
Molecular Characteristics of Bean Common Mosaic Virus Occurring in Inner Mongolia, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Source
2.2. Amplification of the Genomic Fragments of BCMV
2.3. Cloning and Sequencing of the BCMV Fragments
2.4. BCMV Genome Assembly and Sequence Analysis
2.5. BCMV Host Range
3. Results
3.1. Genome Structure and Sequence Identity Analysis of BCMV Isolates
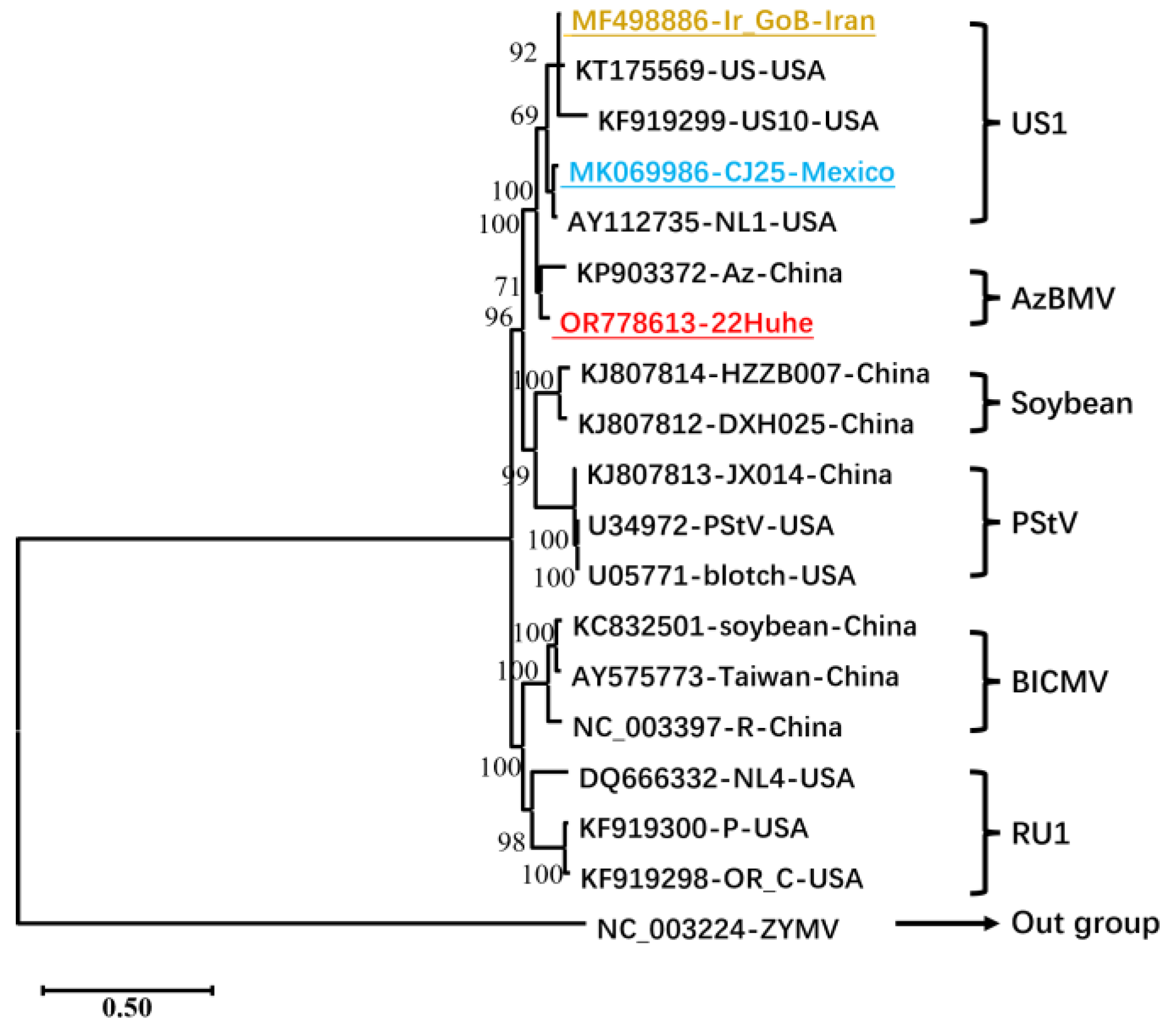
3.2. Phylogeny and Recombination Analyses of BCMV Isolates
3.3. BCMV Host Range and Symptoms
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adams, M.J.; Zerbini, F.M.; French, R.; Rabenstein, F.; Stenger, D.C.; Valkonen, J.P.T. Family Potyviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses. Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: Cambridge, MA, USA, 2012; pp. 1069–1089. [Google Scholar]
- Li, Y.; Liu, Z.; Yang, K.; Li, Y.; Zhao, B.; Fan, Z.; Wan, P. First report of Bean common mosaic virus infecting Azuki bean (Vigna angularis) in China. Plant Dis. 2014, 98, 1017. [Google Scholar] [CrossRef] [PubMed]
- Damayanti, T.A.; Susilo, D.; Nurlaelah, S.; Sartiami, D.; Okuno, T.; Mise, K. First report of Bean common mosaic virus in yam bean [Pachyrhizus erosus (L.) Urban] in Indonesia. J. Gen. Plant Pathol. 2008, 74, 438–442. [Google Scholar] [CrossRef]
- Singh, S.P.; Schwartz, H.F. Breeding common bean for resistance to diseases: A review. Crop Sci. 2010, 50, 2199–2223. [Google Scholar] [CrossRef]
- Zhou, G.-C.; Wu, X.-Y.; Zhang, Y.-M.; Wu, P.; Wu, X.-Z.; Liu, L.-W.; Wang, Q.; Hang, Y.-Y.; Yang, J.-Y.; Shao, Z.-Q. A genomic survey of thirty soybean-infecting bean common mosaic virus (BCMV) isolates from China pointed BCMV as a potential threat to soybean production. Virus Res. 2014, 191, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.A.; Wamonje, F.O.; Mukeshimana, G.; Harvey, J.J.; Carr, J.P.; Mitter, N. Bean common mosaic virus and Bean common mosaic necrosis virus: Relationships, biology, and prospects for control. Adv. Virus Res. 2015, 93, 1–46. [Google Scholar] [PubMed]
- Ekpo, E.; Saettler, A. Distribution pattern of bean common mosaic virus in developing bean seed. Phytopathology 1974, 64, 269–270. [Google Scholar] [CrossRef]
- Tang, M.; Feng, X. Bean common mosaic disease: Etiology, resistance resource, and future prospects. Agronomy 2022, 13, 58. [Google Scholar] [CrossRef]
- Feng, X.; Orellana, G.E.; Green, J.C.; Melzer, M.J.; Hu, J.S.; Karasev, A.V. A New Strain of Bean Common Mosaic Virus From Lima Bean (Phaseolus lunatus): Biological and Molecular Characterization. Plant Dis. 2019, 103, 1220–1227. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Feng, X.; Poplawsky, A.R.; Nikolaeva, O.V.; Myers, J.R.; Karasev, A. Recombinants of Bean common mosaic virus (BCMV) and genetic determinants of BCMV involved in overcoming resistance in common bean. Phytopathology 2014, 104, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Naderpour, M.; Lund, O.S.; Johansen, I.E. Sequence analysis of expressed cDNA of Bean common mosaic virus RU1 isolate. Iran. J. Virol. 2009, 3, 39–41. [Google Scholar] [CrossRef]
- Gunasinghe, U.; Flasinski, S.; Nelson, R.; Cassidy, B. Nucleotide sequence and genome organization of peanut stripe potyvirus. J. Gen. Virol. 1994, 75, 2519–2525. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fang, C. Molecular sequencing and analysis of the viral genomic regions of Blackeye cowpea mosaic virus Taiwan strain. Plant Pathol. Bull. 2004, 13, 117–126. [Google Scholar]
- Li, Y.; Cao, Y.; Fan, Z.; Wan, P. Identification of a naturally occurring Bean common mosaic virus recombinant isolate infecting azuki bean. J. Plant Pathol. 2016, 98, 129–133. [Google Scholar]
- Berger, P.; Wyatt, S.; Shiel, P.; Silbernagel, M.; Druffel, K.; Mink, G. Phylogenetic analysis of the Potyviridae with emphasis on legume-infecting potyviruses. Arch. Virol. 1997, 142, 1979–1999. [Google Scholar] [CrossRef]
- Cui, X.; Shen, L.; Yuan, X.; Gu, H.; Chen, X. First report of Bean common mosaic virus infecting mungbean (Vigna radiata) in China. Plant Dis. 2014, 98, 1590. [Google Scholar] [CrossRef]
- Han, T.; Yang, C.; Fu, J.; Liao, Y.; Lyu, S.; Jia, R.; Shi, C. Identification and sequence analysis of Bean common mosaic virus in Liaoning. Acta Agric. Univ. Jiangxiensis 2018, 40, 378–382. [Google Scholar]
- Chen, Z.; Yang, L.; Chen, L.; Sun, Y.; Wei, Z.; Li, Y.; Zhao, M.; Wen, G. Detection and identification of BCMV infecting Polygonatum kingianum. Curr. Biotechnol. 2019, 9, 122–128. [Google Scholar]
- Su, X.; Zhou, X.; Li, Y.; Ma, L.; Cheng, X.; Guo, K. First Report of Bean common mosaic virus Infecting Heavenly Bamboo (Nandina domestica) in China. Plant Dis. 2022, 106, 1079. [Google Scholar] [CrossRef]
- Zhao, Z.; Dong, Z.; Wang, S.; Fan, Q.; Zhang, Y. First report of bean common mosaic virus naturally infecting rice bean (Vigna umbellata) in China. Plant Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Yan, D.; Zheng, H.; Ji, M.; Wu, X.; Qi, R.; Yan, F. First report of bean common mosaic virus naturally infecting yam bean (Pachyrhizus erosus) in China. Plant Dis. 2023, 107, 238. [Google Scholar] [CrossRef] [PubMed]
- García-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Chare, E.; Holmes, E. A phylogenetic survey of recombination frequency in plant RNA viruses. Arch. Virol. 2006, 151, 933–946. [Google Scholar] [CrossRef]
- Sokhandan-Bashir, N.; Melcher, U. Population genetic analysis of grapevine fanleaf virus. Arch. Virol. 2012, 157, 1919–1929. [Google Scholar] [CrossRef]
- Mihálik, D.; Grešíková, S.; Hančinský, R.; Cejnar, P.; Havrlentová, M.; Kraic, J. A Novel Isolate of Bean Common Mosaic Virus Isolated from Crownvetch (Securigera varia L. Lassen). Agronomy 2023, 13, 1677. [Google Scholar] [CrossRef]
- Moradi, Z.; Mehrvar, M. Genetic variability and molecular evolution of Bean common mosaic virus populations in Iran: Comparison with the populations in the world. Eur. J. Plant Pathol. 2019, 154, 673–690. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′-3′) | Tm/°C | Amplicon Size/bp | Primer Location/bp |
|---|---|---|---|---|
| BCMV-F1 | CTAACTGTGGTCGGTTCACCCA | 60 | 3870 | 253–4060 |
| BCMV-R1 | CCCTACTTGGTGGTGGTTGGA | 60 | 253–4060 | |
| BCMV-F2 | ATGAACTTTCCACAGCTTCTGTAGTGAG | 60 | 3942 | 5875–9817 |
| BCMV-R2 | TTATGGAGAGCATCACTGTAGGGTGC | 62 | 5875–9817 | |
| BCMV-F3 | TATAAAGTTTCTCGTCTTCCTTCCCAT | 58 | 2812 | 3807–6619 |
| BCMV-R3 | CAACTGCGAAGTCACACCTCAGAAG | 61 | 3807–6619 | |
| 5′ RACE | GCTTTTGATTTCTATGAGGTGA | 57 | 9252 | 610–9862 |
| 3′ RACE | CTAGGTTTGAAGAGGAGGTGCG | 59 | 610–9862 | |
| BCMV-F | ACAACACACTCATGGTTGTAAT | 60 | 490 | 1256–1746 |
| BCMV-R | CTGATTCTGCTATGTACGGAGC | 66 | 1256–1746 |
| Methods | p-Value |
|---|---|
| RDP | 9.216 × 10−10 |
| GENECONV | 8.188 × 10−11 |
| BlootScan | 6.100 × 10−20 |
| MaxChi | 1.347 × 10−18 |
| Chimaera | 9.245 × 10−7 |
| SiScan | 1.803 × 10−16 |
| 3Seq | 7.209 × 10−8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Li, Z.; Wu, Z.; Sun, Y.; Niu, S.; Guo, M.; Zhang, L. Molecular Characteristics of Bean Common Mosaic Virus Occurring in Inner Mongolia, China. Genes 2024, 15, 133. https://doi.org/10.3390/genes15010133
Li J, Li Z, Wu Z, Sun Y, Niu S, Guo M, Zhang L. Molecular Characteristics of Bean Common Mosaic Virus Occurring in Inner Mongolia, China. Genes. 2024; 15(1):133. https://doi.org/10.3390/genes15010133
Chicago/Turabian StyleLi, Jingru, Zhengnan Li, Zhanmin Wu, Yu Sun, Suqing Niu, Mengze Guo, and Lei Zhang. 2024. "Molecular Characteristics of Bean Common Mosaic Virus Occurring in Inner Mongolia, China" Genes 15, no. 1: 133. https://doi.org/10.3390/genes15010133
APA StyleLi, J., Li, Z., Wu, Z., Sun, Y., Niu, S., Guo, M., & Zhang, L. (2024). Molecular Characteristics of Bean Common Mosaic Virus Occurring in Inner Mongolia, China. Genes, 15(1), 133. https://doi.org/10.3390/genes15010133