

How Important Are Genetic Diversity and Cultivar Uniformity in Wheat? The Case of Gliadins


Abstract
:1. History: Genetic Diversity Needed
2. The Problem
3. Gliadin Polymorphism Provides the Solution
4. More Knowledge about Wheat Polymorphism Worldwide
5. Why Is Wheat Germplasm So Structured?
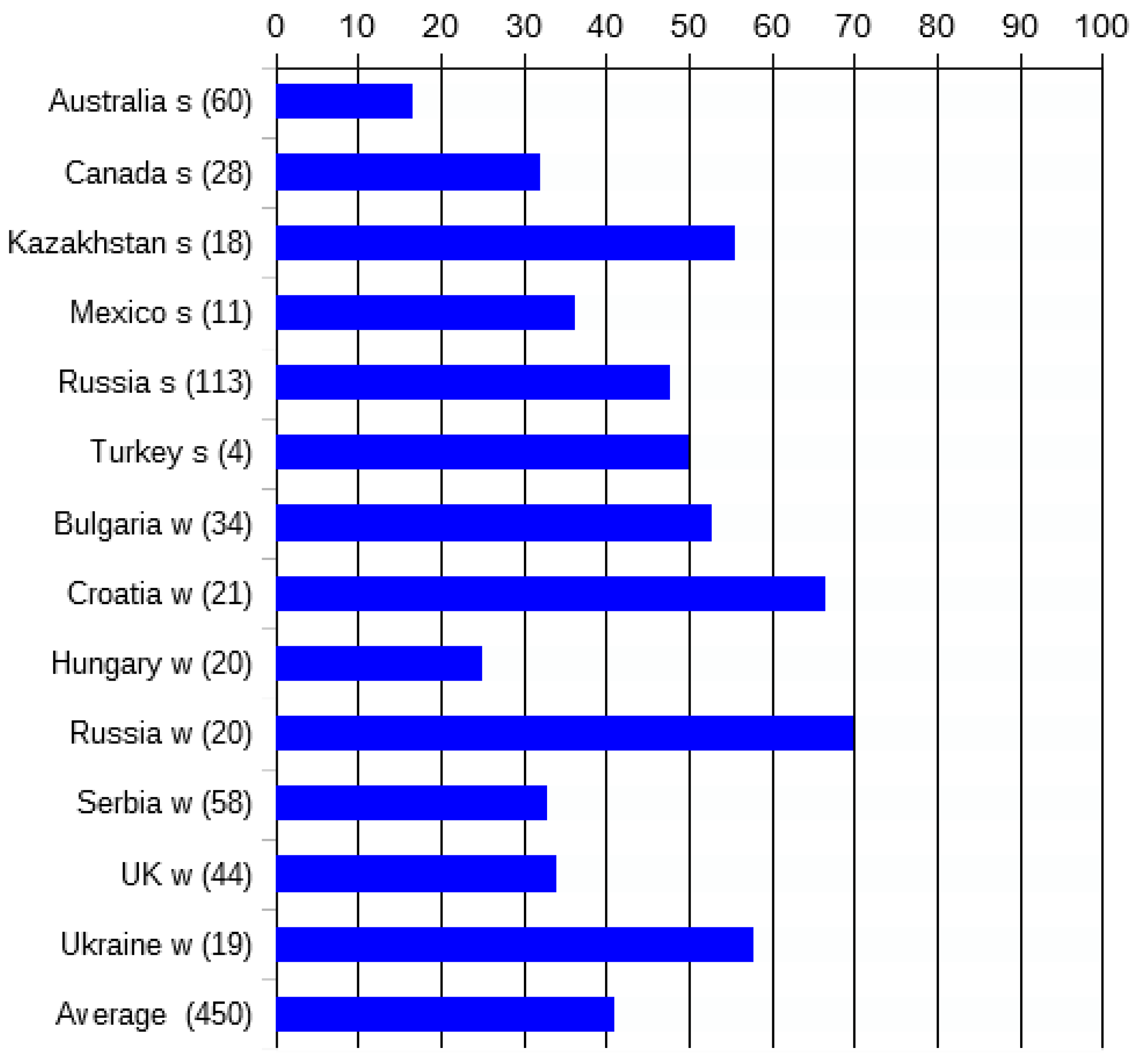
6. Genetic Uniformity Assured? Not for Over 40% of Registrations
7. Is Genetic Uniformity Wholly Desirable?
8. Lessons to Be Learned
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, W.; Scoles, G.; Hucl, P.; Chibbar, R.N. The use of RAPD analysis to classify Triticum accessions. Theor. Appl. Genet. 1999, 98, 602–607. [Google Scholar] [CrossRef]
- Ferns, G.K.; Fitzsimmons, R.W.; Martin, R.H.; Simmonds, D.H.; Wrigley, C.W. Australian Wheat Varieties: Identification according to Growth, Head and Grain Characteristics, 1st ed.; Csiro Publishing: Melbourne, Australia, 1975. [Google Scholar]
- Fitzsimmons, R.W.; Martin, R.H.; Simmonds, D.H.; Wrigley, C.W. Australian Wheat Varieties: Identification according to Growth, Head and Grain Characteristics, 2nd ed.; Csiro Publishing: Melbourne, Australia, 1983. [Google Scholar]
- Ruiz, M.; Metakovsky, E.; Rodriguez-Quijano, M.; Vazquez, J.F.; Carrillo, J.M. Assessment of storage protein variation in relation to some morphological characters in a sample of Spanish landraces of common wheat (Triticum aestivum L. ssp aestivum). Genet. Resour. Crop Evol. 2002, 49, 371–382. [Google Scholar] [CrossRef]
- Juhasz, A.; Larroque, O.R.; Támas, L.; Hsam, S.L.K.; Zeller, F.J.; Békés, F.; Bedö, Z. Bankuti 1201—An old Hungarian wheat variety with special storage protein composition. Theor. Appl. Genet. 2003, 107, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Carvalho, C.; Guedes-Pinto, H.; Igrejas, G.; Stephenson, P.; Schwarzacher, T.; Heslop-Harrison, J.S. High levels of genetic diversity throughout the range of the Portuguese wheat landrace ‘Barbela’. Ann. Bot. 2004, 94, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Caballero, L.; Pena, R.J.; Martin, L.M.; Alvarez, J.B. Characterization of Mexican Creole wheat landraces in relation to morphological characteristics and HMW glutenin subunit composition. Genet. Resour. Crop Evol. 2010, 57, 657–665. [Google Scholar] [CrossRef]
- Wrigley, C.W.; Farrer, W.J.; Guthrie, F.B. The unique breeder-chemist combination that pioneered quality wheats for Australia. Rec. Aust. Acad. Sci. 1979, 4, 7–25. [Google Scholar] [CrossRef]
- Wrigley, C.W.; Rathjen, A. Wheat Breeding in Australia. In Plants and Man in Australia; Carr, D.J., Carr, S.G.M., Eds.; Academic Press: Sydney, Australia, 1981; pp. 96–135. [Google Scholar]
- Macindoe, S.L.; Brown, C.W. Wheat Breeding and Varieties in Australia. In Science Bulletin N76, 3rd ed.; New South Wales Department of Agriculture: Sydney, Australia, 1968. [Google Scholar]
- Cooke, R.J.; Wrigley, C.W. Variety registration and breeders’ rights. In Encyclopedia of Grain Science; Wrigley, C.W., Corke, H., Eds.; Elsevier Ltd.: Oxford, UK, 2004; pp. 313–320. [Google Scholar]
- Harlan, J.R. Genetics of Disaster. J. Environ. Qual. 1972, 1, 212–215. [Google Scholar] [CrossRef]
- Porceddu, E.; Ceoloi, C.; Lafiandra, D.; Tanzarella, O.A.; Munozza, G.T.S. Genetic resources and plant breeding. In Proceedings of the Seventh International Wheat Genetics Symposium, Cambridge, UK, 13–19 July 1988; Problems and Pospects. Miller, T.E., Koebner, R.M.D., Eds.; Institute of Plant Science Research: Cambridge, UK, 1988; pp. 7–22. [Google Scholar]
- Fu, Y.-B.; Somers, D.J. Genome-Wide Reduction of Genetic Diversity in Wheat Breeding. Crop Sci. 2009, 49, 161–168. [Google Scholar] [CrossRef]
- Thormann, I.; Engels, J.M.M. Genetic Diversity and Erosion—A Global Perspective. In Genetic Diversity and Erosion in Plants; Sustainable Development and Biodiversity; Ahuja, M., Jain, S., Eds.; Springer: Cham, Switzerland, 2015; Volume 7, pp. 263–294. [Google Scholar] [CrossRef]
- Barak, S.; Mudgil, D.; Khatkar, B.A. Biochemical and functional properties of wheat gliadins: A review. Critical Reviews in Food. Sci. Nutr. 2015, 55, 357–368. [Google Scholar] [CrossRef]
- Bushuk, W.; Zillman, R.R. Wheat cultivar identification by gliadin electrophoregrams. I. Apparatus, method and nomenclature. Can. J. Plant Sci. 1978, 58, 505–515. [Google Scholar] [CrossRef]
- Autran, J.C.; Bushuk, W.; Wrigley, C.W.; Zillman, R.R. Wheat cultivar identification by gliadin electrophoregrams. IV. Comparison of international methods. Cereal Foods World 1979, 24, 471–475. [Google Scholar]
- Zillman, R.R.; Bushuk, W. Wheat cultivar identification by gliadin electrophoregrams. II. Effects of environmental and experimental factors on the gliadin electrophoregrams. Can. J. Plant Sci. 1979, 59, 281–286. [Google Scholar] [CrossRef]
- Cros, D.L.D.; Wrigley, C.W. Improved electrophoretic methods for identifying cereal varieties. J. Sci. Food Agric. 1979, 30, 785–794. [Google Scholar] [CrossRef]
- Lookhart, G.L.; Finney, K.F. Polyacrylamide gel electrophoresis of wheat gliadins: The effect of environment and germination. Cereal Chem. 1984, 61, 496–499. Available online: https://www.cerealsgrains.org/publications/cc/backissues/1984/Documents/CC1984a138.html (accessed on 14 July 2024).
- ICC Standard №143; Wheat—Identification of Varieties by Electrophoresis. International Association for Cereal Science and Technology: Vienna, Austria, 1995; pp. 1–13.
- Metakovsky, E.; Pascual, L.; Vaccino, P.; Melnik, V.A.; Rodriguez-Quijano, M.; Popovych, Y.; Chebotar, S.; Rogers, W.J. Heteroalleles in Common Wheat: Multiple Differences between Allelic Variants of the Gli-B1 Locus. Int. J. Mol. Sci. 2021, 22, 1832. [Google Scholar] [CrossRef] [PubMed]
- Sozinov, A.A.; Poperelya, F.A. Genetic classification of prolamins and its use for plant breeding. Ann. Technol. Agric. 1980, 29, 229–245. [Google Scholar]
- Metakousky, E.; Melnik, V.A.; Rodriguez-Quijano, M.; Upelniek, V.; Carrillo, J.M. A catalog of gliadin alleles: Polymorphism of 20th-century common wheat germplasm. Crop J. 2018, 6, 628–641. [Google Scholar] [CrossRef]
- Zhang, W.; Gianibelli, M.C.; Ma, W.; Rampling, L.R.; Gale, K.R. Identification of SNPs and development of allele-specific PCR markers for γ-gliadin alleles in Triticum aestivum. Theor. Appl. Genet. 2003, 107, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Popovych, Y.; Chebotar, S.; Melnik, V.A.; Rodriguez-Quijano, M.; Pascual, L.; Rogers, W.J.; Metakovsky, E. Congruity of the polymorphisms in the expressed and noncoding parts of the Gli-B1 locus in common wheat. Agronomy 2020, 10, 1510. [Google Scholar] [CrossRef]
- van de Wouw, M.; van Hintum, T.; Kik, C.; van Treuren, R.; Visser, B. Genetic diversity trends in twentieth century crop cultivars: A meta analysis. Theor. Appl. Genet. 2010, 120, 1241–1252. [Google Scholar] [CrossRef]
- Metakousky, E.; Melnik, V.A.; Pascual, L.; Wrigley, C.W. Gliadin genotypes worldwide for spring wheats (Triticum aestivum L). 1. Genetic diversity and grain-quality gliadin alleles during the 20th century. J. Cereal Sci. 2019, 87, 172–177. [Google Scholar] [CrossRef]
- Sansaloni, C.; Franco, J.; Santos, B.; Percival-Alwyn, L.; Singh, S.; Petroli, C.; Campos, J.; Dreher, K.; Payne, T.; Marshall, D.; et al. Diversity analysis of 80,000 wheat accessions reveals consequences and opportunities of selection footprints. Nat. Commun. 2020, 11, 4572. [Google Scholar] [CrossRef] [PubMed]
- Metakovsky, E.; Melnik, V.A.; Pascual, L.; Wrigley, C.W. Gliadin genotypes worldwide for spring wheats (Triticum aestivum L.) 2. Strong differentiation of polymorphism between countries and regions of origin. J. Cereal Sci. 2019, 87, 311–317. [Google Scholar] [CrossRef]
- Nevo, E. Evolutionary significance of genetic diversity in nature: Environmental stress, pattern and theory. In Isozymes Organization and Roles in Evolution, Genetics and Physiology; Proc. 7th Intern. Congr. Isozymes (Novosibirsk, Rusia 6-13.09.1992); Markert, C.L., Scandalios, J.G., Lim, H.A., Serov, O.L., Eds.; World Scientific: Hackensack, NJ, USA, 1994; pp. 267–296. [Google Scholar]
- Pagnotta, M.A.; Nevo, E.; Beiles, A.; Porceddu, E. Wheat storage proteins—glutenin diversity in wild emmer, Triticum-dicoccoides, in Israel and Turkey. 2. DNA diversity detected by PCR. Theor. Appl. Genet. 1995, 91, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Nevo, E.; Pagnotta, M.A.; Beiles, A.; Porceddu, E. Wheat storage proteins—glutenin DNA diversity in wild emmer wheat, Triticum-dicoccoides, in Israel and Turkey. 3. Environmental correlates and allozymic associations. Theor. Appl. Genet. 1995, 91, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, C.R.; Chao, C.R.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [PubMed]
- Koliuchiĭ, V.T.; Sozinov, A.A. The effect of natural selection on the allelic frequency of gliadin-coding loci and their associations in an artificially created hybrid population of winter wheat. Tsitology Genet. 2000, 34, 32–38. [Google Scholar]
- Raquin, A.L.; Depaulis, F.; Lambert, A.; Galic, H.; Brabant, P.; Goldringer, I. Experimental estimation of mutation rates in a wheat population with a gene genealogy approach. Genetics 2008, 179, 2195–2211. [Google Scholar] [CrossRef]
- Stachel, M.; Lelley, T.; Grausgruber, H.; Vollmann, J. Application of microsatellites in wheat (Triticum aestivum L.) for stadying genetic differentiation caused by selection for adaptation and use. Theor. Appl. Genet. 2000, 100, 242–248. [Google Scholar] [CrossRef]
- Prasad, M.; Varshney, R.K.; Roy, J.K.; Balyan, H.S.; Gupta, P.K. The use of microsatellites for detecting DNA polymorphism, genotype identification and genetic diversity in wheat. Theor. Appl. Genet. 2000, 100, 584–592. [Google Scholar] [CrossRef]
- Ehdaie, B.; Waines, J.G. Adaptation of landrace and improved spring wheat genotypes to stress environments. J. Genet. Breed. 1989, 43, 151–156. [Google Scholar]
- Allard, R.W. Genetic basis of the evolution of adaptedness in plants. Euphytica 1996, 92, 1–11. [Google Scholar] [CrossRef]
- Reif, J.C.; Zhang, P.; Dreisigacker, S.; Warburton, M.L.; van Ginkel, M.; Hoisington, D.; Bohn, M.; Melchinger, A.E. Wheat genetic diversity trends during domestication and breeding. Theor. Appl. Genet. 2005, 110, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Perronne, R.; Makowski, D.; Goffaux, R.; Montalent, P.; Goldringer, I. Temporal evolution of varietal, spatial and genetic diversity of bread wheat between 1980 and 2006 strongly depends upon agricultural regions in France. Agric. Ecosyst. Environ. 2017, 236, 12–20. [Google Scholar] [CrossRef]
- Walkowiak, S.; Gao, L.; Monat, C.; Haberer, G.; Kassa, M.T.; Brinton, J.; Ramirez-Gonzalez, R.H.; Kolodziej, M.C.; Delorean, E.; Thambugala, D.; et al. Multiple wheat genomes reveal global variation in modern breeding. Nature 2020, 588, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, C.W.; Shepherd, K.W. Identification of Australian wheat cultivars by laboratory procedures: Examination of pure samples of grain. Aust. J. Exp. Agric. 1974, 14, 796–804. [Google Scholar] [CrossRef]
- Appleyard, D.B.; McCausland, J.; Wrigley, C.W. Checking the identity and origin of off-types in the propagation of pedigreed wheat seed. Seed Sci. Technol. 1979, 7, 459–466. [Google Scholar]
- Pogna, N.E.; Peruffo, A.D.B.; Boggini, G.; Corbellini, M. Analysis of wheat varieties y gliadin electrophoregrams. II. Nature, origin and quality of biotypes present in six Italian common wheat varieties. Genet. Agrar. 1982, 6, 143–154. [Google Scholar]
- Metakovsky, E.; Melnik, V.; Pascual, L.; Romanov, G.A.; Wrigley, C.W. Types, frequencies and value of intra-varietal genotypic non-uniformity in common wheat cultivars: Authentic biotypes and foreign seeds. J. Cereal Sci. 2019, 89, 102813. [Google Scholar] [CrossRef]
- Landjeva, S.; Korzun, V.; Ganeva, G. Evaluation of genetic diversity among Bulgarian winter wheat (Triticum aestivum L.) varieties during the period 1925–2003 using microsatellites. Genet. Resour. Crop Evol. 2006, 53, 1605–1614. [Google Scholar] [CrossRef]
- Lawrence, G.J.; Moss, H.J.; Shepherd, K.W.; Wrigley, C.W. Dough quality of biotypes of eleven Australian wheat cultivars that differ in high-molecular-weight glutenin subunit composition. J. Cereal Sci. 1987, 6, 99–101. [Google Scholar] [CrossRef]
- Gupta, R.B.; Shepherd, K.W. Low-molecular-weight glutenin subunits in wheat: Their variation, inheritance and association with bread-making quality. In Proceedings of the 7th International Wheat Genetics Symp, IPSR, Cambridge, UK, 13–19 July 1988; Miller, T.E., Koebner, R.M.D., Eds.; Orbach Science Library: Riverside, CA, USA, 1988; pp. 943–949. [Google Scholar]
- Wrigley, C.W.; Autran, J.C.; Bushuk, W. Identification of cereal varieties by gel electrophoresis of the grain proteins. Adv. Cereal Sci. Technol. 1982, 5, 211–259. [Google Scholar]
- Kozub, N.; Pascual, L.; Sobko, T.; Chebotar, S.; Metakovsky, E. Genotypes of common wheat cultivars Bred in south Ukraine can be easily distinguished from any other wheat genotypes. J. Cereal Sci. 2024, 117, 103916. [Google Scholar] [CrossRef]
- Metakovsky, E.; Sozinov, A.A. Genetics of gliadin proteins and the problems of interpreting results obtained with somaclonal variation. In Wheat Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 13, pp. 526–537. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | N1 | Gli-A1 | Gli-B1 | Gli-D1 | Gli-A2 | Gli-B2 | Gli-D2 | Average |
|---|---|---|---|---|---|---|---|---|
| 1901–1933 | 46 | 858 | 888 | 764 | 924 | 870 | 867 | 862 ± 43 |
| 1934–1950 | 60 | 857 | 874 | 695 | 925 | 915 | 881 | 858 ± 67 |
| 1951–1966 | 102 | 889 | 862 | 818 | 917 | 907 | 911 | 884 ± 30 |
| 1967–1976 | 183 | 870 | 824 | 787 | 895 | 887 | 895 | 860 ± 35 |
| 1977–1982 | 191 | 878 | 823 | 694 | 904 | 892 | 877 | 845 ± 63 |
| 1983–1986 | 143 | 867 | 826 | 662 | 888 | 906 | 873 | 837 ± 72 |
| 1987–1998 | 143 | 858 | 773 | 695 | 897 | 886 | 842 | 825 ± 62 |
| Cultivar | Gli-A1 | Gli-B1 | Gli-D1 | Gli-A2 | Gli-B2 | Gli-D2 |
|---|---|---|---|---|---|---|
| Suneca 1 | o | d | f | m | c | m + j 2 |
| Ciano-67 | o | d | f | f | c | j |
| Spica | o | b | a | m | c + o | m |
| Brasilia | a | e + g | a | g + e | v + j | r + a |
| Osječka-20 | a | g | b | g | j | r |
| Zlatna-Dolina | b | e | a | e | e | m |
| Libellula | a | k | b | g | v | a |
| Bezostaya-1 | b | b | b | b | b | b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metakovsky, E.; Melnik, V.A.; Pascual, L.; Wrigley, C.W. How Important Are Genetic Diversity and Cultivar Uniformity in Wheat? The Case of Gliadins. Genes 2024, 15, 927. https://doi.org/10.3390/genes15070927
Metakovsky E, Melnik VA, Pascual L, Wrigley CW. How Important Are Genetic Diversity and Cultivar Uniformity in Wheat? The Case of Gliadins. Genes. 2024; 15(7):927. https://doi.org/10.3390/genes15070927
Chicago/Turabian StyleMetakovsky, Eugene, Viktor A. Melnik, Laura Pascual, and Colin W. Wrigley. 2024. "How Important Are Genetic Diversity and Cultivar Uniformity in Wheat? The Case of Gliadins" Genes 15, no. 7: 927. https://doi.org/10.3390/genes15070927