
Chronic Adolescent Restraint Stress Downregulates miRNA-200a Expression in Male and Female C57BL/6J and BALB/cJ Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Restraint Stress and Body Weight
2.3. Blood Collection and Corticosterone Assay
2.4. Brain Extraction, RNA Isolation, and qPCR
2.5. Bioinformatics Analysis
2.6. Statistics
3. Results
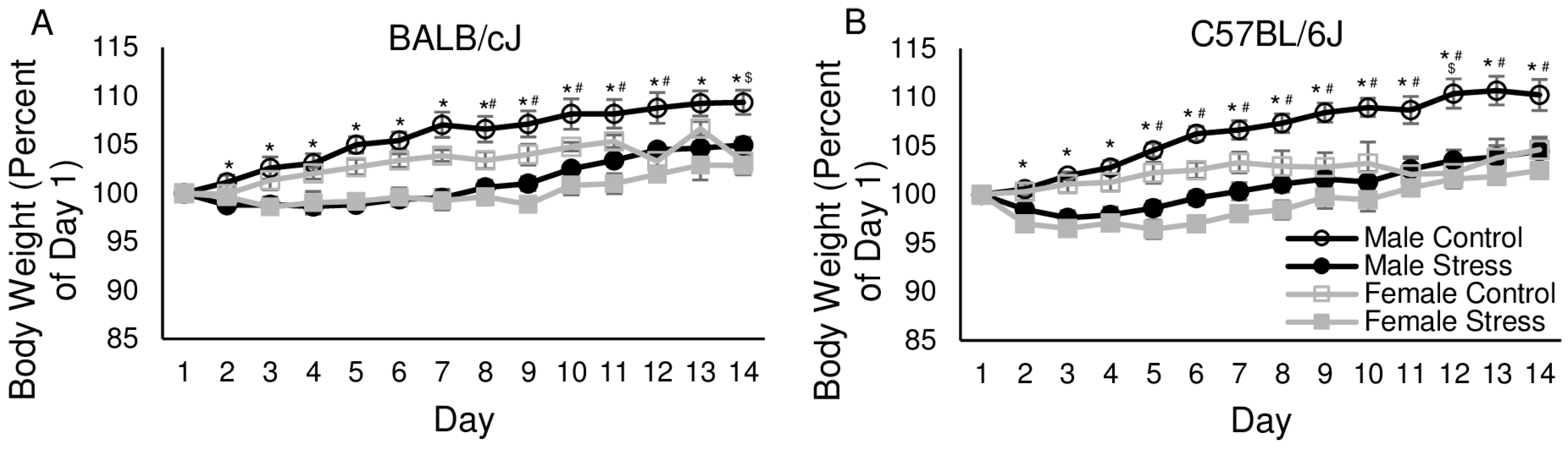
3.1. Body Weight
3.1.1. Day 1
3.1.2. Days 2–14
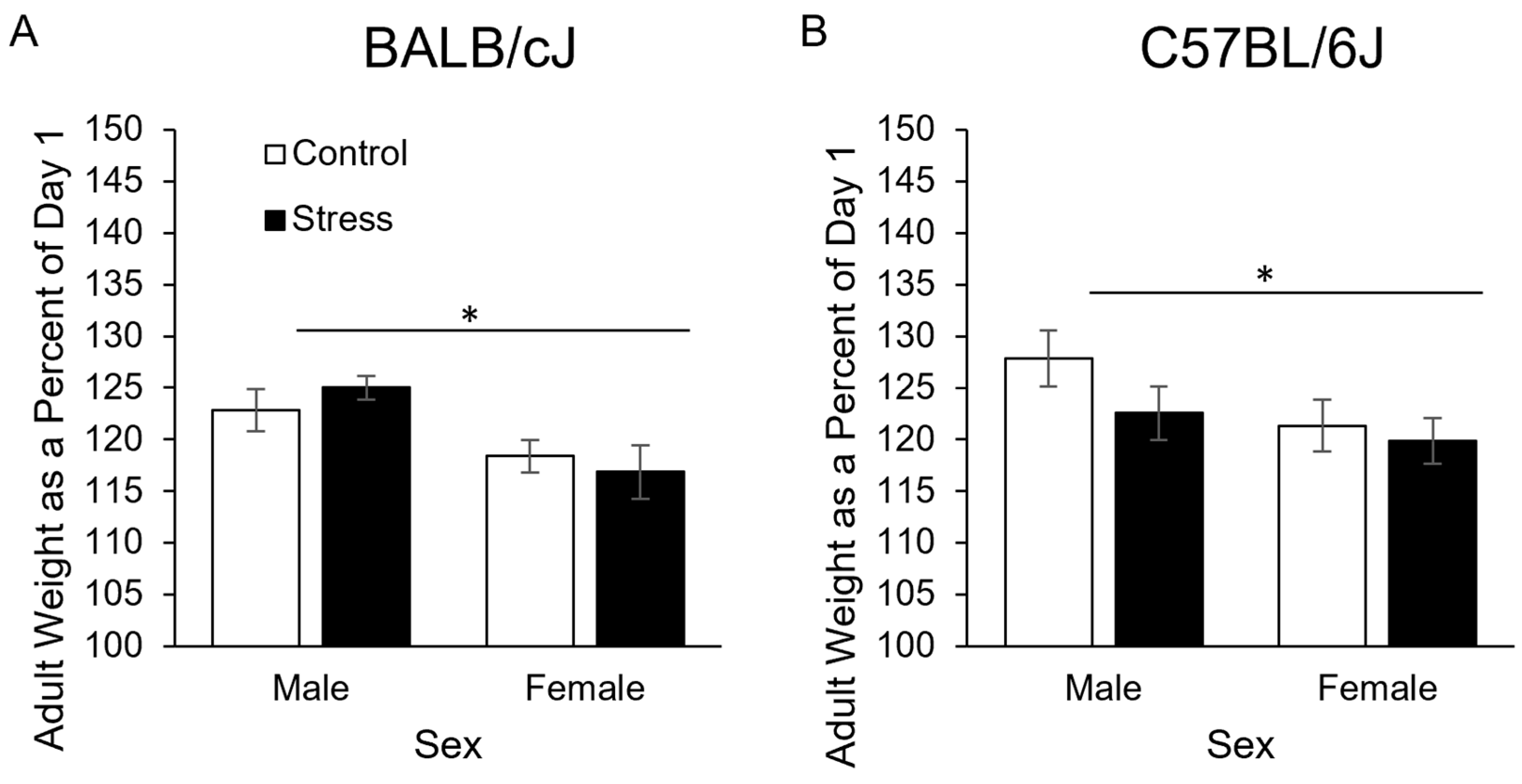
3.1.3. In Adulthood at Sacrifice
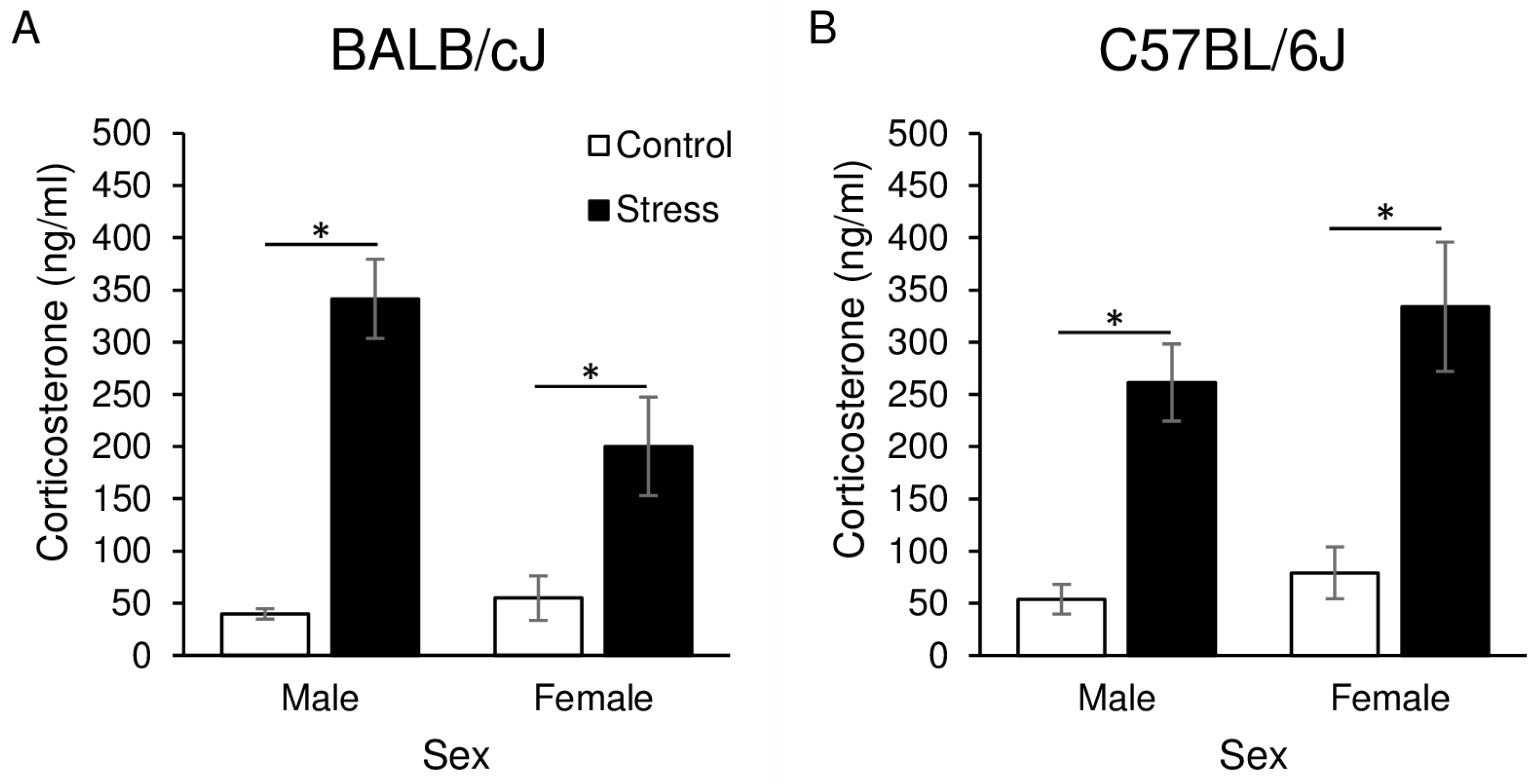
3.2. Corticosterone
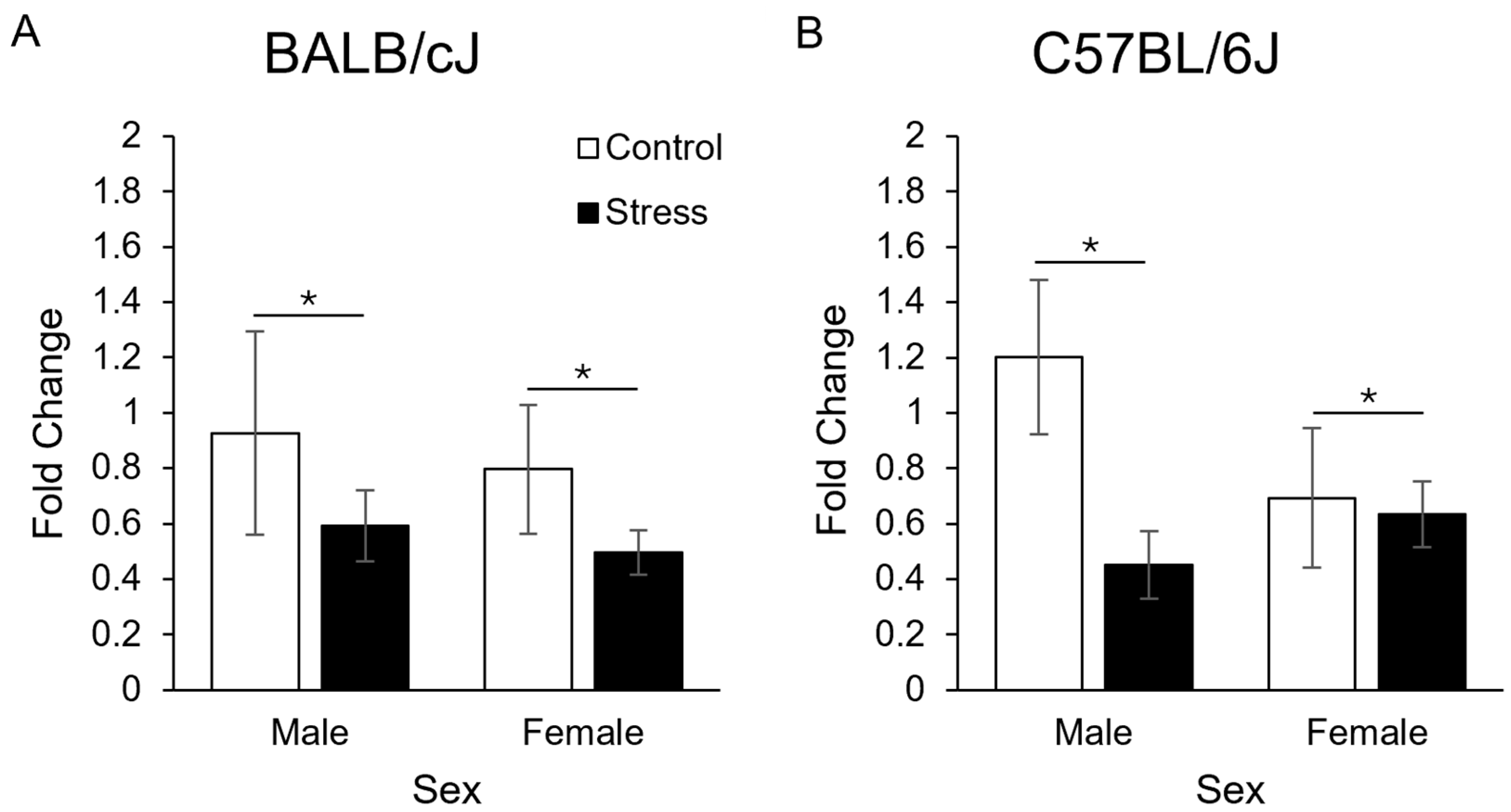
3.3. Gene Expression
3.4. Bioinformatics Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lo Iacono, L.; Carola, V. The impact of adolescent stress experiences on neurobiological development. Semin. Cell Dev. Biol. 2018, 77, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Serafini, G.; Pompili, M.; Innamorati, M.; Giordano, G.; Montebovi, F.; Sher, L.; Dwivedi, Y.; Girardi, P. The role of microRNAs in synaptic plasticity, major affective disorders and suicidal behavior. Neurosci. Res. 2012, 73, 179–190. [Google Scholar] [CrossRef]
- Leung, A.K.L.; Sharp, P.A. MicroRNA functions in stress responses. Mol. Cell. 2010, 40, 205–215. [Google Scholar] [CrossRef]
- Vázquez-Ágredos, A.; Gámiz, F.; Gallo, M. MicroRNA Regulation of the Environmental Impact on Adolescent Neurobehavioral Development: A Systematic Review. Front. Cell. Neurosci. 2022, 16, 956609. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Kamens, H.M.; Miller, C.N.; Caulfield, J.I.; Zeid, D.; Horton, W.J.; Silva, C.P.; Sebastian, A.; Albert, I.; Gould, T.J.; Fishbein, D.; et al. Adolescent Stress Reduces Adult Morphine-Induced Behavioral Sensitization in C57BL/6J Mice. Front. Behav. Neurosci. 2021, 15, 678102. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayanan, S.K.; Shih, Y.-H.; Wen, Y.-R.; Palani, M.; Lin, Y.-W.; Su, H.; Gałecki, P.; Su, K.-P. miR-200a-3p modulates gene expression in comorbid pain and depression: Molecular implication for central sensitization. Brain Behav. Immun. 2019, 82, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-B.; Luo, L.; Liu, X.-L.; Geng, D.; Liu, Q.; Yi, L.-T. 7-Chlorokynurenic acid (7-CTKA) produces rapid antidepressant-like effects: Through regulating hippocampal microRNA expressions involved in TrkB-ERK/Akt signaling pathways in mice exposed to chronic unpredictable mild stress. Psychopharmacology 2015, 232, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Sadler, A.M.; Bailey, S.J. Repeated daily restraint stress induces adaptive behavioural changes in both adult and juvenile mice. Physiol. Behav. 2016, 167, 313–323. [Google Scholar] [CrossRef]
- Romeo, R.D.; Bellani, R.; Karatsoreos, I.N.; Chhua, N.; Vernov, M.; Conrad, C.D.; McEwen, B.S. Stress history and pubertal development interact to shape hypothalamic-pituitary-adrenal axis plasticity. Endocrinology 2006, 147, 1664–1674. [Google Scholar] [CrossRef]
- Duarte, J.O.; Planeta, C.S.; Crestani, C.C. Immediate and long-term effects of psychological stress during adolescence in cardiovascular function: Comparison of homotypic vs. heterotypic stress regimens. Int. J. Dev. Neurosci. 2015, 40, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Gergues, M.M.; Han, K.J.; Choi, H.S.; Brown, B.; Clausing, K.J.; Turner, V.S.; Vainchtein, I.D.; Molofsky, A.V.; Kheirbek, M.A. Circuit and molecular architecture of a ventral hippocampal network. Nat. Neurosci. 2020, 23, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Tsuchimine, S.; Matsuno, H.; O’hashi, K.; Chiba, S.; Yoshimura, A.; Kunugi, H.; Sohya, K. Comparison of physiological and behavioral responses to chronic restraint stress between C57BL/6J and BALB/c mice. Biochem. Biophys. Res. Commun. 2020, 525, 33–38. [Google Scholar] [CrossRef]
- Caruso, M.J.; Kamens, H.M.; Cavigelli, S.A. Exposure to chronic variable social stress during adolescence alters affect-related behaviors and adrenocortical activity in adult male and female inbred mice. Dev. Psychobiol. 2017, 59, 679–687. [Google Scholar] [CrossRef]
- Caruso, M.J.; Crowley, N.A.; Reiss, D.E.; Caulfield, J.I.; Luscher, B.; Cavigelli, S.A.; Kamens, H.M. Adolescent Social Stress Increases Anxiety-like Behavior and Alters Synaptic Transmission, Without Influencing Nicotine Responses, in a Sex-Dependent Manner. Neuroscience 2018, 373, 182–198. [Google Scholar] [CrossRef]
- Caruso, M.J.; Reiss, D.E.; Caulfield, J.I.; Thomas, J.L.; Baker, A.N.; Cavigelli, S.A.; Kamens, H.M. Adolescent chronic variable social stress influences exploratory behavior and nicotine responses in male, but not female, BALB/cJ mice. Brain Res. Bull. 2018, 138, 37–49. [Google Scholar] [CrossRef]
- Tastsoglou, S.; Alexiou, A.; Karagkouni, D.; Skoufos, G.; Zacharopoulou, E.; Hatzigeorgiou, A.G. DIANA-microT 2023: Including predicted targets of virally encoded miRNAs. Nucleic Acids Res. 2023, 51, W148–W153. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- di Salvo, M.L.; Contestabile, R.; Safo, M.K. Vitamin B(6) salvage enzymes: Mechanism, structure and regulation. Biochim. Biophys. Acta 2011, 1814, 1597–1608. [Google Scholar] [CrossRef]
- Al Mughram, M.H.; Ghatge, M.S.; Kellogg, G.E.; Safo, M.K. Elucidating the Interaction between Pyridoxine 5′-Phosphate Oxidase and Dopa Decarboxylase: Activation of B6-Dependent Enzyme. Int. J. Mol. Sci. 2022, 24, 642. [Google Scholar] [CrossRef]
- Schmidt, S.I.; Blaabjerg, M.; Freude, K.; Meyer, M. RhoA Signaling in Neurodegenerative Diseases. Cells 2022, 11, 1520. [Google Scholar] [CrossRef] [PubMed]
- Nowland, M.H.; Hugunin, K.M.S.; Rogers, K.L. Effects of short-term fasting in male Sprague-Dawley rats. Comp. Med. 2011, 61, 138–144. [Google Scholar] [PubMed]
- Bekkevold, C.M.; Robertson, K.L.; Reinhard, M.K.; Battles, A.H.; Rowland, N.E. Dehydration parameters and standards for laboratory mice. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 233–239. [Google Scholar] [PubMed]
- Rinaldi, A.; Vincenti, S.; De Vito, F.; Bozzoni, I.; Oliverio, A.; Presutti, C.; Fragapane, P.; Mele, A. Stress induces region specific alterations in microRNAs expression in mice. Behav. Brain Res. 2010, 208, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-H.; Hong, S.-I.; Ma, S.-X.; Hwang, J.-Y.; Kim, J.-S.; Lee, J.-H.; Seo, J.-Y.; Lee, S.-Y.; Jang, C.-G. Strain differences in the chronic mild stress animal model of depression and anxiety in mice. Biomol. Ther. 2014, 22, 453–459. [Google Scholar] [CrossRef]
- Houdelet, C.; Blondeau-Bidet, E.; Estevez-Villar, M.; Mialhe, X.; Hermet, S.; Ruelle, F.; Dutto, G.; Bajek, A.; Bobe, J.; Geffroy, B. Circulating MicroRNAs Indicative of Sex and Stress in the European Seabass (Dicentrarchus labrax): Toward the Identification of New Biomarkers. Mar. Biotechnol. 2023, 25, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, I.; Ciccarese, F.; Sharova, E.; Urso, L.; Raimondi, V.; Silic-Benussi, M.; D’agostino, D.M.; Ciminale, V. The miR-200 Family of microRNAs: Fine Tuners of Epithelial-Mesenchymal Transition and Circulating Cancer Biomarkers. Cancers 2021, 13, 5874. [Google Scholar] [CrossRef] [PubMed]
- Cattane, N.; Mora, C.; Lopizzo, N.; Borsini, A.; Maj, C.; Pedrini, L.; Rossi, R.; Riva, M.A.; Pariante, C.M.; Cattaneo, A. Identification of a miRNAs signature associated with exposure to stress early in life and enhanced vulnerability for schizophrenia: New insights for the key role of miR-125b-1-3p in neurodevelopmental processes. Schizophr. Res. 2019, 205, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.E.; Chandra, R.; Menken, M.S.; Larkin, E.J.; Nam, H.; Engeln, M.; Francis, T.C.; Lobo, M.K. Dendritic remodeling of D1 neurons by RhoA/Rho-kinase mediates depression-like behavior. Mol. Psychiatry 2020, 25, 1022–1034. [Google Scholar] [CrossRef]
- Zhou, J.; Ma, Y.; Chen, J.; Yao, D.; Feng, C.; Dong, Y.; Ren, Y.; Ma, H.; Wang, Z.; Li, G.; et al. Effects of RhoA on depression-like behavior in prenatally stressed offspring rats. Behav. Brain Res. 2022, 432, 113973. [Google Scholar] [CrossRef]
- Romeo, R.D.; Kaplowitz, E.T.; Ho, A.; Franco, D. The influence of puberty on stress reactivity and forebrain glucocorticoid receptor levels in inbred and outbred strains of male and female mice. Psychoneuroendocrinology 2013, 38, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Chesler, E.J.; Wilson, S.G.; Lariviere, W.R.; Rodriguez-Zas, S.L.; Mogil, J.S. Identification and ranking of genetic and laboratory environment factors influencing a behavioral trait, thermal nociception, via computational analysis of a large data archive. Neurosci. Biobehav. Rev. 2002, 26, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez, S.D.; Riggs, L.M.; Nieto, S.J.; Dayrit, G.; Zamora, N.N.; Shawhan, K.L.; Cruz, B.; Warren, B.L. Social defeat stress induces a depression-like phenotype in adolescent male c57BL/6 mice. Stress 2014, 17, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Pantoja-Urbán, A.H.; Richer, S.; Mittermaier, A.; Giroux, M.; Nouel, D.; Hernandez, G.; Flores, C. Gains and Losses: Resilience to Social Defeat Stress in Adolescent Female Mice. Biol. Psychiatry 2023, 95, 37–47. [Google Scholar] [CrossRef]
- Hyer, M.M.; Shaw, G.A.; Goswamee, P.; Dyer, S.K.; Burns, C.M.; Soriano, E.; Sanchez, C.S.; Rowson, S.A.; McQuiston, A.R.; Neigh, G.N. Chronic adolescent stress causes sustained impairment of cognitive flexibility and hippocampal synaptic strength in female rats. Neurobiol. Stress 2021, 14, 100303. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IPA Top 10 Canonical Pathways | -log(p-Value) |
|---|---|
| Pyridoxal 5′-phosphate Salvage Pathway | 6.86 |
| Epithelial Adherens Junction Signaling | 4.6 |
| Salvage Pathways of Pyrimidine Ribonucleotides | 4.53 |
| DNA Methylation and Transcriptional Repression Signaling | 4.4 |
| Synaptogenesis Signaling Pathway | 4.02 |
| Pulmonary Healing Signaling Pathway | 3.19 |
| Gap Junction Signaling | 3.19 |
| GNRH Signaling | 2.9 |
| RHOA Signaling | 2.78 |
| HOTAIR Regulatory Pathway | 2.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamens, H.M.; Anziano, E.K.; Horton, W.J.; Cavigelli, S.A. Chronic Adolescent Restraint Stress Downregulates miRNA-200a Expression in Male and Female C57BL/6J and BALB/cJ Mice. Genes 2024, 15, 873. https://doi.org/10.3390/genes15070873
Kamens HM, Anziano EK, Horton WJ, Cavigelli SA. Chronic Adolescent Restraint Stress Downregulates miRNA-200a Expression in Male and Female C57BL/6J and BALB/cJ Mice. Genes. 2024; 15(7):873. https://doi.org/10.3390/genes15070873
Chicago/Turabian StyleKamens, Helen M., Emma K. Anziano, William J. Horton, and Sonia A. Cavigelli. 2024. "Chronic Adolescent Restraint Stress Downregulates miRNA-200a Expression in Male and Female C57BL/6J and BALB/cJ Mice" Genes 15, no. 7: 873. https://doi.org/10.3390/genes15070873