
Association of the ACTN3 Gene’s Single-Nucleotide Variant Rs1815739 (R577X) with Sports Qualification and Competitive Distance in Caucasian Athletes of the Southern Urals

 ,
,  ,
,  ,
,  ,
,
Abstract
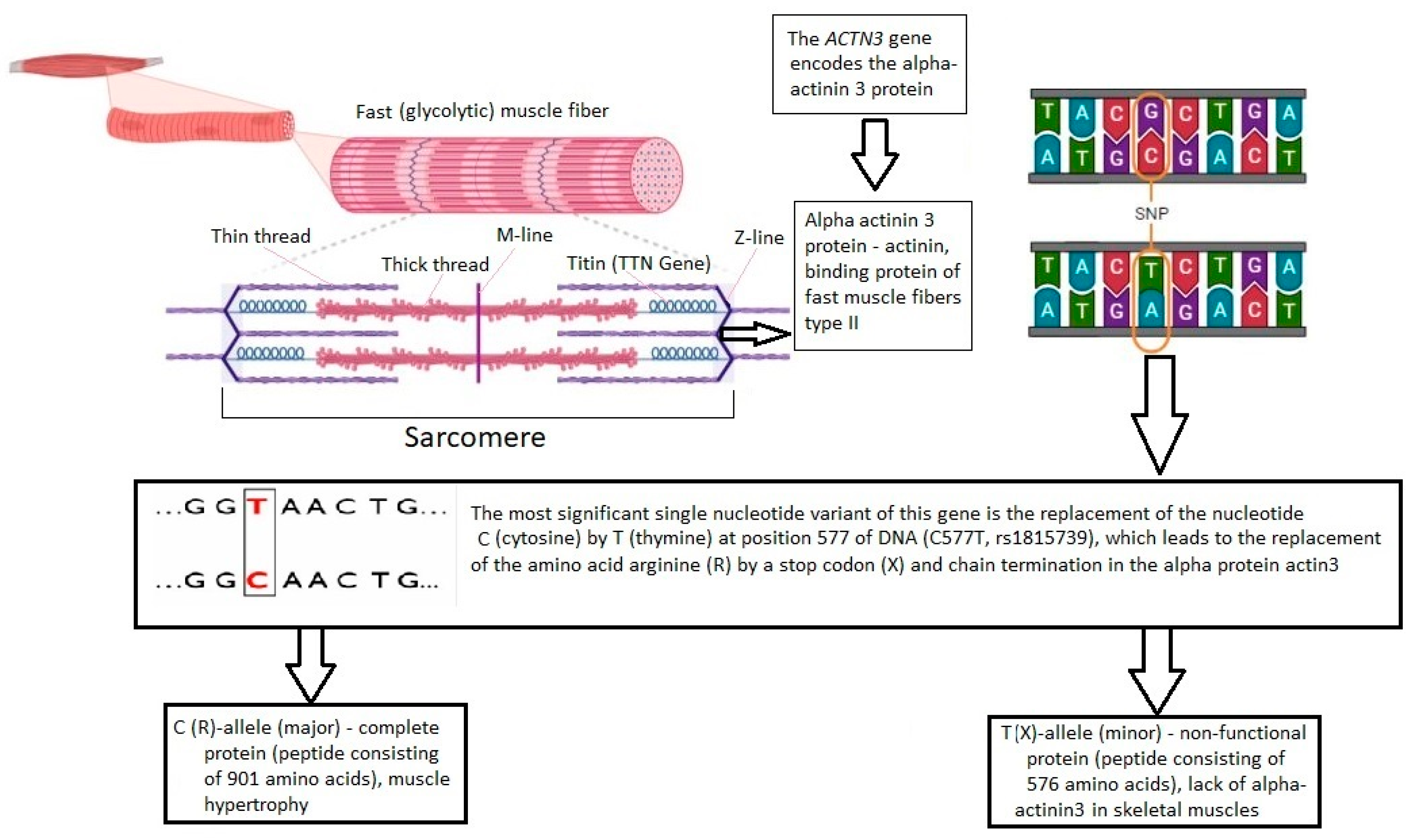
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Participants
2.3. Genetic Analysis
2.4. Body Mass Index Assessment
2.5. Body Component Composition Assessment
2.6. Assessment of Physical Performance
2.7. Statistical Analysis
3. Results
3.1. Baseline Clinical Characteristics
3.2. Distribution of Frequencies of Genotypes and Alleles in Athletes with Different Competitive Distances and Individuals in the Control Group
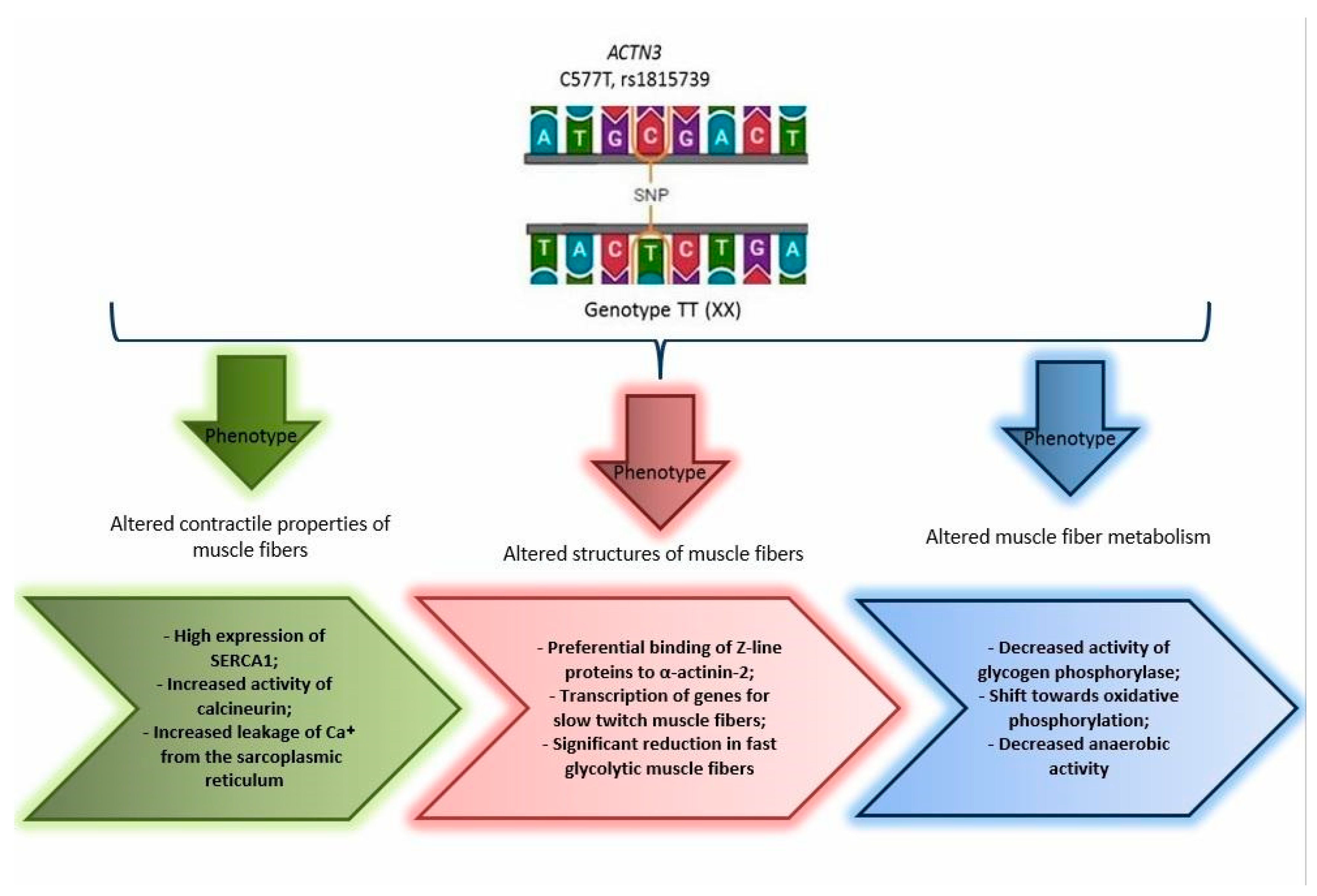
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oreshkina, I.N.; Balberova, O.V.; Bykov, E.V.; Sidorkina, E.G.; Lekontsev, E.V. Construction of training process of skaters in preparatory period based on physical performance. Sci. Notes P.F. Lesgaft Univ. 2019, 2, 256–261. [Google Scholar]
- Balberova, O.V. Candidate genes and single-nucleotide gene variants associated with muscle and tendon injuries in cyclic sports athletes. Pers. Psychiatry Neurol. 2021, 1, 64–72. [Google Scholar] [CrossRef]
- Brendan, E.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar]
- Balberova, O.V.; Bykov, E.V.; Medvedev, G.V. Candidate genes associated with athlete skeletal muscle functions regulation. Pers. Psychiatry Neurol. 2021, 1, 83–94. [Google Scholar] [CrossRef]
- Jacob, Y.; Spiteri, T.; Hart, N.H.; Anderton, R.S. The potential role of genetic markers in talent identification and athlete assessment in elite sport. Sports 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and athletic performance: Aa update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Donnikov, A.E.; Trofimov, D.Y. Actn3 genotype is associated with testosterone levels of athletes. Biol. Sport 2014, 31, 105–108. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Druzhevskaya, A.M.; Lyubaeva, E.V.; Popov, D.V.; Vinogradova, O.L.; Williams, A.G. The dependence of preferred competitive racing distance on muscle fibre type composition and ACTN3 genotype in speed skaters. Exp. Physiol. 2011, 96, 1302–1310. [Google Scholar] [CrossRef]
- Maciejewska-Skrendo, A.; Cięszczyk, P.; Chycki, J.; Sawczuk, M.; Smółka, W. Genetic markers associated with power athlete status. J. Hum. Kinet. 2019, 68, 17–36. [Google Scholar] [CrossRef]
- Garton, F.C.; North, K.N. The Eefect of heterozygosity for the ACTN3 null allele on human muscle performance. Med. Sci. Sports Exerc. 2016, 48, 509–520. [Google Scholar] [CrossRef]
- Baltazar-Martins, G.; Gutiérrez-Hellín, J.; Aguilar-Navarro, M.; Ruiz-Moreno, C.; Moreno-Pérez, V.; López-Samanes, Á.; Domínguez, R.; Del Coso, J. Effect of ACTN3 genotype on sports performance, exercise-induced muscle damage, and injury epidemiology. Sports 2020, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.X.; Houweling, P.J.; North, K.N.; Quinlan, K.G. How does α-actinin-3 deficiency alter muscle function? Mechanistic insights into ACTN3, the ‘gene for speed’. Biochim. Biophys. Acta 2016, 1863, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Houweling, P.J.; Papadimitriou, I.D.; Seto, J.T.; Pérez, L.M.; Coso, J.D.; North, K.N.; Lucia, A.; Eynon, N. Is evolutionary loss our gain? The role of ACTN3 p.Arg577Ter (R577X) genotype in athletic performance, ageing, and disease. Hum. Mutat. 2018, 39, 1774–1787. [Google Scholar] [CrossRef] [PubMed]
- Pilegaard, H.; Saltin, B.; Neufer, P.D. Exercise induces transient transcriptional activation of the PGC-1alpha gene in human skeletal muscle. J Physiol. 2003, 546, 851–858. [Google Scholar] [CrossRef] [PubMed]
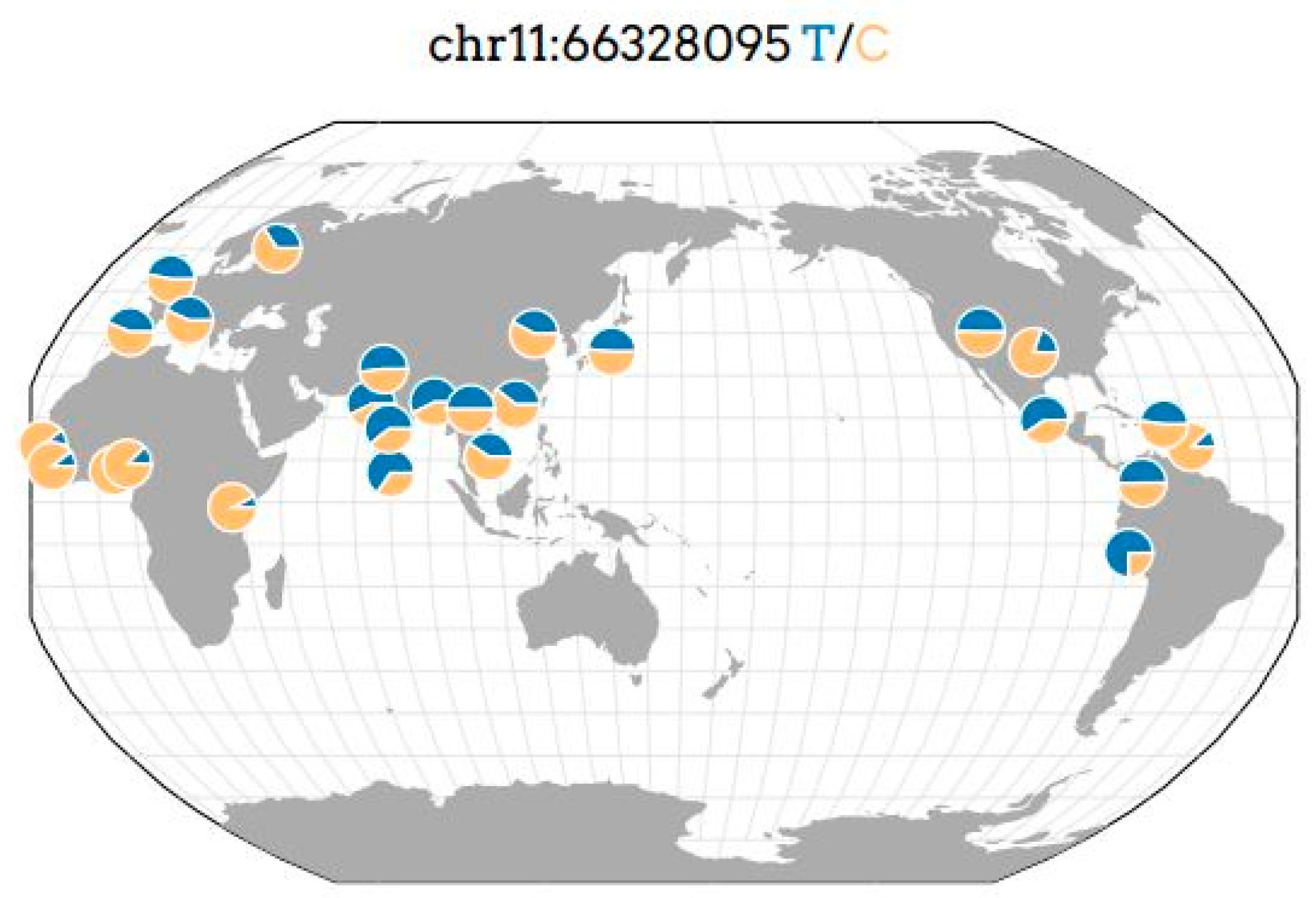
- Geography of Genetic Variants Browser. Available online: http://www.popgen.uchicago.edu/ggv (accessed on 1 September 2022).
- Del Coso, J.; Hiam, D.; Houweling, P.; Pérez, L.M.; Eynon, N.; Lucía, A. More than a ‘speed gene’: ACTN3 R577X genotype, trainability, muscle damage, and the risk for injuries. Eur. J. Appl. Physiol. 2019, 119, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Alfred, T.; Ben-Shlomo, Y.; Cooper, R.; Hardy, R.; Cooper, C.; Deary, I.J.; Gunnell, D.; Harris, S.E.; Kumari, M.; Martin, R.M.; et al. ACTN3 genotype, athletic status, and life course physical capability: Meta-analysis of the published literature and findings from nine studies. Hum. Mutat. 2011, 32, 1008–1018. [Google Scholar] [CrossRef]
- WMA. Available online: https://www.wma.net/ (accessed on 1 September 2022).
- Griffiths, L.J.; Anyim, M.; Doffman, S.R. Comparison of DNA extraction methods for Aspergillus fumigatususing real-time PCR. J. Med. Microbiol. 2006, 55, 1187–1191. [Google Scholar] [CrossRef]
- Kurose, S.; Nishikawa, S.; Nagaoka, T.; Kusaka, M.; Kawamura, J.; Nishioka, Y.; Sato, S.; Tsutsumi, H.; Kimura, Y. Prevalence and risk factors of sarcopenia in community-dwelling older adults visiting regional medical institutions from the Kadoma Sarcopenia Study. Sci. Rep. 2020, 10, 19129. [Google Scholar] [CrossRef]
- Scott, R.A.; Irving, R.; Irwin, L.; Morrison, E.; Charlton, V.; Austin, K.; Tladi, D.; Deason, M.; Headley, S.A.; Kolkhorst, F.W.; et al. ACTN3 and ACE genotypes in elite Jamaican and US sprinters. Med. Sci. Sports Exerc. 2010, 42, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, I.D.; Lucia, A.; Pitsiladis, Y.P.; Pushkarev, V.P.; Dyatlov, D.A.; Orekhov, E.F.; Artioli, G.G.; Guilherme, J.P.; Lancha, A.H., Jr.; Ginevičienė, V.; et al. ACTN3 R577X and ACE I/D gene variants influence performance in elite sprinters: A multi-cohort study. BMC Genom. 2016, 17, 285. [Google Scholar] [CrossRef]
- Silva, M.S.; Bolani, W.; Alves, C.R.; Biagi, D.G.; Lemos, J.R., Jr.; da Silva, J.L.; de Oliveira, P.A.; Alves, G.B.; de Oliveira, E.M.; Negrão, C.E.; et al. Elimination of influences of the ACTN3 R577X variant on oxygen uptake by endurance training in healthy individuals. Int. J. Sports Physiol. Perform. 2015, 10, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.E.; Acuña-Alonzo, V.; Salzano, F.M.; Bortolini, M.C.; Hünemeier, T. Differing evolutionary histories of the ACTN3*R577X polymorphism among the major human geographic groups. PLoS ONE 2015, 10, e0115449. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| General Characteristics | Criteria for Inclusion | Criteria for Exclusion | |
|---|---|---|---|
| Group (Athletes) | Group (Nonathletes) | ||
| Residents | Residents of Southern Urals (the city of Chelyabinsk and the Chelyabinsk region, Russia) | Residents of other regions of Russia | |
| Ethnic group | Caucasians | Not Caucasians | |
| Gender | Male | Female | |
| Age | From 17 to 22 years | Under 17 | |
| Sports qualification | 1st Category, candidate for master of sports (CMS), master of sports (MS), master of sports of international class (MSIC) | No sports qualification | |
| Experience in cyclic sports (speed skating, track and athletics) | 5 years or more | No experience in cyclic sports | |
| General Characteristics | Group 1 (Athletes) (n1 = 76) | Group 2, Control (Nonathletes) (n = 50) | p-Value | |
|---|---|---|---|---|
| SD Subgroup, Short Distances (n1 = 40) | LD Subgroup, Long Distances (n2 = 36) | |||
| Age, years (M ± SD) | 22.1 ± 2.4 | 22.6 ± 2.7 | 21.4 ± 2.7 | >0.05 |
| Height, cm (M ± SD) | 183.0 ± 6.43 | 182.76 ± 5.16 | 176.39 ± 6.37 | >0.05 |
| Weight, kg (M ± SD) | 77.01 ± 9.04 | 70.54 ± 5.11 | 70.12 ± 9.27 | >0.05 |
| BMI according to Quetelet, kg/m2 (M ± SD) | 22.92 ± 1.94 | 21.09 ± 1.28 | 22.52 ± 2.30 | >0.05 |
| General Characteristics | SD Subgroup (Athletes, Short Distances) (n1 = 40) | LD Subgroup (Athletes, Long Distances), (n2 = 36) | p-Value |
|---|---|---|---|
| The value of the muscle component, % (M ± SD) | 50.96 ± 3.71 | 48.21 ± 2.33 | >0.05 |
| The value of the fat component, % (M ± SD) | 8.49 ± 1.34 | 6.98 ± 1.37 | >0.05 |
| Maximum power of the executed load, W (M ± SD) | 292.11 ± 32.79 | 297.50 ± 39.52 | >0.05 |
| Alleles, Genotypes | SD Subgroup, Short Distances (n = 40) | LD Subgroup, Long Distances (n = 36) | Control Group (n = 50) |
|---|---|---|---|
| R | 32 (80.00%) | 43 (59.70%) | 62 (62.00%) |
| X | 8 (20.00%) | 29 (40.30%) | 38 (38.00%) |
| RR | 12 (60.00%) | 11 (30.55%) | 17 (34.00%) |
| RX | 8 (40.00%) | 21 (58.34%) | 28 (56.00%) |
| TT (XX) | 0 (0.00%) | 4 (11.12%) | 5 (10.00%) |
| Alleles, Genotypes | χ2 | p-Value | Odds Ratio | 95% Confidential Interval |
|---|---|---|---|---|
| SD Subgroup vs. Control Group | ||||
| R | 4.20 | 0.04 | 2.45 | 1.02–5.87 |
| X | 0.41 | 0.17–0.98 | ||
| RR | 5.03 | 0.02 | 2.91 | 1.00–8.48 |
| RX | 0.52 | 0.18–1.50 | ||
| XX | 0.20 | 0.01–3.82 | ||
| LD Subgroup vs. Control Group | ||||
| R | 0.33 | 0.57 | 0.83 | 0.45–1.56 |
| X | 1.20 | 0.64–2.24 | ||
| RR | 0.42 | 0.81 | 0.78 | 0.31–1.95 |
| RX | 1.10 | 0.46–2.62 | ||
| XX | 1.14 | 0.33–6.17 | ||
| SD Subgroup vs. LD Subgroup | ||||
| R | 4.78 | 0.03 | 0.37 | 0.15–0.92 |
| X | 2.70 | 1.09–6.68 | ||
| RR | 5.77 | 0.02 | 0.29 | 0.09–0.92 |
| RX | 2.10 | 0.69–6.39 | ||
| XX | 5.68 | 0.29–111.06 | ||
| Alleles, Genotypes | MSIC, MS (n = 14) | CMS (n = 20) | 1st Adult Category (n = 6) |
|---|---|---|---|
| R | 22 (78.57%) | 26 (65.00%) | 8 (66.67%) |
| X | 6 (21,43%) | 14 (35.00%) | 4 (33.33%) |
| RR | 8 (57.14%) | 8 (40.00%) | 2 (33.33%) |
| RX | 6 (42.86%) | 12 (60.00%) | 4 (66.67%) |
| XX | 0 (0.00%) | 0 (0.00%) | 0 (0.00%) |
| Alleles, Genotypes | MSIC, MS (n = 15) | CMS (n = 13) | 1st Adult Category (n = 8) |
|---|---|---|---|
| R | 17 (56.67%) | 16 (61.54%) | 10 (62.50%) |
| X | 13 (43.33%) | 10 (38.46%) | 6 (37.5%) |
| RR | 4 (26.67%) | 4 (30.77%) | 3 (37.5%) |
| RX | 9 (60.00%) | 8 (61.54%) | 4 (50.00%) |
| XX | 2 (13.33%) | 1 (7.69%) | 1 (12.5%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balberova, O.V.; Shnayder, N.A.; Bykov, E.V.; Zakaryukin, Y.E.; Petrova, M.M.; Soloveva, I.A.; Narodova, E.A.; Chumakova, G.A.; Al-Zamil, M.; Asadullin, A.R.; et al. Association of the ACTN3 Gene’s Single-Nucleotide Variant Rs1815739 (R577X) with Sports Qualification and Competitive Distance in Caucasian Athletes of the Southern Urals. Genes 2023, 14, 1512. https://doi.org/10.3390/genes14081512
Balberova OV, Shnayder NA, Bykov EV, Zakaryukin YE, Petrova MM, Soloveva IA, Narodova EA, Chumakova GA, Al-Zamil M, Asadullin AR, et al. Association of the ACTN3 Gene’s Single-Nucleotide Variant Rs1815739 (R577X) with Sports Qualification and Competitive Distance in Caucasian Athletes of the Southern Urals. Genes. 2023; 14(8):1512. https://doi.org/10.3390/genes14081512
Chicago/Turabian StyleBalberova, Olga V., Natalia A. Shnayder, Evgeny V. Bykov, Yuri E. Zakaryukin, Marina M. Petrova, Irina A. Soloveva, Ekaterina A. Narodova, Galina A. Chumakova, Mustafa Al-Zamil, Azat R. Asadullin, and et al. 2023. "Association of the ACTN3 Gene’s Single-Nucleotide Variant Rs1815739 (R577X) with Sports Qualification and Competitive Distance in Caucasian Athletes of the Southern Urals" Genes 14, no. 8: 1512. https://doi.org/10.3390/genes14081512
APA StyleBalberova, O. V., Shnayder, N. A., Bykov, E. V., Zakaryukin, Y. E., Petrova, M. M., Soloveva, I. A., Narodova, E. A., Chumakova, G. A., Al-Zamil, M., Asadullin, A. R., Vaiman, E. E., Trefilova, V. V., & Nasyrova, R. F. (2023). Association of the ACTN3 Gene’s Single-Nucleotide Variant Rs1815739 (R577X) with Sports Qualification and Competitive Distance in Caucasian Athletes of the Southern Urals. Genes, 14(8), 1512. https://doi.org/10.3390/genes14081512