. H. Xiong 2021 (CDBI) |
| R. stans Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Muli, X. H. Xiong 2031 (CDBI) |
| R. cf. stans Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Cuona, X. H. Xiong 1577 (CDBI) |
| R. cf. stans Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Xizang, Milin, X. H. Xiong 1584 (CDBI) |
| R. subcoreanus T. T. Yu et L. T. Lu 1 | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1327 (CDBI) |
| R. subcoreanus T. T. Yu et L. T. Lu 2 | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1329 (CDBI) |
| R. cf. subcoreanus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Shaanxi, Hanzhong, X. H. Xiong 1328 (CDBI) |
| R. subinopertus T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1407 (CDBI) |
| R. subornatus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1314 (CDBI) |
| R. subornatus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Linzhi, X. H. Xiong 1374 (CDBI) |
| R. subornatus Focke 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Xizang, Milin, X. H. Xiong 1582 (CDBI) |
| R. subornatus Focke 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Jinyang, X. H. Xiong 2028 (CDBI) |
| R. subornatus Focke var. melanadenus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1264 (CDBI) |
| R. subornatus Focke var. melanadenus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Baoxing, X. H. Xiong 2011 (CDBI) |
| R. subtibetanus Hand.-Mazz. 1 | — | R. sect. Idaeobatus | NA | China: Sichuan, X. H. Xiong 1762 (CDBI) |
| R. subtibetanus Hand.-Mazz. 2 | — | R. sect. Idaeobatus | NA | China: Sichuan, Kangding, X. H. Xiong 1999 (CDBI) |
| R. sumatranus Miq. 1 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1911, correct) | R. sect. Idaeobatus | 2x | China: Xizang, Motuo, X. H. Xiong 1392 (CDBI) |
| R. sumatranus Miq. 2 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1912, correct) | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1805 (CDBI) |
| R. sumatranus Miq. 3 | R. subg. Malachobatus (1910, erroneous), R. subg. Idaeobatus (1913, correct) | R. sect. Idaeobatus | 2x | China: Jiangxi, Jinggangshan, X. H. Xiong 1989 (CDBI) |
| R. thibetanus Franch. 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Gansu, Longnan, X. H. Xiong 1336 (CDBI) |
| R. thibetanus Franch. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1529 (CDBI) |
| R. thibetanus Franch. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1764 (CDBI) |
| R. trijugus Focke 1 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Danba, X. H. Xiong 2167 (CDBI) |
| R. trijugus Focke 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | NA | China: Sichuan, Jinchuan, X. H. Xiong 2175 (CDBI) |
| R. cf. tsangii Merr. | — | R. sect. Idaeobatus | 2x | China: Sichuan, X. H. Xiong 1783 (CDBI) |
| R. wallichianus Wight et Arn. 1 (R. pinfaensis H. Lév. et Vaniot) | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Yunnan, X. H. Xiong 1743 (CDBI) |
| R. wallichianus Wight et Arn. 2 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Chengdu, X. H. Xiong 1785 (CDBI) |
| R. wallichianus Wight et Arn. 3 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Sichuan, Luzhou, X. H. Xiong 1799 (CDBI) |
| R. wallichianus Wight et Arn. 4 | R. subg. Idaeobatus | R. sect. Idaeobatus | 2x | China: Guizhou, X. H. Xiong 1820 (CDBI) |
| R. wushanensis T. T. Yu et L. T. Lu | — | R. sect. Idaeobatus | NA | China: Chongqing, Wushan, X. H. Xiong 1183 (CDBI) |
| R. paniculatus Sm. 1 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 966 (CDBI) |
| R. paniculatus Sm. 2 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 969 (CDBI) |
| R. paniculatus Sm. 3 | R. subg. Malachobatus | R. sect. Malachobatus | 2x | China: Xizang, Motuo, X. H. Xiong 971 (CDBI) |
| R. allegheniensis Porter | R. subg. Rubus | — | 2x, 3x, 4x | USA: Pennsylvania, Lehighton, T. A. Merkel s. n. (NPGS, Accession PI 553970) |
| R. argutus Link | R. subg. Rubus | — | 2x, 3x | USA: South Carolina, J. R. Ballington et J. A. Payne s. n. (NPGS, Accession PI 606490) |
| R. caesius L. 1 | R. subg. Rubus | R. sect. Rubus | 4x | Russia: Stavropol, H. J. Brooks s. n. (NPGS, Accession PI 324058) |
| R. caesius L. 2 | R. subg. Rubus | R. sect. Rubus | 4x | Spain: Zaragoza, Erickson s. n. (NPGS, Accession PI 420407) |
| R. canadensis L. | R. subg. Rubus | — | 2x, 3x | USA: Georgia, J. R. Ballington s. n. (NPGS, Accession PI 553141) |
| R. canescens DC. | R. subg. Rubus | — | 2x | Former Serbia and Montenegro: L. Aladzajkov s. n. (NPGS, Accession PI 379537) |
| R. hispidus L. 1 | R. subg. Rubus | — | 2x | USA: North Carolina, Beaufort County, J. R. Ballington s. n. (NPGS, Accession PI 606507) |
| R. hispidus L. 2 | R. subg. Rubus | — | 2x | USA: Tennessee, Cumberland County, Ano. s. n. (NPGS, Accession PI 618281) |
| R. sanctus Schreb. 1 | R. subg. Rubus | — | 2x | Pakistan: Chitral District, D. Brenner et M. Thompson s. n. (NPGS, Accession PI 553879) |
| R. sanctus Schreb. 2 | R. subg. Rubus | — | 2x | Syria: Damascus, M. Thompson s. n. (NPGS, Accession PI 553930) |
| R. trivialis Michx. | R. subg. Rubus | — | 2x | USA: Texas, Austin, L. Whiting s. n. (NPGS, Accession PI 553704) |
| R. ulmifolius Schott 1 | R. subg. Rubus | — | 2x | Syria: Damascus, M. Thompson s. n. (NPGS, Accession PI 553931) |
| R. ulmifolius Schott 2 | R. subg. Rubus | — | 2x | USA: California, Napa County, L. Clark s. n. (NPGS, Accession PI 672650) |
| Outgroups | ||||
| Fragaria daltoniana J. Gay | 2x | China: Xizang, Milin, X. H. Xiong 1590 (CDBI) | ||
| Rosa cymosa Tratt. 1 | 2x | China: Jiangxi, Dayu, X. H. Xiong 803-2 (CDBI) | ||
| R. cymosa Tratt. 2 | 2x | China: Jiangxi, Xingan, X. H. Xiong 813-1 (CDBI) | ||
| R. luciae Franch. et Rochebr. | 2x | China: Zhejiang, Wenzhou, C. Zhang s. n. (CDBI) |
References
- Alice, L.A.; Campbell, C.S. Phylogeny of Rubus (Rosaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Am. J. Bot. 1999, 86, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Alice, L.A.; Goldman, D.H.; Macklin, J.A.; Moore, G. Rubus L. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2014; Volume 9, pp. 28–56. [Google Scholar]
- Bailey, L.H. Gentes Herbarum: Rubus in North America; Ithaca: New York, NY, USA, 1945; Volume 5, pp. 1–932. [Google Scholar]
- Thuan, N. Flore du Cambodge du Laos et du Vietnam; Fascicule 7; Museum National D’Histoire Naturelle: Paris, France, 1968; pp. 1–83. [Google Scholar]
- Lu, L.T. A study on the genus Rubus of China. Acta Phytotax. Sin. 1983, 21, 13–25. [Google Scholar]
- Lu, L.T.; Yu, T.T. Rubus L. In Flora Reipublicae Popularis Sinicae; Yu, T.T., Ed.; Science Press: Beijing, China, 1985; Volume 37, pp. 10–218. [Google Scholar]
- Lu, L.T.; Boufford, D.E. Rubus L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 9, pp. 195–285. [Google Scholar]
- Abrams, L. Illustrated Flora of the Pacific States, Washington, Oregon, and California; Stanford University Press: Stanford, CA, USA, 1944; Volume Ⅱ, pp. 454–458. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi prodromus, Pars I. Biblioth. Bot. 1910, 17, 1–120. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi Prodromus, Pars II. Biblioth. Bot. 1911, 17, 121–223. [Google Scholar]
- Focke, W.O. Species Ruborum, monographiae generis Rubi Prodromus, Pars III. Biblioth. Bot. 1914, 19, 1–274. [Google Scholar]
- Royen, P. The Alpine Flora of New Guinea; Strauss et Cramer: Hirschberg, Germany, 1983; Volume 4, pp. 2458–2490. [Google Scholar]
- Thompson, M.M. Survey of chromosome numbers in Rubus (Rosaceae: Rosoideae). Ann. Mo. Bot. Gard. 1997, 84, 128–164. [Google Scholar] [CrossRef]
- Howarth, D.G.; Gardner, D.E.; Morden, C.W. Phylogeny of Rubus subgenus Idaeobatus (Rosaceae) and its implications toward colonization of the Hawaiian Islands. Syst. Bot. 1997, 22, 433–441. [Google Scholar] [CrossRef]
- Gustafsson, A. The genesis of the European blackberry flora. Lunds Univ. Arsskrift. 1943, 39, 1–200. [Google Scholar]
- Aalders, L.E.; Hall, I.V. A cytotaxonomic survey of the native blackberries of Nova Scotia. Canad. J. Genet. Cytol. 1966, 8, 528–532. [Google Scholar] [CrossRef]
- Richards, A.J.; Kirschner, J.; Stepanek, J.; Marhold, K. Apomixis and taxonomy: An introduction. Folia Geobot. Phytotax. 1996, 31, 281–282. [Google Scholar] [CrossRef]
- Weber, H.E. Former and modern taxonomic treatment of the apomictic Rubus complex. Folia Geobot. Phytotax. 1996, 31, 373–380. [Google Scholar] [CrossRef]
- Robertson, K.R. The genera of Rosaceae in the southeastern United States. J. Arnold Arbor. 1974, 55, 352–360. [Google Scholar] [CrossRef]
- Thompson, M.M. Chromosome numbers of Rubus species at the National Clonal Germplasm Repository. Hortscience 1995, 30, 1447–1452. [Google Scholar] [CrossRef]
- Alice, L.A.; Dodson, T.M.; Sutherland, B.L. Diversity and relationships of Bhutanese Rubus. Acta Hort. 2008, 777, 63–69. [Google Scholar] [CrossRef]
- Waugh, R.; van de Ven, W.T.G.; Phillips, M.S.; Powell, W. Chloroplast DNA diversity in the genus Rubus (Rosaceae) revealed by Southern hybridization. Plant Syst. Evol. 1990, 172, 65–75. [Google Scholar] [CrossRef]
- Gustafsson, A. Chromosomenzahlen in der gattung Rubus. Hereditas 1933, 18, 77–80. [Google Scholar] [CrossRef]
- Gustafsson, A. Differential polyploidy within the blackberries. Hereditas 1939, 25, 33–47. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y. Cytological studies in the Genus Rubus L. Ⅰ. Chromosome numbers in the British Rubus Flora. New Phytol. 1953, 52, 22–39. [Google Scholar] [CrossRef]
- Bammi, R.K. Complement fractionation in a natural hybrid between Rubus procerus Muell. and R. laciniatus Willd. Nature 1965, 208, 608. [Google Scholar] [CrossRef]
- Malik, C.P. Cytology of some Indian species of Rosaceae. Caryologia 1965, 18, 139–149. [Google Scholar] [CrossRef]
- Nybom, H. Chromosome numbers in Rubus species from Sri Lanka. Bot. Not. 1980, 133, 47–48. [Google Scholar]
- Nybom, H. Chromosome numbers and reproduction in Rubus subgen. Malachobatus. Plant Syst. Evol. 1986, 152, 211–218. [Google Scholar] [CrossRef]
- Dale, A.; Ingram, R. Chromosome numbers of some South-American blackberries. Hort. Res. 1981, 21, 107. [Google Scholar]
- Subramanian, D. Cytotaxonomic studies of South Indian Rosaceae. Cytologia 1987, 52, 395–403. [Google Scholar] [CrossRef]
- Thompson, M.M.; Zhao, C.M. Chromosome numbers of Rubus species in Southwest China. Acta Hort. 1993, 352, 493–502. [Google Scholar] [CrossRef]
- Lin, S.H.; Zhang, B.B.; Fang, C.Q.; Lin, F.Q.; Pu, F.S. Chromosome numbers and karyotypes of 8 Rubus species in China. Acta Hort. Sin. 1994, 21, 313–319. [Google Scholar]
- Iwatsubo, Y.; Naruhashi, N.; Weber, H.E. Chromosome numbers of European blackberries (Rubus subg Rubus, Rosaceae). Plant Syst. Evol. 1995, 198, 143–149. [Google Scholar] [CrossRef]
- Meng, R.G.; Finn, C. Determining ploidy level and nuclear DNA content in Rubus by flow cytometry. J. Am. Soc. Hortic. Sci. 2002, 127, 767–775. [Google Scholar] [CrossRef]
- Naruhashi, N.; Iwatsubo, Y.; Peng, C.I. Chromosome numbers in Rubus (Rosaceae) of Taiwan. Bot. Bull. Acad. Sin. 2002, 43, 193–201. [Google Scholar] [CrossRef]
- Michael, K. Clarification of Basal Relationships in Rubus (Rosaceae) and the Origin of Rubus chamaemorus. Master’s Thesis, Western Kentucky University, Bowling Green, KY, USA, 2006. [Google Scholar]
- Wang, X.R.; Tang, H.R.; Duan, J.; Li, L. A comparative study on karyotypes of 28 taxa in Rubus sect. Idaeobatus and sect. Malachobatus (Rosaceae) from China. J. Syst. Evol. 2008, 46, 505–515. [Google Scholar] [CrossRef]
- Wang, X.R.; Tang, H.R.; Fu, H.Q.; Luo, Y.; Deng, Q.X.; Dong, X.L.; Li, L.; Duan, J. Chromosome numbers and karyotypes of 10 wild bramble (Rubus L.) species from Southwest of China. Acta Hort. Sin. 2008, 35, 343–350. [Google Scholar]
- Krahulcova, A.; Travnicek, B.; Sarhanova, P. Karyological variation in the genus Rubus subgenus Rubus: New data from the Czech Republic and synthesis of the current knowledge of European species. Preslia 2013, 85, 19–39. [Google Scholar]
- Wang, Y.; Wang, X.R.; Chen, Q.; Zhang, L.; Tang, H.R.; Luo, Y.; Liu, Z.J. Phylogenetic insight into subgenera Idaeobatus and Malachobatus (Rubus, Rosaceae) inferring from ISH analysis. Mol. Cytogenet. 2015, 8, 11–23. [Google Scholar] [CrossRef]
- Carter, K.A.; Liston, A.; Bassil, N.V.; Alice, L.A.; Bushakra, J.M.; Sutherland, B.L.; Mockler, T.C.; Bryant, D.W.; Hummer, K.E. Target capture sequencing unravels Rubus evolution. Front. Plant Sci. 2019, 10, 1615. [Google Scholar] [CrossRef]
- Bean, A.R. A revision of Rubus subg. Malachobatus (Focke) Focke and Rubus subg. Diemenicus A. R. Bean (Rosaceae) in Australia. Austrobaileya 1997, 5, 39–51. [Google Scholar]
- Kalkman, C. The genus Rubus (Rosaceae) in Malesia 3. The subgenus Micranthobatus. Blumea 1987, 32, 323–341. [Google Scholar]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.H.; Smedmark, J.E.E.; Campbell, C.S. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Chen, T.; Tang, H.R.; Liu, L.; Wang, X.R. Phylogenetic insights into Chinese Rubus (Rosaceae) from multiple chloroplast and nuclear DNAs. Front. Plant Sci. 2016, 7, 968. [Google Scholar] [CrossRef]
- Morden, C.W.; Gardner, D.E.; Weniger, D.A. Phylogeny and biogeography of Pacific Rubus subgenus Idaeobatus (Rosaceae) species: Investigating the origin of the endemic Hawaiian raspberry R. macraei. Pac. Sci. 2003, 57, 181–197. [Google Scholar] [CrossRef]
- Stafne, E.T.; Clark, J.R.; Szalanski, A.L. Nuclear ribosomal ITS region sequences for differentiation of Rubus genotypes. J. Ark. Acad. Sci. 2003, 57, 176–180. [Google Scholar]
- Sutherland, B. Phylogenetics of Rubus ursinus and R. macraai (Rosaceae): Evidence of Hybrid Origin. Master’s Thesis, Western Kentucky University, Bowling Green, KY, USA, 2005. [Google Scholar]
- Yang, J.Y.; Pak, J.H. Phylogeny of Korean Rubus (Rosaceae) based on ITS (nrDNA) and trn L/F intergenic region (cpDNA). J. Plant Biol. 2006, 49, 44–54. [Google Scholar] [CrossRef]
- Imanishi, H.; Nakahara, K.; Tsuyuzaki, H. Genetic relationships among native and introduced Rubus species in Japan based on rbcL sequence. Acta Hort. 2008, 769, 195–199. [Google Scholar] [CrossRef]
- Wang, Y.N. Relationships among Rubus (Rosaceae) Species Used in Traditional Chinese Medicine. Master’s Dissertation, Western Kentucky University, Bowling Green, KY, USA, 2011. [Google Scholar]
- Yang, J.Y.; Yoon, H.S.; Pak, J.H. Phylogeny of Korean Rubus (Rosaceae) based on the second intron of the LEAFY gene. Can. J. Plant Sci. 2012, 92, 461–472. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Chen, Q.; Luo, Y.; Zhang, Y.; Tang, H.R.; Wang, X.R. Phylogenetic utility of Chinese Rubus (Rosaceae) based on ndhF sequence. Acta Hort. Sin. 2015, 42, 19–30. [Google Scholar] [CrossRef]
- Sochor, M.; Trávníček, B. Melting pot of biodiversity: First insights into the evolutionary patterns of the Colchic bramble flora (Rubus subgenus Rubus, Rosaceae). Bot. J. Linn. Soc. 2016, 181, 610–620. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, É.; Laroche, J.; Larose, S.; Jean, M.; et al. An improved genotyping by sequencing (GBS) approach offering increased versatility and efficiency of SNP discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef]
- Arbizu, C.I.; Ellison, S.L.; Senalik, D.; Simon, P.W.; Spooner, D.M. Genotyping-by-sequencing provides the discriminating power to investigate the subspecies of Daucus carota (Apiaceae). BMC Evol. Biol. 2016, 16, 234. [Google Scholar] [CrossRef]
- Ariani, A.; Berny Mier y Teran, J.C.; Gepts, P. Genome-wide identification of SNPs and copy number variation in common bean (Phaseolus vulgaris L.) using genotyping-by-sequencing (GBS). Mol. Breed. 2016, 36, 87. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Chong, C.; Bragg, J.G.; Ong, C.R.; Aitken, N.C.; Chuah, A.; Lepschi, B.; Borevitz, J.O. Population and phylogenomic decomposition via genotyping-by-sequencing in Australian Pelargonium. Mol. Ecol. 2016, 25, 2000–2014. [Google Scholar] [CrossRef]
- Stetter, M.G.; Schmid, K.J. Analysis of phylogenetic relationships and genome size evolution of the Amaranthus genus using GBS indicates the ancestors of an ancient crop. Mol. Phylogenet. Evol. 2017, 109, 80–92. [Google Scholar] [CrossRef]
- Wang, X.K.; Jiang, D.; Sun, Z.Z. Study on phylogeny of 240 mandarin accessions with Genotyping-by-Sequencing technology. Sci. Agric. Sin. 2017, 50, 1666–1673. [Google Scholar] [CrossRef]
- Zhao, P.; Zhou, H.J.; Potter, D.; Hu, Y.H.; Feng, X.J.; Dang, M.; Feng, L.; Zulfiqar, S.; Liu, W.Z.; Zhao, G.F.; et al. Population genetics, phylogenomics and hybrid speciation of Juglans in China determined from whole chloroplast genomes, transcriptomes, and genotyping-by-sequencing (GBS). Mol. Phylogenet. Evol. 2018, 126, 250–265. [Google Scholar] [CrossRef]
- Morigengaowa; Shang, H.; Liu, B.D.; Kang, M.; Yan, Y.H. One or more species? GBS sequencing and morphological traits evidence reveal species diversification of Sphaeropteris brunoniana in China. Biodivers. Sci. 2019, 27, 1196–1204. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Sun, B.; Chen, T.; Luo, Y.; Zhang, Y.; Wang, X.R.; Tang, H.R. The complete chloroplast genome sequence of Rubus coreanus, an excellent diseases-resistant resource. Mitochondrial DNA B Resour. 2019, 4, 216–217. [Google Scholar] [CrossRef]
- VanBuren, R.; Wai, C.M.; Colle, M.; Wang, J.; Sullivan, S.; Bushakra, J.M.; Liachko, I.; Vining, K.J.; Dossett, M.; Finn, C.E.; et al. A near complete, chromosome-scale assembly of the black raspberry (Rubus occidentalis) genome. GigaScience 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup, The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Farris, J.S.; Albert, V.A.; Källersjö, M.; Lipscomb, D.; Kluge, A.G. Parsimony jackknifing outperforms neighbor-joining. Cladistics 1996, 12, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Maximum likelihood and minimum-steps methods for estimating evolutionary trees from data on discrete characters. Syst. Biol. 1973, 22, 240–249. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Pol, D. Empirical problems of the hierarchical likelihood ratio test for model selection. Syst. Biol. 2004, 53, 949–962. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Yu, T.T.; Lu, L.T. Taxa nova Rosacearum sinicarum (Ⅳ). Acta Phytotaxon. Sin. 1982, 20, 295–310. [Google Scholar]
- Naruhashi, N. Rubus L. In Flora of Japan; Iwatsuki, K., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 2001; Volume Ⅱb, pp. 145–169. [Google Scholar]
- Nakai, T. Praecursores ad Floram Sylvaticam Coreanam VII. (Rosaceae). Bot. Mag. 1916, 30, 217–242. [Google Scholar] [CrossRef]
- Nakai, T. Flora Sylvatica Koreana; Pars VII; The government of Chosen: Seoul, Republic of Korea, 1918; pp. 48–77. [Google Scholar]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)—A community resource of plant chromosome numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef]
- Holmgren, P.K.; Holmgren, N.H. Index Herbariorum, A Global Directory of Public Herbaria and Associated Staff [Continuously Updated]; New York Botanical Garden: New York, NY, USA, 1998. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.-F.; Xiong, X.-H.; Boufford, D.E.; Gao, Y.-D.; Xu, B.; Zhang, C. Phylogeny of the Diploid Species of Rubus (Rosaceae). Genes 2023, 14, 1152. https://doi.org/10.3390/genes14061152
Gao X-F, Xiong X-H, Boufford DE, Gao Y-D, Xu B, Zhang C. Phylogeny of the Diploid Species of Rubus (Rosaceae). Genes. 2023; 14(6):1152. https://doi.org/10.3390/genes14061152
Chicago/Turabian StyleGao, Xin-Fen, Xian-Hua Xiong, David E. Boufford, Yun-Dong Gao, Bo Xu, and Cheng Zhang. 2023. "Phylogeny of the Diploid Species of Rubus (Rosaceae)" Genes 14, no. 6: 1152. https://doi.org/10.3390/genes14061152
APA StyleGao, X.-F., Xiong, X.-H., Boufford, D. E., Gao, Y.-D., Xu, B., & Zhang, C. (2023). Phylogeny of the Diploid Species of Rubus (Rosaceae). Genes, 14(6), 1152. https://doi.org/10.3390/genes14061152