
Tomato-Thaumatin-like Protein Genes Solyc08g080660 and Solyc08g080670 Confer Resistance to Five Soil-Borne Diseases by Enhancing β-1,3-Glucanase Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Bacterial Infection
2.2. Identification of Disease Resistance in Tomato Leaves via RT-qPCR
2.3. Generation of Overexpressing Transgenic Tomato Plants
2.4. Generation of Gene-Edited Tomato Plants
2.5. Disease Resistance of Transgenic Plants
2.6. Determination of ROS Antioxidant Physiological Indexes in Transgenic Plants
2.7. Determination of β-1,3-GA Activity
2.8. Data Statistics
3. Results
3.1. Response of the SlTLP5 and SlTLP6 Genes in Five Soil-Borne Diseases
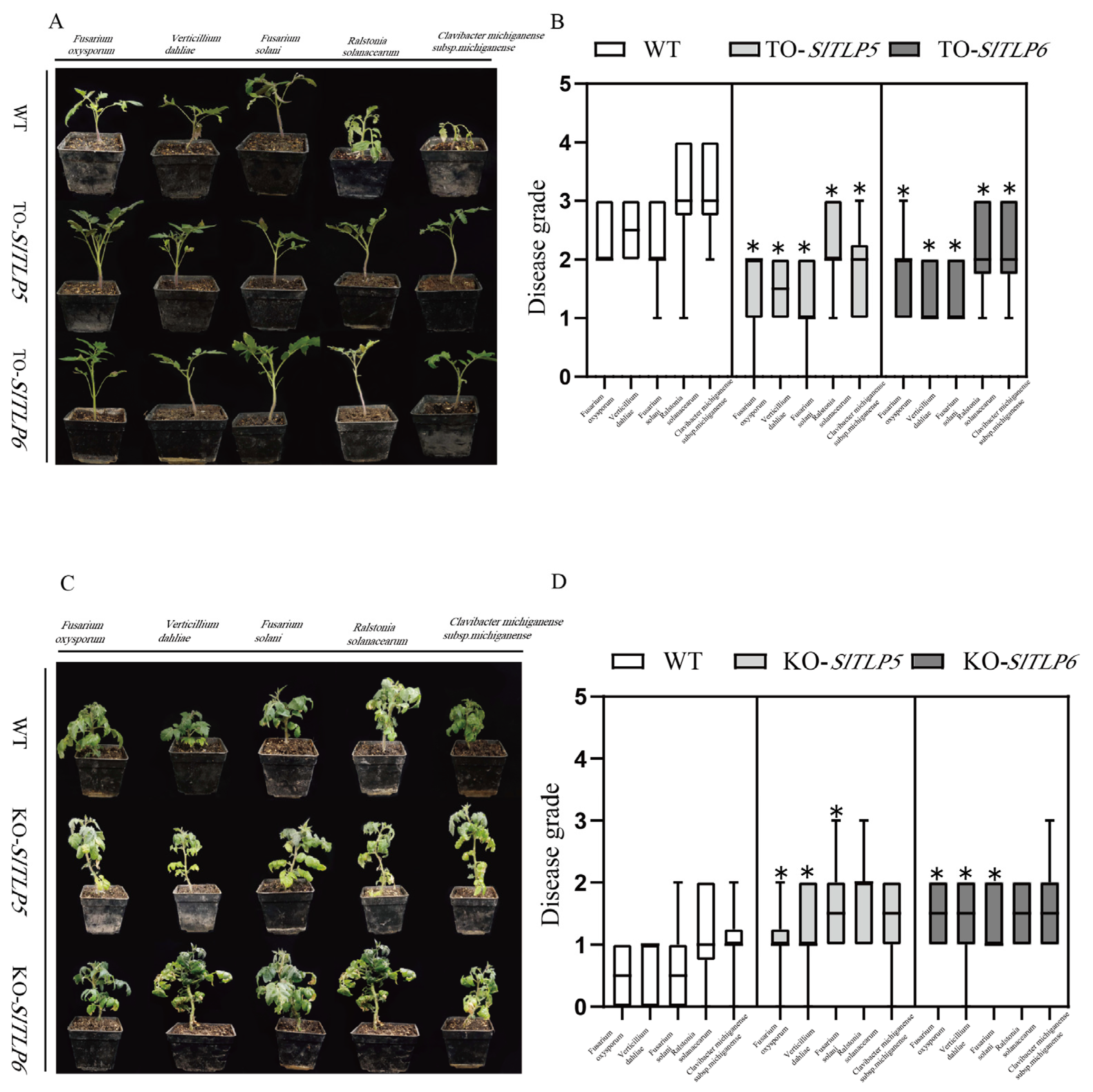
3.2. Overexpression of TLP Conferred Increased Disease Resistance to Tomato Plants
3.3. TLP Knockout Reduces Disease Resistance in Tomato Plants
3.4. Physiological Changes in Resistance of Overexpressing Plants of Susceptible Cultivars
3.5. Physiological Changes in Resistance of Knockout Plants of Disease-Resistant Cultivars
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Loon, L.; Rep, M.; Pieterse, C. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narusaka, Y.; Narusaka, M.; Abe, H.; Hosaka, N.; Kobayashi, M.; Shiraishi, T.; Iwabuchi, M. High-throughput screening for plant defense activators using a β-glucuronidase-reporter gene assay in Arabidopsis thaliana. Plant Biotechnol. 2009, 26, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, L.; Zheng, Z.; Sun, Y.; Zhou, L.; Yang, Y.; Cheng, F.; Zhang, Z.; Wang, X.; Huang, S.; et al. Fine mapping of the Ph-3 gene conferring resistance to late blight (Phytophthora infestans) in tomato. Theor. Appl. Genet. 2013, 126, 2643–2653. [Google Scholar] [CrossRef]
- Rajam, M.V.; Chandola, N.; Saiprasad Goud, P.; Singh, D.; Kashyap, V.; Choudhary, M.L.; Sihachakr, D. Thaumatin gene confers resistance to fungal pathogens as well as tolerance to abiotic stresses in transgenic tobacco plants. Biol. Plant. 2007, 51, 135–141. [Google Scholar] [CrossRef]
- Abad, L.R.; D’Urzo, M.P.; Liu, D.; Narasimhan, M.L.; Reuveni, M.; Zhu, J.K.; Niu, X.; Singh, N.K.; Hasegawa, P.M.; Bressan, R.A. Antifungal activity of tobacco osmotin has specificity and involves plasma membrane permeabilization. Plant Sci. 1996, 118, 11–23. [Google Scholar] [CrossRef]
- Radhajeyalakshmi, R.; Velazhahan, R.; Balasubramanian, P.; Doraiswamy, S. Overexpression of thaumatin-like protein in transgenic tomato plants confers enhanced resistance to Alternaria solani. Arch. Phytopathol. Plant Prot. 2005, 38, 257–265. [Google Scholar] [CrossRef]
- Aghazadeh, R.; Zamani, M.; Motallebi, M.; Moradyar, M. Agrobacterium-Mediated Transformation of the Oryza sativa Thaumatin-Like Protein to Canola (R Line Hyola308) for Enhancing Resistance to Sclerotinia sclerotiorum. Iran. J. Biotechnol. 2017, 15, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Qiao, H.; Zhang, X.; Guo, C.; Wang, M.; Wang, Y.; Wang, X. Analysis of the grape (Vitis vinifera L.) thaumatin-like protein (TLP) gene family and demonstration that TLP29 contributes to disease resistance. Sci. Rep. 2017, 7, 4269. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Basu, A.; Kundu, S. Overexpression of a New Osmotin-Like Protein Gene (SindOLP) Confers Tolerance against Biotic and Abiotic Stresses in Sesame. Front. Plant Sci. 2017, 8, 410. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; Sturrock, R.; Ekramoddoullah, A.K.M. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef]
- Wang, X.; Tang, C.; Deng, L.; Cai, G.; Liu, X.; Liu, B.; Han, Q.; Buchenauer, H.; Wei, G.; Han, D.; et al. Characterization of a pathogenesis-related thaumatin-like protein gene TaPR5 from wheat induced by stripe rust fungus. Physiol. Plant. 2010, 139, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-G.; Fukumoto, T.; Tatano, S.; Gomi, K.; Ohtani, K.; Tada, Y.; Akimitsu, K. Molecular cloning and characterization of a thaumatin-like protein-encoding cDNA from rough lemon. Physiol. Mol. Plant Pathol. 2009, 74, 3–10. [Google Scholar] [CrossRef]
- Kim, D.H.; Noh, M.Y.; Park, K.B.; Jo, Y.H. Expression profiles of two thaumatin-like protein (TmTLP) genes in responses to various micro-organisms from Tenebrio molitor. Entomol. Res. 2017, 47, 35–40. [Google Scholar] [CrossRef]
- Beatrice, C.; Linthorst, J.M.H.; Cinzia, F.; Luca, R. Enhancement of PR1 and PR5 gene expressions by chitosan treatment in kiwifruit plants inoculated with Pseudomonas syringae pv. actinidiae. Eur. J. Plant Pathol. 2016, 148, 163–179. [Google Scholar] [CrossRef]
- Rout, E.; Nanda, S.; Joshi, R.K. Molecular characterization and heterologous expression of a pathogen induced PR5 gene from garlic (Allium sativum L.) conferring enhanced resistance to necrotrophic fungi. Eur. J. Plant Pathol. 2016, 144, 345–360. [Google Scholar] [CrossRef]
- Kalpana, K.; Maruthasalam, S.; Rajesh, T.; Poovannan, K.; Kumar, K.K.; Kokiladevi, E.; Raja, J.A.; Sudhakar, D.; Velazhahan, R.; Samiyappan, R.; et al. Engineering sheath blight resistance in elite indica rice cultivars using genes encoding defense proteins. Plant Sci. 2006, 170, 203–215. [Google Scholar] [CrossRef]
- De A Campos, M.; Silva, M.S.; Magalhães, C.P.; Ribeiro, S.G.; Sarto, R.P.; Vieira, E.A.; de Sá, M.F.G. Expression in Escherichia coli, purification, refolding and antifungal activity of an osmotin from Solanum nigrum. Microb. Cell Factories 2008, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Deng, M.; Yang, Z.; Mao, L.; Jiang, S.; Yue, Y.; Zhao, K. Two Tomato (Solanum lycopersicum) Thaumatin-Like Protein Genes Confer Enhanced Resistance to Late Blight (Phytophthora infestans). Phytopathology 2021, 111, 1790–1799. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Meng, J.; Luan, Y. A Tomato Nucleotide Binding Sites—Leucine-Rich Repeat Gene Is Positively Involved in Plant Resistance to Phytophthora infestans. Phytopathology 2018, 108, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Shen, X.; Yuan, H.; Liu, Y.; Liao, X.; Wang, Q.; Liu, L.; Li, F.; Li, T. Isolation and Characterization of Dehydration-Responsive Element-Binding Factor 2C (MsDREB2C) from Malus sieversii Roem. Plant Cell Physiol. 2013, 54, 1415–1430. [Google Scholar] [CrossRef] [Green Version]
- Løvdal, T.; Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem. 2009, 387, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Cui, J.; Liu, Z.; Luan, Y. SpWRKY6 acts as a positive regulator during tomato resistance to Phytophthora infestans infection. Biochem. Biophys. Res. Commun. 2018, 506, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.; Lin, Y.R.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, B.; Vasse, J.; Le-Courtois, V.; Trigalet-Démery, D.; López, M.M.; Trigalet, A. Comparative Behavior of Ralstonia solanacearum Biovar 2 in Diverse Plant Species. Phytopathology 2008, 98, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastià, P.; de Pedro-Jové, R.; Daubech, B.; Kashyap, A.; Coll, N.S.; Valls, M. The Bacterial Wilt Reservoir Host Solanum dulcamara Shows Resistance to Ralstonia solanacearum Infection. Front. Plant Sci. 2021, 12, 755708. [Google Scholar] [CrossRef]
- Ji, H.; Mao, H.; Li, S.; Feng, T.; Zhang, Z.; Cheng, L.; Luo, S.; Borkovich, K.A.; Ouyang, S. Fol-milR1, a pathogenicity factor of Fusarium oxysporum, confers tomato wilt disease resistance by impairing host immune responses. New Phytol. 2021, 232, 705–718. [Google Scholar] [CrossRef]
- Validov, S.Z.; Kamilova, F.D.; Lugtenberg, B.J.J. Monitoring of pathogenic and non-pathogenic Fusarium oxysporum strains during tomato plant infection. Microb. Biotechnol. 2011, 4, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Çakır, B.; Gül, A.; Yolageldi, L.; Özaktan, H. Response to Fusarium oxysporum f.sp. radicis-lycopersici in tomato roots involves regulation of SA- and ET-responsive gene expressions. Eur. J. Plant Pathol. 2014, 139, 379–391. [Google Scholar] [CrossRef]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncRNA16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Jiang, N.; Zhou, X.; Hou, X.; Yang, G.; Meng, J.; Luan, Y. Tomato MYB49 enhances resistance to Phytophthora infestans and tolerance to water deficit and salt stress. Planta 2018, 248, 1487–1503. [Google Scholar] [CrossRef]
- Zong, Y.; Liu, J.; Li, B.; Qin, G.; Tian, S. Effects of yeast antagonists in combination with hot water treatment on postharvest diseases of tomato fruit. Biol. Control 2010, 54, 316–321. [Google Scholar] [CrossRef]
- De Jesús-Pires, C.; Ferreira-Neto, J.R.; Pacifico Bezerra-Neto, J.; Kido, E.A.; de Oliveira Silva, R.L.; Pandolfi, V.; Wanderley-Nogueira, A.C.; Binneck, E.; da Costa, A.F.; Pio-Ribeiro, G.; et al. Plant Thaumatin-like Proteins: Function, Evolution and Biotechnological Applications. Curr. Protein Pept. Sci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, H.; Wei, X.; Zhang, J.; Wang, H.; Liu, D. Expression analysis and functional characterization of a pathogen-induced thaumatin-like gene in wheat conferring enhanced resistance to Puccinia triticina. J. Plant Interact. 2017, 12, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Ojola, P.O.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of rice thaumatin-like protein (Ostlp) gene in transgenic cassava results in enhanced tolerance to Colletotrichum gloeosporioides f. sp. manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, I.; Vera, P.; Frank, R.; Conejero, V. Identification of the viroid-induced tomato pathogenesis-related (PR) protein P23 as the thaumatin-like tomato protein NP24 associated with osmotic stress. Plant Mol. Biol. 1991, 16, 931–934. [Google Scholar] [CrossRef]
- He, L.; Li, L.; Zhu, Y.; Pan, Y.; Zhang, X.; Han, X.; Li, M.; Chen, C.; Li, H.; Wang, C. BolTLP1, a Thaumatin-like Protein Gene, Confers Tolerance to Salt and Drought Stresses in Broccoli (Brassica oleracea L. var. Italica). Int. J. Mol. Sci. 2021, 22, 11132. [Google Scholar] [CrossRef]
- Futamura, N.; Tani, N.; Tsumura, Y.; Nakajima, N.; Sakaguchi, M.; Shinohara, K. Characterization of genes for novel thaumatin-like proteins in Cryptomeria japonica. Tree Physiol. 2006, 26, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, F.; Zhang, X.; Zhang, Y.; Hou, Y.; Zhang, S.; Wu, Z. Purification and Characterization of a CkTLP Protein from Cynanchum komarovii Seeds that Confers Antifungal Activity. PLoS ONE 2011, 6, e16930. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, C.; Takezawa, D.; Shimada, T.; Hamada, T.; Fujikawa, S.; Arakawa, K. Abscisic acid- and cold-induced thaumatin-like protein in winter wheat has an antifungal activity against snow mold, Microdochium nivale. Physiol. Plant. 2002, 115, 101–110. [Google Scholar] [CrossRef]
- Chen, W.P.; Chen, P.D.; Liu, D.J.; Kynast, R.; Friebe, B.; Velazhahan, R.; Muthukrishnan, S.; Gill, B.S. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor. Appl. Genet. 1999, 99, 755–760. [Google Scholar] [CrossRef]
- Liu, D.; Raghothama, K.G.; Hasegawa, P.M.; Bressan, R.A. Osmotin overexpression in potato delays development of disease symptoms. Proc. Natl. Acad. Sci. USA 1994, 91, 1888–1892. [Google Scholar] [CrossRef]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, M.L.; Damsz, B.; Coca, M.A.; Ibeas, J.I.; Yun, D.-J.; Pardo, J.M.; Hasegawa, P.M.; Bressan, R.A. A Plant Defense Response Effector Induces Microbial Apoptosis. Mol. Cell 2001, 8, 921–930, Erratum in Mol. Cell 2001, 8, 1153. [Google Scholar] [CrossRef]
- Anand, A.; Lei, Z.; Sumner, L.W.; Mysore, K.S.; Arakane, Y.; Bockus, W.W.; Muthukrishnan, S. Apoplastic extracts from a transgenic wheat line exhibiting lesion-mimic phenotype have multiple pathogenesis-related proteins that are antifungal. Mol. Plant-Microbe Interact. 2004, 17, 1306–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenier, J.; Potvin, C.; Asselin, A. Barley Pathogenesis-Related Proteins with Fungal Cell Wall Lytic Activity Inhibit the Growth of Yeasts. Plant Physiol. 1993, 103, 1277–1283. [Google Scholar] [CrossRef]
- Osmond, R.I.; Hrmova, M.; Fontaine, F.; Imberty, A.; Fincher, G.B. Binding interactions between barley thaumatin-like proteins and (1,3)-β-D-glucans: Kinetics, specificity, structural analysis and biological implications. Eur. J. Biochem. 2001, 268, 4190–4199. [Google Scholar] [CrossRef]
- El-Kereamy, A.; El-Sharkawy, I.; Ramamoorthy, R.; Taheri, A.; Errampalli, D.; Kumar, P.; Jayasankar, S. Prunus domestica Pathogenesis-Related Protein-5 Activates the Defense Response Pathway and Enhances the Resistance to Fungal Infection. PLoS ONE 2011, 6, e17973. [Google Scholar] [CrossRef] [Green Version]
- Theis, T.; Stahl, U. Antifungal proteins: Targets, mechanisms and prospective applications. Cell. Mol. Life Sci. 2004, 61, 437–455. [Google Scholar] [CrossRef]
- Chand, S.K.; Nanda, S.; Mishra, R.; Joshi, R.K. Multiple garlic (Allium sativum L.) microRNAs regulate the immunity against the basal rot fungus Fusarium oxysporum f. sp. Cepae. Plant Sci. 2017, 257, 9–21. [Google Scholar] [CrossRef]
- Anisimova, O.K.; Shchennikova, A.V.; Kochieva, E.Z.; Filyushin, M.A. Pathogenesis-Related Genes of PR1, PR2, PR4, and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021, 22, 6688. [Google Scholar] [CrossRef] [PubMed]
- Filyushin, M.A.; Anisimova, O.K.; Kochieva, E.Z.; Shchennikova, A.V. Genome-Wide Identification and Expression of Chitinase Class I Genes in Garlic (Allium sativum L.) Cultivars Resistant and Susceptible to Fusarium proliferatum. Plants 2021, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.; Zhu, W.; Li, W.; Wang, F. Transcriptome Analysis Reveals the Effects of Chinese Chive (Allium tuberosum R.) Extract on Fusarium oxysporum f. sp. radicis-lycopersici Spore Germination. Curr. Microbiol. 2020, 77, 855–864. [Google Scholar] [CrossRef]
- Jayasankar, S.; Li, Z.; Gray, D.J. In-vitro selection of Vitis vinifera ‘Chardonnay’ with Elsinoe ampelina culture filtrate is accompanied by fungal resistance and enhanced secretion of chitinase. Planta 2000, 211, 200–208. [Google Scholar] [CrossRef]
- Schickler, H.; Chet, I. Heterologous chitinase gene expression to improve plant defense against phytopathogenic fungi. J. Ind. Microbiol. Biotechnol. 1997, 19, 196–201. [Google Scholar] [CrossRef]
- Singh, N.K.; Kumar, K.R.R.; Kumar, D.; Shukla, P.; Kirti, P.B. Characterization of a Pathogen Induced Thaumatin-Like Protein Gene AdTLP from Arachis diogoi, a Wild Peanut. PLoS ONE 2013, 8, e83963. [Google Scholar] [CrossRef]
- Di Matteo, A.; Federici, L.; Mattei, B.; Salvi, G.; Johnson, K.A.; Savino, C.; De Lorenzo, G.; Tsernoglou, D.; Cervone, F. The crystal structure of polygalacturonase-inhibiting protein (PGIP), a leucine-rich repeat protein involved in plant defense. Proc. Natl. Acad. Sci. USA 2003, 100, 10124–10128. [Google Scholar] [CrossRef]
- Blein, J.-P.; Coutos-Thévenot, P.; Marion, D.; Ponchet, M. From elicitins to lipid-transfer proteins: A new insight in cell signalling involved in plant defence mechanisms. Trends Plant Sci. 2002, 7, 293–296. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Xu, B.; Xu, J.; Li, Z.; Jiang, C.; Zhou, Y.; Yang, Z.; Deng, M.; Lv, J.; Zhao, K. Tomato-Thaumatin-like Protein Genes Solyc08g080660 and Solyc08g080670 Confer Resistance to Five Soil-Borne Diseases by Enhancing β-1,3-Glucanase Activity. Genes 2023, 14, 1622. https://doi.org/10.3390/genes14081622
Li X, Xu B, Xu J, Li Z, Jiang C, Zhou Y, Yang Z, Deng M, Lv J, Zhao K. Tomato-Thaumatin-like Protein Genes Solyc08g080660 and Solyc08g080670 Confer Resistance to Five Soil-Borne Diseases by Enhancing β-1,3-Glucanase Activity. Genes. 2023; 14(8):1622. https://doi.org/10.3390/genes14081622
Chicago/Turabian StyleLi, Xinyun, Bin Xu, Junqiang Xu, Zuosen Li, Caiqian Jiang, Ying Zhou, Zhengan Yang, Minghua Deng, Junheng Lv, and Kai Zhao. 2023. "Tomato-Thaumatin-like Protein Genes Solyc08g080660 and Solyc08g080670 Confer Resistance to Five Soil-Borne Diseases by Enhancing β-1,3-Glucanase Activity" Genes 14, no. 8: 1622. https://doi.org/10.3390/genes14081622