Abstract
Inosine is a nucleotide resulting from the deamination of adenosine in RNA. This chemical modification process, known as RNA editing, is typically mediated by a family of double-stranded RNA binding proteins named Adenosine Deaminase Acting on dsRNA (ADAR). While the presence of ADAR orthologs has been traced throughout the evolution of metazoans, the existence and extension of RNA editing have been characterized in a more limited number of animals so far. Undoubtedly, ADAR-mediated RNA editing plays a vital role in physiology, organismal development and disease, making the understanding of the evolutionary conservation of this phenomenon pivotal to a deep characterization of relevant biological processes. However, the lack of direct high-throughput methods to reveal RNA modifications at single nucleotide resolution limited an extended investigation of RNA editing. Nowadays, these methods have been developed, and appropriate bioinformatic pipelines are required to fully exploit this data, which can complement existing approaches to detect ADAR editing. Here, we review the current literature on the “bioinformatics for inosine” subject and we discuss future research avenues in the field.
1. Inosine and ADAR Protein Discovery
The presence of naturally occurring RNA modifications was first reported in 1960 [1,2,3], with over 170 distinct RNA modifications identified so far [4]. These modifications can be present in different types of RNA and can influence RNA stability and structure, and the interactions between RNA and cellular or viral proteins [5,6]. Indeed, RNA modifications play crucial roles in regulating RNA metabolism by affecting RNA processing, localization, and translation into proteins, particularly when messenger RNAs (mRNAs), transfer RNAs (tRNAs), and ribosomal RNAs (rRNAs) are impacted [7]. Accordingly, RNA modifications can be seen as a bridge linking gene transcription to protein synthesis, thereby influencing cell functioning. Several tools and methods have been developed to trace the different modifications occurring in RNA molecules. Most of the efforts were in the study of N1-Methyladenosine (m1A), 5-Methylcytosine (m5C), 5-Methyluridine (m5U), N6-Methyladenosine (m6A), N6,2′-O-dimethyladenosine (m6Am), 7-methylguanosine (m7G), pseudouridine (Ψ), and 2′-O-Methylation (Nm) (Table 1, with more details in Supplementary Table S1). RNA editing is a post-transcriptional activity, defined as the insertion, deletion or chemical modification of ribonucleotides in target RNA molecules [8,9,10]. Compared to the eukaryotic RNA splicing and polyadenylation processes, RNA editing is less frequent but still pivotal in modifying the function and stability of RNAs, and its involvement in health and disease is documented by several lines of evidence in humans [11,12,13]. The Adenosine (A) to Inosine (I) deamination (known as A-to-I editing) represents one of the most frequent RNA modifications in metazoans [10]. Inosine was first discovered in tRNAs in 1965 [14] (Figure 1), and later, in 1966, it was shown that inosine could pair with multiple nucleotides, de facto expanding the flexibility in the genetic code during protein synthesis [10] and therefore opening the avenue for the study of RNA editing [15] and modification, a field dubbed epitranscriptomics [16].

Table 1.
List of the most studied RNA modifications with the tools developed for their computational detection. For each tool, the detected modification, the methodology used, the extended reference and the reference number are reported in Supplementary Table S1.
The protein responsible for the enzymatic deamination of adenosine into inosine was initially described in 1987 as a protein capable of unwinding the RNA duplexes in Xenopus laevis embryo, thus impairing the inhibition of mRNA translation by the injection of a complementary antisense RNA [17,18]. One year later, further characterization of this protein demonstrated its capability to modify the RNA enzymatically (Figure 1) [19]. The protein, called Double-stranded RNA-specific adenosine deaminase (abbreviated into DRADA, ADAR or dsRAD), was then deeply studied, revealing a pivotal role in maintaining the organism homeostasis [10,20,21,22].
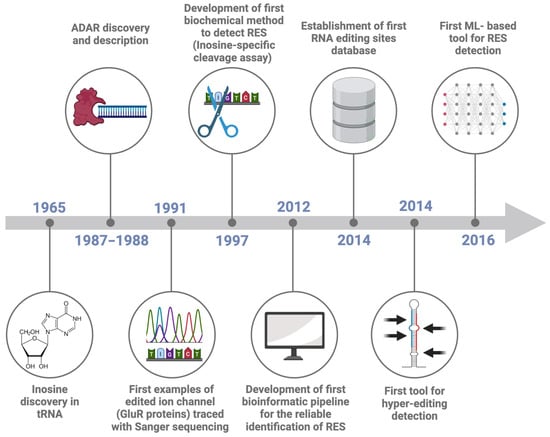
Figure 1.
Timeline of RNA Editing Sites (RES) detection. The Figure reports the most important events in the history of advancing in RNA editing site detection, namely: inosine discovery in tRNA (1965, [14]), the discovery of the protein responsible for RNA editing (1987–1988, [17,18,19]), the first example of edited ion channel (1991, [21]), the development of a first biochemical method to detect inosine (1997, [23]), the development of a first bioinformatic pipeline for the reliable identification of RES (2012, [24]), the creation of the first RES database (2014, [25]), a first tool for hyper-editing detection (2014, [26]) and a first machine learning based tool for RES detection (2016, [27]).
Figure 1.
Timeline of RNA Editing Sites (RES) detection. The Figure reports the most important events in the history of advancing in RNA editing site detection, namely: inosine discovery in tRNA (1965, [14]), the discovery of the protein responsible for RNA editing (1987–1988, [17,18,19]), the first example of edited ion channel (1991, [21]), the development of a first biochemical method to detect inosine (1997, [23]), the development of a first bioinformatic pipeline for the reliable identification of RES (2012, [24]), the creation of the first RES database (2014, [25]), a first tool for hyper-editing detection (2014, [26]) and a first machine learning based tool for RES detection (2016, [27]).
ADAR proteins are characterized by a variable number of dsRNA binding domains and by a deaminase domain. The dsRNA binding domain allows the binding of ADAR to dsRNA structures, whereas the deaminase domain performs the deamination of the target adenosine [28]. In humans, three ADAR paralogs are present. Consistently, in the evolution of animals, ADAR1 generally possesses one or two extra domains, enabling binding to Z-DNA/Z-RNA structures [28]. As for the three mammalian ADAR genes (ADAR1-3), ADAR1 and ADAR2 are expressed throughout the body, but ADAR1 is generally more expressed than ADAR2 and accounts for the majority of the editing activity. Conversely, ADAR3, which is catalytically inactive, is exclusively expressed in the brain [29,30].
At this point, it becomes essential to distinguish between two different types of ADAR-mediated RNA modifications. The first, called single-base editing, results in one or a few edited adenosines per RNA molecule. In contrast, the second type of modification, called hyper-editing, involves multiple A-to-I modifications on the same molecule [26,31,32]. Since the inosine pattern of hydrogen bonds is like that of guanosine, inosine is read as guanosine during protein translation [33,34]. Therefore, A-to-I editing has the potential to alter the coding capacity of mRNAs, in some cases with critical biological consequences [35]. Actually, A-to-I editing of GluR2 transcripts is needed for the normal development of the nervous system in mice and zebrafish [35,36], and perturbed A-to-I editing has been associated with behavioral changes in the fruit fly [37]. In the squid nervous system, extensive A-to-I editing mainly occurs in the giant axons rather than in the cell body—indicating tissue-specific RNA editing—where it can modulate the microtubule motor protein function in response to environmental changes [38,39]. Moreover, A-to-I editing contributes to germline cell integrity by preventing the spread of repetitive transposable elements, such as Alu and SINE in humans and mice, respectively [22,40,41]. Overall, these studies have revealed the importance of ADAR editing in physiology, development and disease, making a better understanding of the occurrence and extension of this phenomenon across the evolution of metazoans necessary.
2. The Development of Inosine Detection Methods
Since 1997, several methods have been developed to trace the enzymatic activity of ADAR. For the sake of clarity, we divided all these methods into direct or indirect ones, according to whether they directly detected inosine or whether the inosine detection was performed indirectly (Table 2).
Table 2.
Description of the direct and indirect methods to detect inosine in biological samples. The name, type, a brief description, and known limitations are reported for each method.
Direct inosine detection methods include low- to medium-throughput approaches, such as two-dimensional, thin-layer chromatography (2D-TLC) [42], inosine-specific cleavage assays [23] and mass spectrometry [43]. In addition, some specific protocols were developed to retrieve the A-to-I modifications in RNA, such as DARTS [44], ICE-seq [45], ALES [46] and hEndoV-seq [47]. Regarding the indirect methods, the most used consists of tracing the inosine footprint using Sanger or high-throughput sequencers. The classical Sanger sequencing method can be adapted for inosine detection by reverse transcribing RNA into cDNA [48]. During reverse transcription, inosine pairs with cytidine and is read as guanosine, resulting in A-to-G mismatches in the sequencing data [21]. Next-Generation Sequencing (NGS) offers both the possibility to generate high-throughput data and the single-nucleotide resolution necessary to detect RNA modifications, following the same principle applied to Sanger sequencing. Although NGS approaches can provide comprehensive and detailed insights about RNA editing, they require extensive data analysis efforts [24].
3. Strategies for Genome-Wide Identification of ADAR-Mediated RNA Editing Sites
Inosine detection methods, particularly indirect ones, present several challenges in the computational detection of genuine RNA Editing Sites (RES) through transcriptional noise. This is the case of massive RNA sequencing data, where the high-throughput output of modern sequencers coupled with an imperfect detection of nucleotides easily introduces confounding factors (i.e., noise), hampering a straightforward detection of low-frequency modifications. False positive RES detection can lead researchers to incorrect conclusions about editing frequencies and their biological implications. Both improving sequencing accuracy and depth as well as validating potential editing sites with independent methods can reduce the impact of false positives. In addition, RNA sequences, especially mRNAs, can contain repetitive elements that complicate the alignment of sequencing reads to reference genomes. Misalignment can lead to incorrect identification of editing sites, leading to both false positives and false negatives. To overcome such a limitation, the use of alignment software optimized for RNA-seq data and the adjustment of alignment parameters to be more stringent with mismatches and gaps can be adopted. Finally, a high-quality, well-annotated reference genome is crucial for accurate mapping of sequencing reads and the subsequent identification of RES. Incomplete or incorrectly-assembled genomes can lead to erroneous conclusions about the presence and extent of RNA editing.
To perform a correct RNA editing analysis, all analytical steps should be planned appropriately. The main steps are file preprocessing, RES detection and RES filtering.
In this review, we discussed the published resources that can facilitate setting up the best possible pipeline for different objectives and organisms, including non-model species lacking a reference genome.
3.1. File Preprocessing
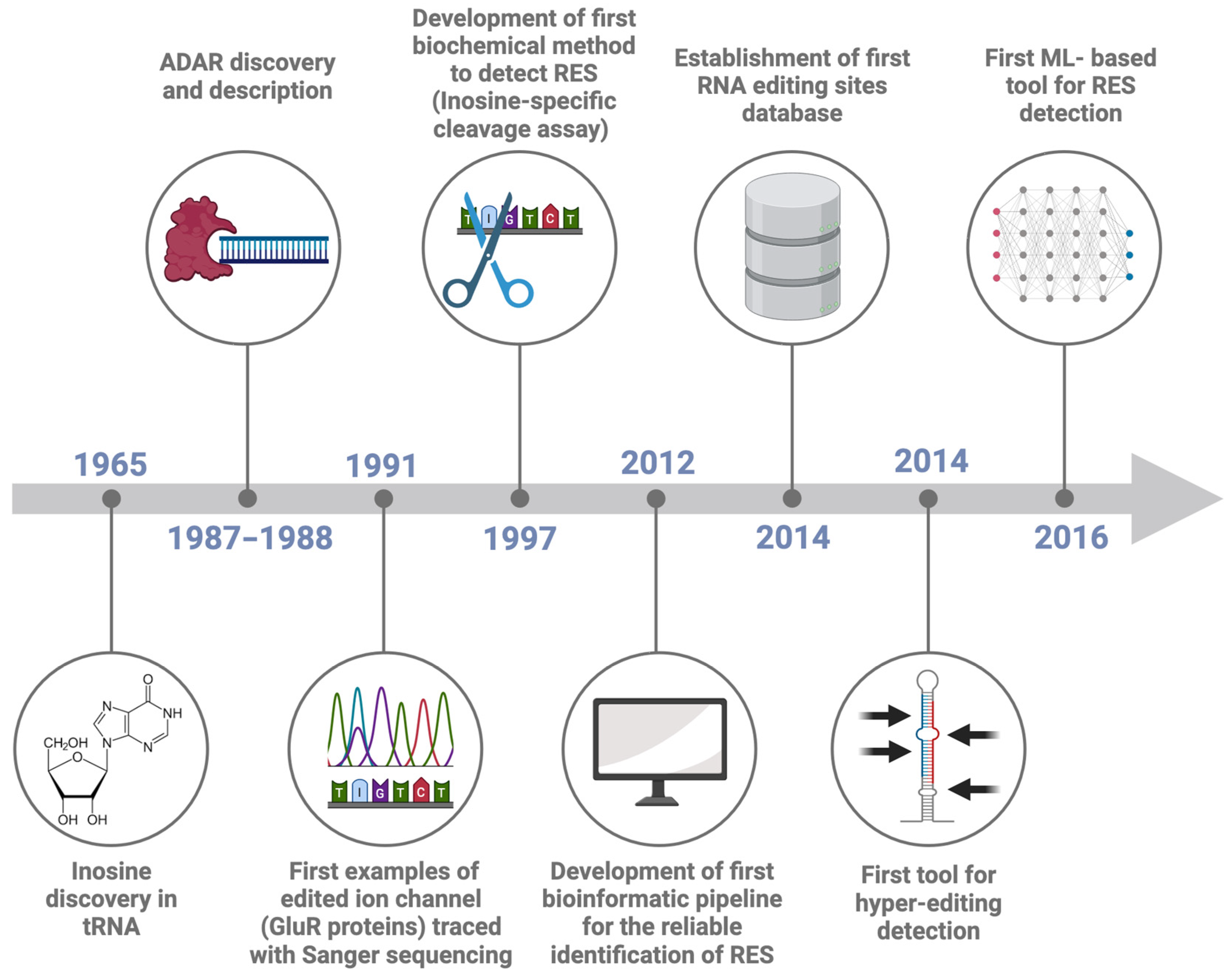
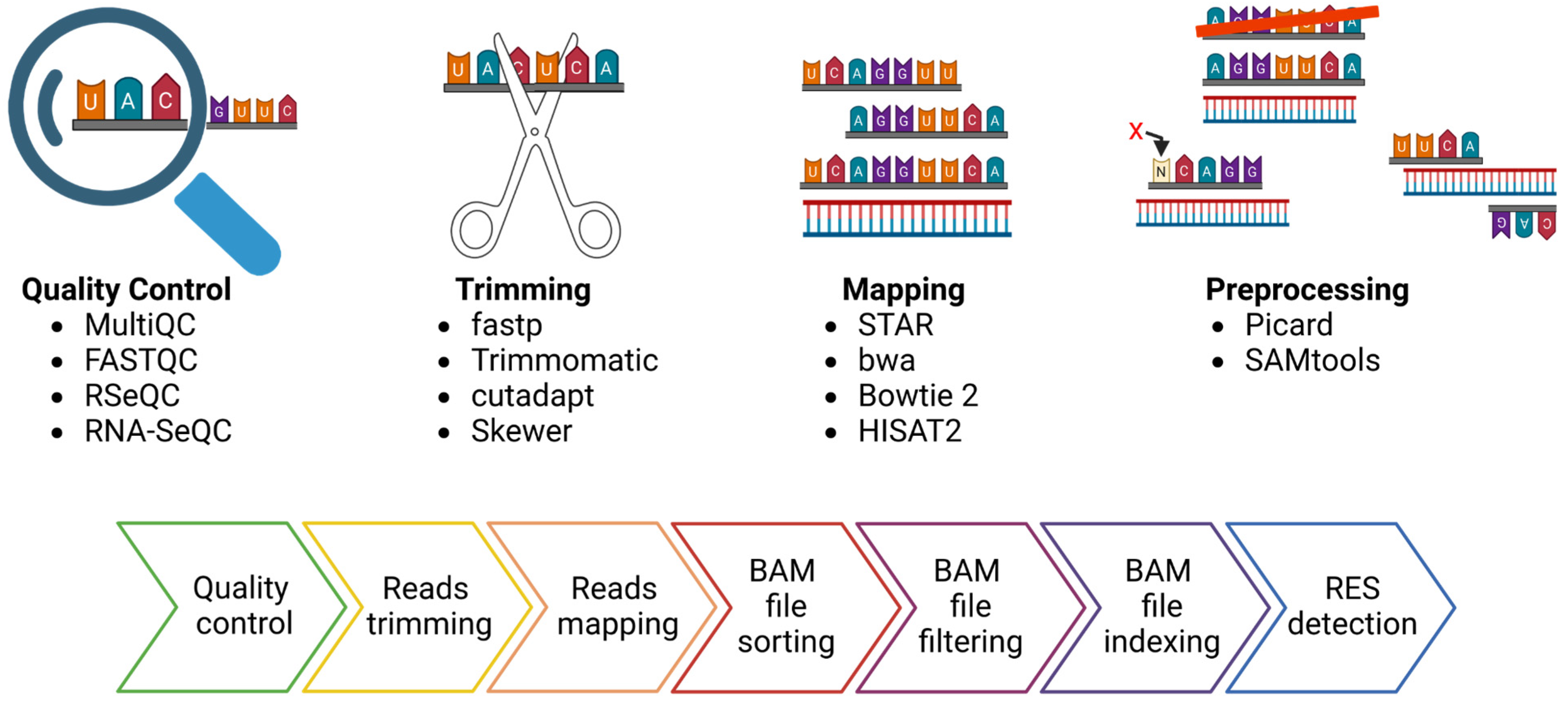
Starting from the sequencer outputs, usually consisting of files in FASTQ format, the RES are traced by performing the following steps: reads quality control and trimming, alignment of the reads to the reference genome and preprocessing of the alignment file (Figure 2). The read quality check and trimming steps are not mandatory, but their application will improve the mapping rate and the alignment quality, two elements that can affect the detection of RES. For these two steps, several tools can be used, such as FASTQC (https://github.com/s-andrews/FastQC (accessed on 1 June 2024)), RSeQC [49] or RNA-SeQC [50], as well as MultiQC [51], a versatile tool designed to create a single comprehensive report by aggregating output from multiple tools applied on several samples. The manual inspection of the outputs of these tools facilitates the identification of global trends and biases in the raw and/or trimmed sequencing data. Similarly, there are multiple tools for data trimming, including fastp [52], Trimmomatic [53], cutadapt [54] and Skewer [55]. The main goal of data trimming is to remove all the remaining sequencing adapters and low-quality bases, which can impair the detection of RES.
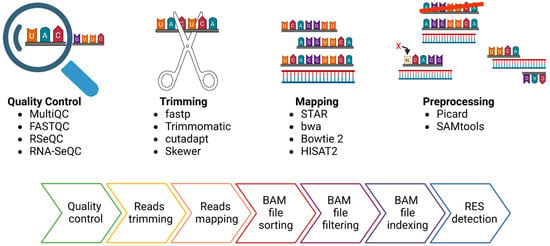
Figure 2.
FASTQ files preprocessing for the detection of RNA Editing Sites (RES). In the upper part, the picture depicts the four most essential steps in preprocessing FASTQ files for RES detection and some available tools for each step. In the lower part of the figure, a more detailed description of the bioinformatic pipeline steps is presented.
As regards the mapping step, there are a variety of mapping tools and algorithms [56]. Several publications have evaluated the performance of the most commonly used mapping tools in terms of speed, accuracy, and usage of computation resources [57,58,59,60]. A recent publication confirmed that using different tools can be critical for the accuracy and resolution of the analysis [57]. Indeed, they analyzed a Genotype-Tissue Expression RNA-seq experiment from the human cerebellum (run accession ID SRR607967), showing that the number of detected sites varied depending on the aligner used. For example, the sequence alignment carried out with STAR resulted in the lowest false positive rate, suggesting its higher performance. An essential parameter regarding the mapping step is to keep only the uniquely mapping reads, avoiding the reads that map in more than one genomic location [57,58]. After the alignment, the resulting SAM file needs to be converted into BAM format, and before using the tools dedicated to RES detection, following some steps are essential to obtain reliable results. The GATK best practices provide a complete pipeline for the BAM preprocessing [61,62]. In this case, the most important steps include the BAM file sorting, the removal of duplicates and the reads filtering, keeping the mapped and properly paired reads with a minimum alignment quality decided by the user (for example, 20). Picard (https://github.com/broadinstitute/picard (accessed on 1 June 2024)) and SAMtools [63] are necessary for these steps (Figure 2).
3.2. Detection of RNA Editing Sites
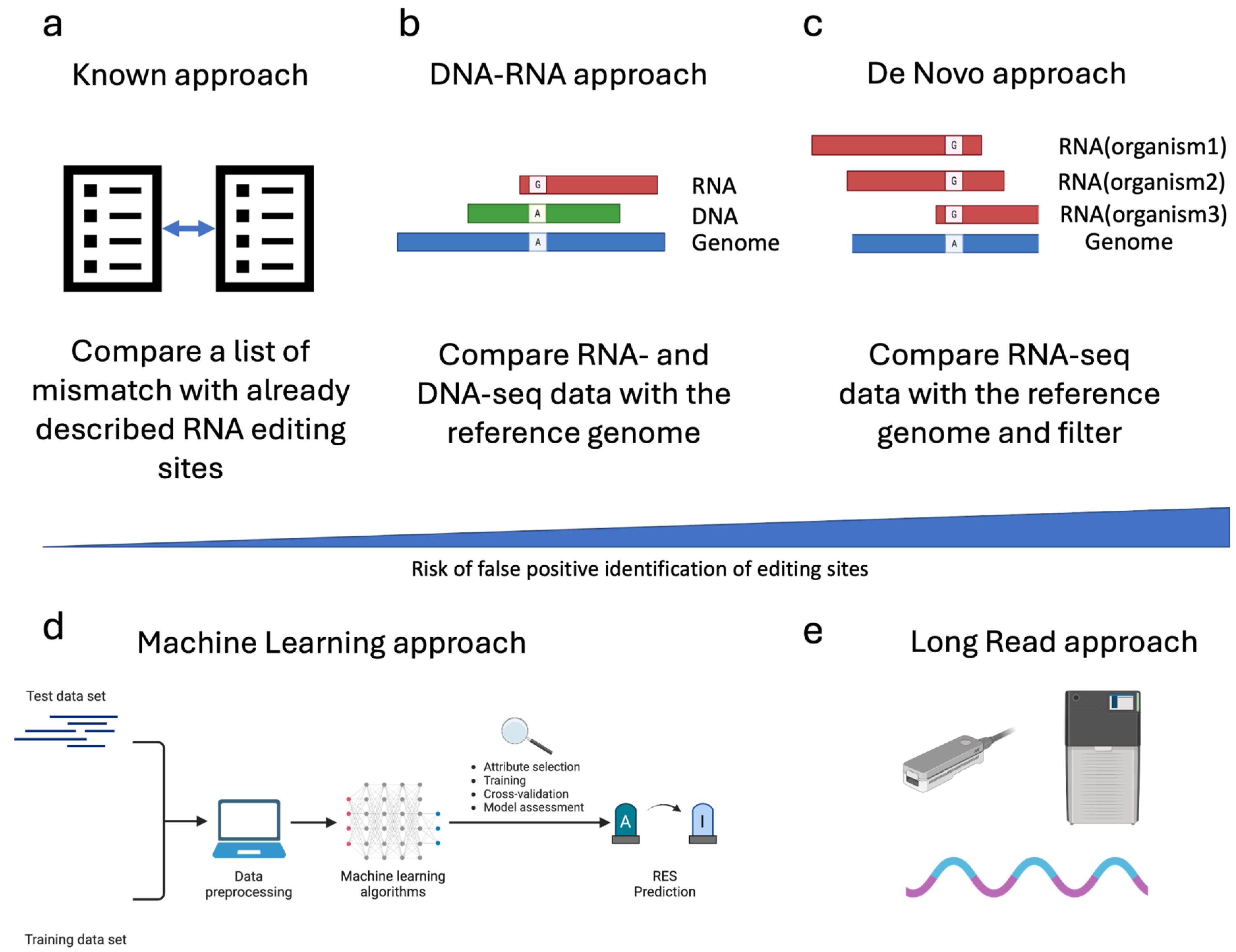
Several strategies can be applied to detect RES. One of the most straightforward approaches is comparing a list of putative editing sites with the already described ones. This approach (named “Known”, Figure 3a) requires a list of validated editing sites, which, so far, are available only for a few species and can be found in the REDIportal database [64]. The most used tool for detecting “Known” sites is REDItools, which has one dedicated module [31].
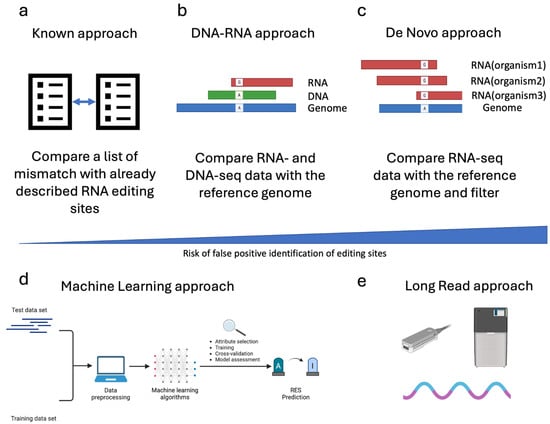
Figure 3.
Strategies used to trace RNA Editing Sites (RES). This figure reports the most common strategies employed to trace RNA editing sites. In the case of Illumina sequencing data, in addition to the approach described based on REDItools nomenclature (Known panel (a), DNA-RNA panel (b) and De Novo panel (c)), an indicative bar showing the risk of false positive rate associated with the different approaches is depicted. Moreover, the most innovative methods still under development for the trace of RES are shown (the Machine learning and long-read approaches, panel (d) and panel (e), respectively).
A second approach, called “DNA-RNA”, requires the sequencing of RNA and DNA from the same individual (Figure 3b). The comparison of Single Nucleotide Variations (SNVs) by matching DNA- and RNA-seq data with the reference genome allows the correct filtering out of pre-existing (genomic) SNVs, thus lowering the false positive rate. Various tools have been developed adopting different algorithms and statistical methods to maximize the accuracy of the analysis. Additionally, editing sites can be traced to “De Novo” by exploiting RNA-seq data only. In this case, the filtering step and the statistical approach are crucial for lowering the false positive rate (Figure 3c). Some of the most used tools for RES detection in Illumina sequencing data are summarized in Table 3.
Table 3.
Description of the different tools useful for tracing RES in Illumina sequencing data. The name, input datasets accepted and a brief description are reported for each tool considered.
In recent years, due to the expansion of the Machine Learning algorithm (ML), some machine learning techniques have also been developed to detect RES (Figure 3d).
RDDpred [27] is one of the first tools designed using an ML approach to predict RNA editing sites from RNA sequencing data. It employs a Random Forest algorithm to accurately distinguish between genuine RNA editing events and false positives. The tool constructs condition-specific training datasets by incorporating data from established RNA-editing databases. Additionally, RDDpred utilizes the Mapping Errors Set method to identify regions prone to alignment errors.
RED-ML [70] leverages information from various features and the properties of RES to make predictions using a logistic regression classifier for genome-wide RES identification. If DNA-seq data are available, SNPs can be specified and included in the analysis. This tool is limited to human RNA-seq data and can only identify sites with relatively high editing levels.
DeepRed [71] employs deep and ensemble learning to identify genuine RES from a list of traced SNVs. Thus, it can help validate a list of putative RNA editing sites.
Finally, the advent of long-read sequencing technology, such as Pacific Biosciences (PacBio) and Oxford Nanopore Technology (ONT), opened the avenue for new tools to exploit the potential of these novel types of data (Figure 3e). In particular, the possibility of sequencing native RNA by the direct RNA (dRNA) protocol of ONT theoretically makes the direct detection of inosine in RNA molecules possible.
Dinopore [72] is a deep learning-based tool designed to detect RNA editing sites, specifically A-to-I editing, using nanopore direct RNA sequencing data.
DeepEdit [73] is a cutting-edge tool designed to detect A-to-I RNA editing events using nanopore direct RNA sequencing data. This tool leverages a fully connected neural network model to analyze the raw electrical signals generated during nanopore sequencing.
L-GIREMI [74] is another tool for detecting and analyzing RNA editing sites within long-read RNA-seq data. It adeptly manages sequencing errors and read biases and demonstrates high accuracy when applied to PacBio RNA-seq data.
Overall, both the ML and long-reads approaches present some limitations; namely, both strategies require good bioinformatic knowledge and considerable computational effort. In addition, these techniques are accurate for species with well-curated reference genomes, whereas they could be somewhat inaccurate if the reference genome used is not of high-quality. Moreover, for ML algorithms, the training sets mostly derive from model organisms, lowering the sensitivity and accuracy if applied to phylogenetically distant species.
3.3. RNA Editing Sites Filtering
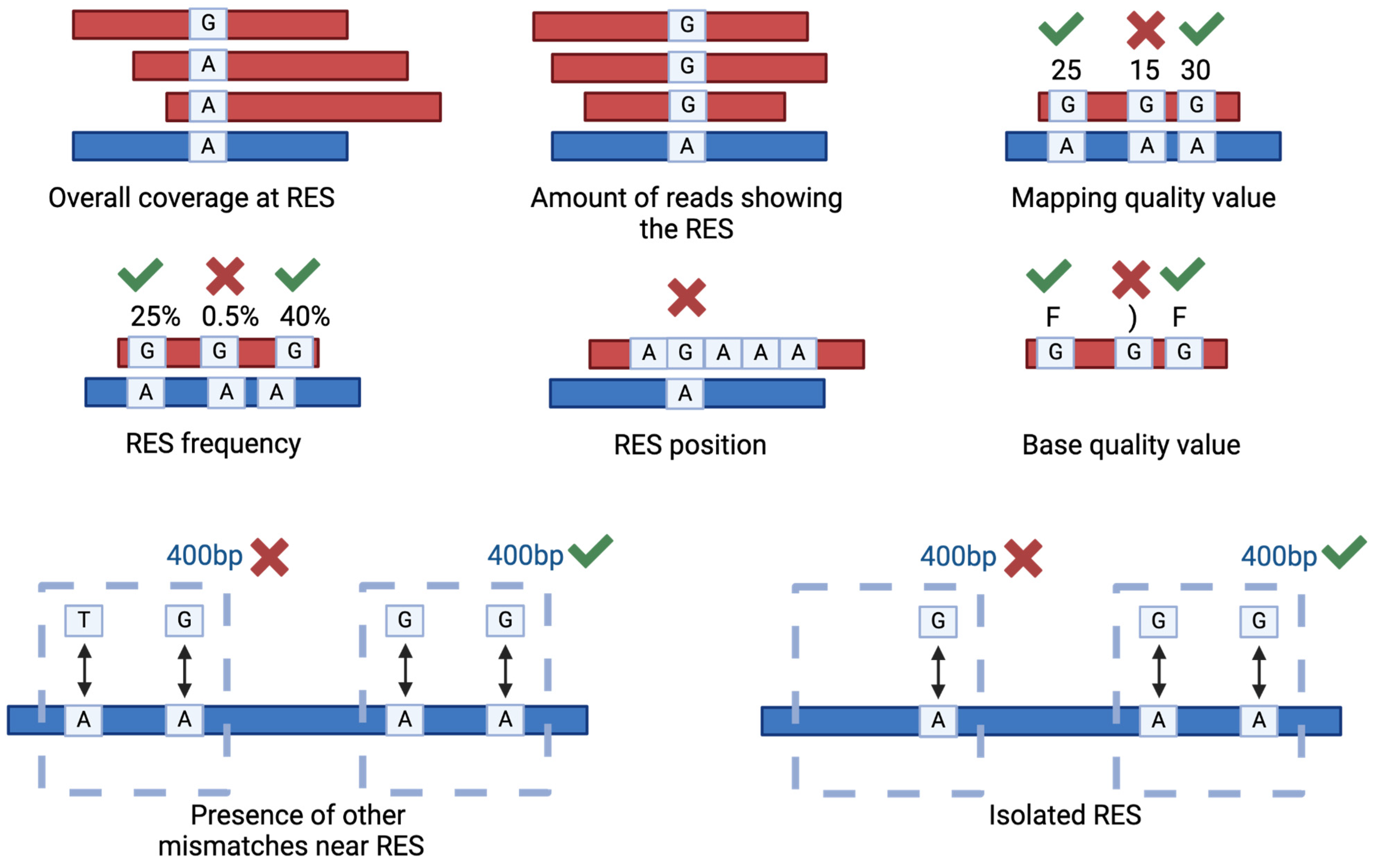
Several parameters can be used to filter the putative RES in order to increase the resolution and the accuracy of the analysis (Figure 4). The most used parameters are read coverage, amount of reads supporting the RES, mapping quality, RES frequency and position. As an example, it is a good practice to exclude RES occurring in homopolymeric regions [58]. Sometimes, these parameters are insufficient for obtaining an acceptable false positive rate. For instance, in the case of zebrafish and Xenopus sp., even if using matching DNA- and RNA-seq data, only with additional filters were the authors able to identify high-quality RES [75,76]. These additional filters are based on the sequence position of the putative RES in comparison to other putative RES or to different types of mismatches [75]. Specifically, since in most of the cases the editing sites are clustered together, the RES were excluded from the results if another non-editing compatible mismatch was traced in a 400 bp genomic window, and if the RES was isolated (no other A-to-G variations in a 400 bp genomic window), then the genomic window can be adapted to match specific organisms [75].
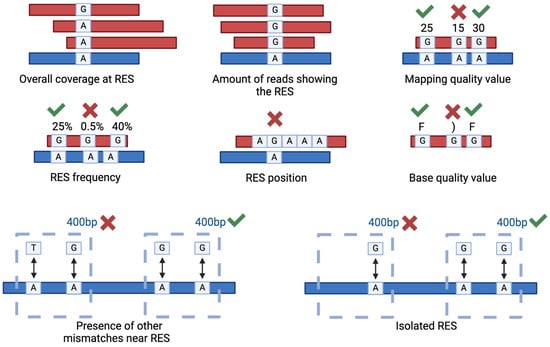
Figure 4.
Main filtering criteria used to select genuine RNA Editing Sites (RES). The picture illustrates the main parameters used to filter the RES. The parameters include the overall coverage at RES, the number of reads displaying the RES, the associated mapping quality value, the frequency and position of the RES and the base quality values. Low quality values are, as examples, 15 or “)”, good quality ones are 30 or “F”. In addition, two strong filters are represented by the relative position of the identified RES compared to other putative RES or different types of mismatches.
4. RNA Editing Indexes
RNA editing indexes are essential for rapidly evaluating the extension of RNA editing in a given sample. Indeed, they provide the quantification of RNA editing events. Considering that most of the RNA editing in humans occurred in Alu repeats [57,58,77] and that ADAR targets the majority of Alu adenosines to some extent, one of the first RNA editing indexes was developed by considering the RES in the Alu repeats. The so-called Alu Editing Index (AEI), developed in 2019, is based on the ratio of the number of A-to-G mismatches to the total coverage of adenosines (that is, the sum of the number of A-to-G RNA-DNA mismatches and A-A matches in these regions) [78]. Correctly tracing the RNA editing sites requires considerable computation time and power, which is why the main advantage of AEI is that it allows rapid estimation of the editing level, as well as the considering of a large number of samples. However, since Alu repeats are primate-exclusive, this index cannot be used for other species as it is. However, modification of this index was proposed in order to investigate RNA editing in zebrafish by modifying the repeat type [75]. Further modification will allow us to exploit the RNA editing index in other species.
Another index is represented by the overall editing, defined as the total number of reads with G at all known editing positions over the number of all reads covering the positions, without imposing specific sequencing coverage criteria [57]. It can be calculated using REDItools tables. Nevertheless, this index requires performing the RES calling and a database containing described RES, making it inapplicable to species not covered in these databases.
Finally, the overall editing calculated at recoding positions, namely the editing events located in coding genomic regions, is referred to as the recoding index (REI) [79]. This metric, used to investigate the activity of ADAR2, can be calculated using REDItools tables and a list of recoding sites from a RES database.
5. Conclusions
ADAR-mediated RNA editing is an essential biological process that can result in the alteration of protein sequences or in the modulation of mRNAs, and its analysis can contribute to improving our understanding of organisms’ homeostasis, disease and development processes. Understanding the connection between RNA editing and the fate of RNA in different species and conditions will probably untangle intimate regulative processes, which can be evolutionary conserved or species-specific. Because RNA editing can impact various layers of cellular regulation, integrating different types of omics data (such as genomics, transcriptomics, proteomics, and metabolomics) could provide a comprehensive view of the functional consequences and regulatory networks promoted by ADAR enzymatic and non-enzymatic functions. The integration of transcriptomic and genomics data has been widely used to distinguish between real RES and genomic mutations. In this context, proteomic data could provide an additional layer of validation, revealing the consequences of ADAR editing on protein synthesis. However, the integration of these omics requires sophisticated bioinformatic tools and statistical methods to manage and interpret the data appropriately. Therefore, despite its potential, multi-omics integration faces challenges such as data heterogeneity, the need for large sample sizes to achieve statistical power and computational complexity of data analysis.
Appropriate bioinformatic pipelines are needed to fully exploit datasets produced with existing technologies, as well as with novel emerging technologies able to provide a single-base resolution of any kind of RNA modification. Undoubtedly, applying neural networks and AI-based algorithms can contribute to developing novel tools. However, the requirement of an appropriate training dataset is the current challenge for their broad application. At the same time, the advancement of the accuracy of long-read sequencing and of protocols to sequence native RNA molecules could drastically contribute to the detection of inosine.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes15080996/s1, Supplementary Table S1. PTM detection tools. For each tool, the targeted RNA modifications, the algorithm used, the year of release and the associated publication are reported [80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157].
Author Contributions
Conceptualization, E.B. and U.R.; formal analysis, E.B.; resources, U.R.; writing—original draft preparation, E.B.; writing—review and editing, E.B and U.R.; supervision, U.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Italian Ministry of University and Research (MIUR), PRIN projects 202292P4R7 and P2022JEEMT.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cohn, W.E. Pseudouridine, a Carbon-Carbon Linked Ribonucleoside in Ribonucleic Acids: Isolation, Structure, and Chemical Characteristics. J. Biol. Chem. 1960, 235, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, R.J.; Stoute, J.; Fange Liu, K. The Chemical Diversity of RNA Modifications. Biochem. J. 2019, 476, 1227–1245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, L.; Li, X. Detection Technologies for RNA Modifications. Exp. Mol. Med. 2022, 54, 1601–1616. [Google Scholar] [CrossRef] [PubMed]
- Boo, S.H.; Kim, Y.K. The Emerging Role of RNA Modifications in the Regulation of mRNA Stability. Exp. Mol. Med. 2020, 52, 400–408. [Google Scholar] [CrossRef]
- Srinivas, K.P.; Depledge, D.P.; Abebe, J.S.; Rice, S.A.; Mohr, I.; Wilson, A.C. Widespread Remodeling of the m6A RNA-Modification Landscape by a Viral Regulator of RNA Processing and Export. Proc. Natl. Acad. Sci. USA 2021, 118, e2104805118. [Google Scholar] [CrossRef]
- Cui, L.; Ma, R.; Cai, J.; Guo, C.; Chen, Z.; Yao, L.; Wang, Y.; Fan, R.; Wang, X.; Shi, Y. RNA Modifications: Importance in Immune Cell Biology and Related Diseases. Signal Transduct. Target. Ther. 2022, 7, 334. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.; Emeson, R.B. RNA Editing. Annu. Rev. Neurosci. 1996, 19, 27–52. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M.; Kapoor, U.; Jantsch, M.F. Understanding RNA Modifications: The Promises and Technological Bottlenecks of the ‘Epitranscriptome’. Open Biol. 2017, 7, 170077. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. A-to-I RNA Editing—Immune Protector and Transcriptome Diversifier. Nat. Rev. Genet. 2018, 19, 473–490. [Google Scholar] [CrossRef]
- Hartner, J.C.; Walkley, C.R.; Lu, J.; Orkin, S.H. ADAR1 Is Essential for the Maintenance of Hematopoiesis and Suppression of Interferon Signaling. Nat. Immunol. 2009, 10, 109–115. [Google Scholar] [CrossRef]
- Hwang, T.; Park, C.-K.; Leung, A.K.L.; Gao, Y.; Hyde, T.M.; Kleinman, J.E.; Rajpurohit, A.; Tao, R.; Shin, J.H.; Weinberger, D.R. Dynamic Regulation of RNA Editing in Human Brain Development and Disease. Nat. Neurosci. 2016, 19, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.J.; Shi, H.; He, C. Epitranscriptomic Influences on Development and Disease. Genome Biol. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Holley, R.W.; Apgar, J.; Everett, G.A.; Madison, J.T.; Marquisee, M.; Merrill, S.H.; Penswick, J.R.; Zamir, A. Structure of a Ribonucleic Acid. Science 1965, 147, 1462–1465. [Google Scholar] [CrossRef] [PubMed]
- Ashley, C.N.; Broni, E.; Miller, W.A. ADAR Family Proteins: A Structural Review. Curr. Issues Mol. Biol. 2024, 46, 3919–3945. [Google Scholar] [CrossRef]
- Saletore, Y.; Meyer, K.; Korlach, J.; Vilfan, I.D.; Jaffrey, S.; Mason, C.E. The Birth of the Epitranscriptome: Deciphering the Function of RNA Modifications. Genome Biol. 2012, 13, 175. [Google Scholar] [CrossRef]
- Bass, B. A Developmentally Regulated Activity That Unwinds RNA Duplexes. Cell 1987, 48, 607–613. [Google Scholar] [CrossRef]
- Rebagliati, M.R.; Melton, D.A. Antisense RNA Injections in Fertilized Frog Eggs Reveal an RNA Duplex Unwinding Activity. Cell 1987, 48, 599–605. [Google Scholar] [CrossRef]
- Bass, B.L.; Weintraub, H. An Unwinding Activity That Covalently Modifies Its Double-Stranded RNA Substrate. Cell 1988, 55, 1089–1098. [Google Scholar] [CrossRef]
- Hartner, J.C.; Schmittwolf, C.; Kispert, A.; Müller, A.M.; Higuchi, M.; Seeburg, P.H. Liver Disintegration in the Mouse Embryo Caused by Deficiency in the RNA-Editing Enzyme ADAR1. J. Biol. Chem. 2004, 279, 4894–4902. [Google Scholar] [CrossRef]
- Sommer, B.; Köhler, M.; Sprengel, R.; Seeburg, P.H. RNA Editing in Brain Controls a Determinant of Ion Flow in Glutamate-Gated Channels. Cell 1991, 67, 11–19. [Google Scholar] [CrossRef]
- Levanon, E.Y.; Eisenberg, E. Does RNA Editing Compensate for Alu Invasion of the Primate Genome? BioEssays 2015, 37, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.P.; Bass, B.L. Detection of Inosine in Messenger RNA by Inosine-Specific Cleavage. Biochemistry 1997, 36, 8429–8434. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, G.; Lin, W.; Piskol, R.; Tan, M.H.; Davis, C.; Li, J.B. Accurate Identification of Human Alu and Non-Alu RNA Editing Sites. Nat. Methods 2012, 9, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, G.; Li, J.B. RADAR: A Rigorously Annotated Database of A-to-I RNA Editing. Nucleic Acids Res. 2014, 42, D109–D113. [Google Scholar] [CrossRef] [PubMed]
- Porath, H.T.; Carmi, S.; Levanon, E.Y. A Genome-Wide Map of Hyper-Edited RNA Reveals Numerous New Sites. Nat. Commun. 2014, 5, 4726. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Hur, B.; Kim, S. RDDpred: A Condition-Specific RNA-Editing Prediction Model from RNA-Seq Data. BMC Genom. 2016, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and Regulation of RNA Editing by ADAR Deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) Project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Wang, Y.; Chung, D.H.; Monteleone, L.R.; Li, J.; Chiang, Y.; Toney, M.D.; Beal, P.A. RNA Binding Candidates for Human ADAR3 from Substrates of a Gain of Function Mutant Expressed in Neuronal Cells. Nucleic Acids Res. 2019, 47, 10801–10814. [Google Scholar] [CrossRef]
- Picardi, E.; Pesole, G. REDItools: High-Throughput RNA Editing Detection Made Easy. Bioinformatics 2013, 29, 1813–1814. [Google Scholar] [CrossRef] [PubMed]
- Porath, H.T.; Knisbacher, B.A.; Eisenberg, E.; Levanon, E.Y. Massive A-to-I RNA Editing Is Common across the Metazoa and Correlates with dsRNA Abundance. Genome Biol. 2017, 18, 185. [Google Scholar] [CrossRef] [PubMed]
- Suspène, R.; Renard, M.; Henry, M.; Guétard, D.; Puyraimond-Zemmour, D.; Billecocq, A.; Bouloy, M.; Tangy, F.; Vartanian, J.-P.; Wain-Hobson, S. Inversing the Natural Hydrogen Bonding Rule to Selectively Amplify GC-Rich ADAR-Edited RNAs. Nucleic Acids Res. 2008, 36, e72. [Google Scholar] [CrossRef] [PubMed]
- Licht, K.; Hartl, M.; Amman, F.; Anrather, D.; Janisiw, M.P.; Jantsch, M.F. Inosine Induces Context-Dependent Recoding and Translational Stalling. Nucleic Acids Res. 2019, 47, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Maas, S.; Single, F.N.; Hartner, J.; Rozov, A.; Burnashev, N.; Feldmeyer, D.; Sprengel, R.; Seeburg, P.H. Point Mutation in an AMPA Receptor Gene Rescues Lethality in Mice De®cient in the RNA-Editing Enzyme ADAR2. Nature 2000, 406, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Li, I.-C.; Chen, Y.-C.; Wang, Y.-Y.; Tzeng, B.-W.; Ou, C.-W.; Lau, Y.-Y.; Wu, K.-M.; Chan, T.-M.; Lin, W.-H.; Hwang, S.-P.L.; et al. Zebrafish Adar2 Edits the Q/R Site of AMPA Receptor Subunit Gria2α Transcript to Ensure Normal Development of Nervous System and Cranial Neural Crest Cells. PLoS ONE 2014, 9, e97133. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, M.; Carlson, J.W.; Celniker, S.E. RNA Editing in Drosophila melanogaster: New Targets and Functional Consequences. RNA 2006, 12, 1922–1932. [Google Scholar] [CrossRef]
- Alon, S.; Garrett, S.C.; Levanon, E.Y.; Olson, S.; Graveley, B.R.; Rosenthal, J.J.C.; Eisenberg, E. The Majority of Transcripts in the Squid Nervous System Are Extensively Recoded by A-to-I RNA Editing. eLife 2015, 4, e05198. [Google Scholar] [CrossRef]
- Rangan, K.J.; Reck-Peterson, S.L. RNA Recoding in Cephalopods Tailors Microtubule Motor Protein Function. Cell 2023, 186, 2531–2543. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Rich, A.; Maas, S. Widespread A-to-I RNA Editing of Alu-Containing mRNAs in the Human Transcriptome. PLoS Biol. 2004, 2, e391. [Google Scholar] [CrossRef]
- Neeman, Y.; Levanon, E.Y.; Jantsch, M.F.; Eisenberg, E. RNA Editing Level in the Mouse Is Determined by the Genomic Repeat Repertoire. RNA 2006, 12, 1802–1809. [Google Scholar] [CrossRef]
- Bass, B.L. Adenosine Deaminases That Act on RNA, Then and Now. RNA 2024, 30, 521–529. [Google Scholar] [CrossRef]
- Baquero-Pérez, B.; Bortoletto, E.; Rosani, U.; Delgado-Tejedor, A.; Medina, R.; Novoa, E.M.; Venier, P.; Díez, J. Elucidation of the Epitranscriptomic RNA Modification Landscape of Chikungunya Virus. Viruses 2024, 16, 945. [Google Scholar] [CrossRef]
- Min, Y.-H.; Shao, W.-X.; Hu, Q.-S.; Xie, N.-B.; Zhang, S.; Feng, Y.-Q.; Xing, X.-W.; Yuan, B.-F. Simultaneous Detection of Adenosine-to-Inosine Editing and N6-Methyladenosine at Identical RNA Sites through Deamination-Assisted Reverse Transcription Stalling. Anal. Chem. 2024, 96, 8730–8739. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, M.; Suzuki, T. Biochemical Identification of A-to-I RNA Editing Sites by the Inosine Chemical Erasing (ICE) Method. In RNA and DNA Editing: Methods and Protocols; Aphasizhev, R., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 89–99. ISBN 978-1-61779-018-8. [Google Scholar]
- Ding, J.-H.; Chen, M.-Y.; Xie, N.-B.; Xie, C.; Xiong, N.; He, J.-G.; Wang, J.; Guo, C.; Feng, Y.-Q.; Yuan, B.-F. Quantitative and Site-Specific Detection of Inosine Modification in RNA by Acrylonitrile Labeling-Mediated Elongation Stalling. Biosens. Bioelectron. 2023, 219, 114821. [Google Scholar] [CrossRef]
- Chen, J.-J.; You, X.-J.; Li, L.; Xie, N.-B.; Ding, J.-H.; Yuan, B.-F.; Feng, Y.-Q. Single-Base Resolution Detection of Adenosine-to-Inosine RNA Editing by Endonuclease-Mediated Sequencing. Anal. Chem. 2022, 94, 8740–8747. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA Sequencing with Chain-Terminating Inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality Control of RNA-Seq Experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef]
- DeLuca, D.S.; Levin, J.Z.; Sivachenko, A.; Fennell, T.; Nazaire, M.-D.; Williams, C.; Reich, M.; Winckler, W.; Getz, G. RNA-SeQC: RNA-Seq Metrics for Quality Control and Process Optimization. Bioinformatics 2012, 28, 1530–1532. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Jiang, H.; Lei, R.; Ding, S.-W.; Zhu, S. Skewer: A Fast and Accurate Adapter Trimmer for next-Generation Sequencing Paired-End Reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef]
- Sahlin, K.; Baudeau, T.; Cazaux, B.; Marchet, C. A Survey of Mapping Algorithms in the Long-Reads Era. Genome Biol. 2023, 24, 133. [Google Scholar] [CrossRef]
- Lo Giudice, C.; Silvestris, D.A.; Roth, S.H.; Eisenberg, E.; Pesole, G.; Gallo, A.; Picardi, E. Quantifying RNA Editing in Deep Transcriptome Datasets. Front. Genet. 2020, 11, 194. [Google Scholar] [CrossRef]
- Lo Giudice, C.; Tangaro, M.A.; Pesole, G.; Picardi, E. Investigating RNA Editing in Deep Transcriptome Datasets with REDItools and REDIportal. Nat. Protoc. 2020, 15, 1098–1131. [Google Scholar] [CrossRef]
- Picardi, E.; Pesole, G. (Eds.) RNA Editing: Methods and Protocols; Methods in Molecular Biology; Springer US: New York, NY, USA, 2021; Volume 2181, ISBN 978-1-07-160786-2. [Google Scholar]
- Morales, D.R.; Rennie, S.; Uchida, S. Benchmarking RNA Editing Detection Tools. BioTech 2023, 12, 56. [Google Scholar] [CrossRef]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A Framework for Variation Discovery and Genotyping Using Next-Generation DNA Sequencing Data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to High Confidence Variant Calls: The Genome Analysis Toolkit Best Practices Pipeline. Curr. Protoc. Bioinforma. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Picardi, E.; D’Erchia, A.M.; Lo Giudice, C.; Pesole, G. REDIportal: A Comprehensive Database of A-to-I RNA Editing Events in Humans. Nucleic Acids Res. 2017, 45, D750–D757. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lian, J.; Li, Q.; Zhang, P.; Zhou, Y.; Zhan, X.; Zhang, G. RES-Scanner: A Software Package for Genome-Wide Identification of RNA-Editing Sites. GigaScience 2016, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Piechotta, M.; Wyler, E.; Ohler, U.; Landthaler, M.; Dieterich, C. JACUSA: Site-Specific Identification of RNA Editing Events from Replicate Sequencing Data. BMC Bioinform. 2017, 18, 7. [Google Scholar] [CrossRef]
- Light, D.; Haas, R.; Yazbak, M.; Elfand, T.; Blau, T.; Lamm, A.T. RESIC: A Tool for Comprehensive Adenosine to Inosine RNA Editing Site Identification and Classification. Front. Genet. 2021, 12, 686851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, X. Genome Sequence–Independent Identification of RNA Editing Sites. Nat. Methods 2015, 12, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lu, Y.; Yan, S.; Xing, Q.; Tian, W. SPRINT: An SNP-Free Toolkit for Identifying RNA Editing Sites. Bioinformatics 2017, 33, 3538–3548. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Liu, D.; Li, Q.; Lei, M.; Xu, L.; Wu, L.; Wang, Z.; Ren, S.; Li, W.; Xia, M.; et al. RED-ML: A Novel, Effective RNA Editing Detection Method Based on Machine Learning. GigaScience 2017, 6, gix012. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Liu, F.; Zhao, C.; Ren, C.; An, G.; Mei, C.; Bo, X.; Shu, W. Accurate Identification of RNA Editing Sites from Primitive Sequence with Deep Neural Networks. Sci. Rep. 2018, 8, 6005. [Google Scholar] [CrossRef]
- Nguyen, T.A.; Heng, J.W.J.; Kaewsapsak, P.; Kok, E.P.L.; Stanojević, D.; Liu, H.; Cardilla, A.; Praditya, A.; Yi, Z.; Lin, M.; et al. Direct Identification of A-to-I Editing Sites with Nanopore Native RNA Sequencing. Nat. Methods 2022, 19, 833–844. [Google Scholar] [CrossRef]
- Chen, L.; Ou, L.; Jing, X.; Kong, Y.; Xie, B.; Zhang, N.; Shi, H.; Qin, H.; Li, X.; Hao, P. DeepEdit: Single-Molecule Detection and Phasing of A-to-I RNA Editing Events Using Nanopore Direct RNA Sequencing. Genome Biol. 2023, 24, 75. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Quinones-Valdez, G.; Fu, T.; Huang, E.; Choudhury, M.; Reese, F.; Mortazavi, A.; Xiao, X. L-GIREMI Uncovers RNA Editing Sites in Long-Read RNA-Seq. Genome Biol. 2023, 24, 171. [Google Scholar] [CrossRef] [PubMed]
- Buchumenski, I.; Holler, K.; Appelbaum, L.; Eisenberg, E.; Junker, J.P.; Levanon, E.Y. Systematic Identification of A-to-I RNA Editing in Zebrafish Development and Adult Organs. Nucleic Acids Res. 2021, 49, 4325–4337. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.A.; Heng, J.W.J.; Ng, Y.T.; Sun, R.; Fisher, S.; Oguz, G.; Kaewsapsak, P.; Xue, S.; Reversade, B.; Ramasamy, A.; et al. Deep Transcriptome Profiling Reveals Limited Conservation of A-to-I RNA Editing in Xenopus. BMC Biol. 2023, 21, 251. [Google Scholar] [CrossRef] [PubMed]
- Levanon, E.Y.; Eisenberg, E.; Yelin, R.; Nemzer, S.; Hallegger, M.; Shemesh, R.; Fligelman, Z.Y.; Shoshan, A.; Pollock, S.R.; Sztybel, D.; et al. Systematic Identification of Abundant A-to-I Editing Sites in the Human Transcriptome. Nat. Biotechnol. 2004, 22, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.H.; Levanon, E.Y.; Eisenberg, E. Genome-Wide Quantification of ADAR Adenosine-to-Inosine RNA Editing Activity. Nat. Methods 2019, 16, 1131–1138. [Google Scholar] [CrossRef]
- Silvestris, D.A.; Picardi, E.; Cesarini, V.; Fosso, B.; Mangraviti, N.; Massimi, L.; Martini, M.; Pesole, G.; Locatelli, F.; Gallo, A. Dynamic Inosinome Profiles Reveal Novel Patient Stratification and Gender-Specific Differences in Glioblastoma. Genome Biol. 2019, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, P.; Ding, H.; Lin, H.; Chou, K.-C. iRNA-Methyl: Identifying N(6)-Methyladenosine Sites Using Pseudo Nucleotide Composition. Anal. Biochem. 2015, 490, 26–33. [Google Scholar] [CrossRef]
- Chen, W.; Tran, H.; Liang, Z.; Lin, H.; Zhang, L. Identification and Analysis of the N(6)-Methyladenosine in the Saccharomyces Cerevisiae Transcriptome. Sci. Rep. 2015, 5, 13859. [Google Scholar] [CrossRef]
- Li, Y.-H.; Zhang, G.; Cui, Q. PPUS: A Web Server to Predict PUS-Specific Pseudouridine Sites. Bioinforma. Oxf. Engl. 2015, 31, 3362–3364. [Google Scholar] [CrossRef]
- Chen, W.; Feng, P.; Tang, H.; Ding, H.; Lin, H. RAMPred: Identifying the N(1)-Methyladenosine Sites in Eukaryotic Transcriptomes. Sci. Rep. 2016, 6, 31080. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, P.; Ding, H.; Lin, H. Identifying N 6-Methyladenosine Sites in the Arabidopsis Thaliana Transcriptome. Mol. Genet. Genom. MGG 2016, 291, 2225–2229. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.-Z.; Zhang, J.-J.; Gu, W.-Z. RNA-MethylPred: A High-Accuracy Predictor to Identify N6-Methyladenosine in RNA. Anal. Biochem. 2016, 510, 72–75. [Google Scholar] [CrossRef]
- Li, G.-Q.; Liu, Z.; Shen, H.-B.; Yu, D.-J. TargetM6A: Identifying N6-Methyladenosine Sites From RNA Sequences via Position-Specific Nucleotide Propensities and a Support Vector Machine. IEEE Trans. Nanobiosci. 2016, 15, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xiao, X.; Yu, D.-J.; Jia, J.; Qiu, W.-R.; Chou, K.-C. pRNAm-PC: Predicting N(6)-Methyladenosine Sites in RNA Sequences via Physical-Chemical Properties. Anal. Biochem. 2016, 497, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liu, K.; Yan, Z.; Zhang, Y.; Sun, Z. RNAMethPre: A Web Server for the Prediction and Query of mRNA m6A Sites. PLoS ONE 2016, 11, e0162707. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Yan, Z.; Liu, K.; Zhang, Y.; Sun, Z. AthMethPre: A Web Server for the Prediction and Query of mRNA m6A Sites in Arabidopsis Thaliana. Mol. Biosyst. 2016, 12, 3333–3337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, J.-W.; Liu, Z.; Ren, M.-W.; Shen, H.-B.; Yu, D.-J. Improving N(6)-Methyladenosine Site Prediction with Heuristic Selection of Nucleotide Physical-Chemical Properties. Anal. Biochem. 2016, 508, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zeng, P.; Li, Y.-H.; Zhang, Z.; Cui, Q. SRAMP: Prediction of Mammalian N6-Methyladenosine (m6A) Sites Based on Sequence-Derived Features. Nucleic Acids Res. 2016, 44, e91. [Google Scholar] [CrossRef]
- Chen, W.; Feng, P.; Tang, H.; Ding, H.; Lin, H. Identifying 2’-O-Methylationation Sites by Integrating Nucleotide Chemical Properties and Nucleotide Compositions. Genomics 2016, 107, 255–258. [Google Scholar] [CrossRef]
- Chen, W.; Tang, H.; Ye, J.; Lin, H.; Chou, K.-C. iRNA-PseU: Identifying RNA Pseudouridine Sites. Mol. Ther. Nucleic Acids 2016, 5, e332. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.-R.; Jiang, S.-Y.; Xu, Z.-C.; Xiao, X.; Chou, K.-C. iRNAm5C-PseDNC: Identifying RNA 5-Methylcytosine Sites by Incorporating Physical-Chemical Properties into Pseudo Dinucleotide Composition. Oncotarget 2017, 8, 41178–41188. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Ding, H.; Yang, H.; Chen, W.; Lin, H.; Chou, K.-C. iRNA-PseColl: Identifying the Occurrence Sites of Different RNA Modifications by Incorporating Collective Effects of Nucleotides into PseKNC. Mol. Ther. Nucleic Acids 2017, 7, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Tang, H.; Lin, H. MethyRNA: A Web Server for Identification of N6-Methyladenosine Sites. J. Biomol. Struct. Dyn. 2017, 35, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xing, P.; Zou, Q. Detecting N6-Methyladenosine Sites from RNA Transcriptomes Using Ensemble Support Vector Machines. Sci. Rep. 2017, 7, 40242. [Google Scholar] [CrossRef] [PubMed]
- Xing, P.; Su, R.; Guo, F.; Wei, L. Identifying N6-Methyladenosine Sites Using Multi-Interval Nucleotide Pair Position Specificity and Support Vector Machine. Sci. Rep. 2017, 7, 46757. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, P.; Yang, H.; Ding, H.; Lin, H.; Chou, K.-C. iRNA-3typeA: Identifying Three Types of Modification at RNA’s Adenosine Sites. Mol. Ther. Nucleic Acids 2018, 11, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhai, J.; Bian, E.; Song, Y.; Yu, J.; Ma, C. Transcriptome-Wide Annotation of m5C RNA Modifications Using Machine Learning. Front. Plant Sci. 2018, 9, 519. [Google Scholar] [CrossRef]
- Sabooh, M.F.; Iqbal, N.; Khan, M.; Khan, M.; Maqbool, H.F. Identifying 5-Methylcytosine Sites in RNA Sequence Using Composite Encoding Feature into Chou’s PseKNC. J. Theor. Biol. 2018, 452, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Huang, Y.; Yang, X.; Zhou, Y.; Zhou, Y. RNAm5Cfinder: A Web-Server for Predicting RNA 5-Methylcytosine (m5C) Sites Based on Random Forest. Sci. Rep. 2018, 8, 17299. [Google Scholar] [CrossRef]
- Akbar, S.; Hayat, M. iMethyl-STTNC: Identification of N6-Methyladenosine Sites by Extending the Idea of SAAC into Chou’s PseAAC to Formulate RNA Sequences. J. Theor. Biol. 2018, 455, 205–211. [Google Scholar] [CrossRef]
- Chen, W.; Ding, H.; Zhou, X.; Lin, H.; Chou, K.-C. iRNA(m6A)-PseDNC: Identifying N6-Methyladenosine Sites Using Pseudo Dinucleotide Composition. Anal. Biochem. 2018, 561–562, 59–65. [Google Scholar] [CrossRef]
- Zhao, Z.; Peng, H.; Lan, C.; Zheng, Y.; Fang, L.; Li, J. Imbalance Learning for the Prediction of N6-Methylation Sites in mRNAs. BMC Genomics 2018, 19, 574. [Google Scholar] [CrossRef]
- Wei, L.; Chen, H.; Su, R. M6APred-EL: A Sequence-Based Predictor for Identifying N6-Methyladenosine Sites Using Ensemble Learning. Mol. Ther. Nucleic Acids 2018, 12, 635–644. [Google Scholar] [CrossRef]
- Huang, Y.; He, N.; Chen, Y.; Chen, Z.; Li, L. BERMP: A Cross-Species Classifier for Predicting m6A Sites by Integrating a Deep Learning Algorithm and a Random Forest Approach. Int. J. Biol. Sci. 2018, 14, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Qiang, X.; Chen, H.; Ye, X.; Su, R.; Wei, L. M6AMRFS: Robust Prediction of N6-Methyladenosine Sites With Sequence-Based Features in Multiple Species. Front. Genet. 2018, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, R. RFAthM6A: A New Tool for Predicting m6A Sites in Arabidopsis Thaliana. Plant Mol. Biol. 2018, 96, 327–337. [Google Scholar] [CrossRef]
- Yang, H.; Lv, H.; Ding, H.; Chen, W.; Lin, H. iRNA-2OM: A Sequence-Based Predictor for Identifying 2’-O-Methylation Sites in Homo Sapiens. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2018, 25, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cui, Q.; Zhou, Y. NmSEER: A Prediction Tool for 2’-O-Methylation (Nm) Sites Based on Random Forest. In Proceedings of the Intelligent Computing Theories and Application: 14th International Conference, ICIC 2018, Wuhan, China, 15–18 August 2018; Proceedings, Part I. Springer-Verlag: Berlin, Heidelberg, 2018; pp. 893–900. [Google Scholar]
- He, J.; Fang, T.; Zhang, Z.; Huang, B.; Zhu, X.; Xiong, Y. PseUI: Pseudouridine Sites Identification Based on RNA Sequence Information. BMC Bioinform. 2018, 19, 306. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Zhang, Z.; Sun, R.; Zhu, L.; He, J.; Huang, B.; Xiong, Y.; Zhu, X. RNAm5CPred: Prediction of RNA 5-Methylcytosine Sites Based on Three Different Kinds of Nucleotide Composition. Mol. Ther. Nucleic Acids 2019, 18, 739–747. [Google Scholar] [CrossRef]
- Sun, P.P.; Chen, Y.B.; Liu, B.; Gao, Y.X.; Han, Y.; He, F.; Ji, J.C. DeepMRMP: A New Predictor for Multiple Types of RNA Modification Sites Using Deep Learning. Math. Biosci. Eng. MBE 2019, 16, 6231–6241. [Google Scholar] [CrossRef]
- Wei, L.; Su, R.; Wang, B.; Li, X.; Zou, Q.; Gao, X. Integration of Deep Feature Representations and Handcrafted Features to Improve the Prediction of N6-Methyladenosine Sites. Neurocomputing 2019, 324, 3–9. [Google Scholar] [CrossRef]
- Zou, Q.; Xing, P.; Wei, L.; Liu, B. Gene2vec: Gene Subsequence Embedding for Prediction of Mammalian N6-Methyladenosine Sites from mRNA. RNA N. Y. N 2019, 25, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-Y.; Zhang, S.-W.; Fan, X.-N.; Zhang, T.; Meng, J.; Huang, Y. FunDMDeep-m6A: Identification and Prioritization of Functional Differential m6A Methylation Genes. Bioinformatics 2019, 35, i90–i98. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, P.; Song, X.; Lv, H.; Lin, H. iRNA-m7G: Identifying N7-Methylguanosine Sites by Fusing Multiple Features. Mol. Ther. Nucleic Acids 2019, 18, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Tayara, H.; Chong, K.T. iRNA-PseKNC(2methyl): Identify RNA 2’-O-Methylation Sites by Convolution Neural Network and Chou’s Pseudo Components. J. Theor. Biol. 2019, 465, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cui, Q.; Zhou, Y. NmSEER V2.0: A Prediction Tool for 2’-O-Methylation Sites Based on Random Forest and Multi-Encoding Combination. BMC Bioinform. 2019, 20, 690. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Tayara, H.; Chong, K.T. iPseU-CNN: Identifying RNA Pseudouridine Sites Using Convolutional Neural Networks. Mol. Ther. Nucleic Acids 2019, 16, 463–470. [Google Scholar] [CrossRef]
- Nguyen-Vo, T.-H.; Nguyen, Q.H.; Do, T.T.T.; Nguyen, T.-N.; Rahardja, S.; Nguyen, B.P. iPseU-NCP: Identifying RNA Pseudouridine Sites Using Random Forest and NCP-Encoded Features. BMC Genom. 2019, 20, 971. [Google Scholar] [CrossRef]
- Chen, K.; Wei, Z.; Zhang, Q.; Wu, X.; Rong, R.; Lu, Z.; Su, J.; de Magalhães, J.P.; Rigden, D.J.; Meng, J. WHISTLE: A High-Accuracy Map of the Human N6-Methyladenosine (m6A) Epitranscriptome Predicted Using a Machine Learning Approach. Nucleic Acids Res. 2019, 47, e41. [Google Scholar] [CrossRef]
- Lv, H.; Zhang, Z.-M.; Li, S.-H.; Tan, J.-X.; Chen, W.; Lin, H. Evaluation of Different Computational Methods on 5-Methylcytosine Sites Identification. Brief. Bioinform. 2020, 21, 982–995. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, Y.; Liu, Y.; Chen, Y.; Bi, S.; Zhu, X. m5CPred-SVM: A Novel Method for Predicting m5C Sites of RNA. BMC Bioinformatics 2020, 21, 489. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Song, B.; Tang, Y.; Chen, K.; Wei, Z.; Meng, J. m5UPred: A Web Server for the Prediction of RNA 5-Methyluridine Sites from Sequences. Mol. Ther. Nucleic Acids 2020, 22, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, P.; Li, F.; Wang, Y.; Smith, A.I.; Webb, G.I.; Akutsu, T.; Baggag, A.; Bensmail, H.; Song, J. Comprehensive Review and Assessment of Computational Methods for Predicting RNA Post-Transcriptional Modification Sites from RNA Sequences. Brief. Bioinform. 2020, 21, 1676–1696. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Chen, W. iMRM: A Platform for Simultaneously Identifying Multiple Kinds of RNA Modifications. Bioinforma. Oxf. Engl. 2020, 36, 3336–3342. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, Z.; Mao, X.; Li, Q. m7GPredictor: An Improved Machine Learning-Based Model for Predicting Internal m7G Modifications Using Sequence Properties. Anal. Biochem. 2020, 609, 113905. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xiang, D.; Ge, Z.; Li, F.; Jia, C.; Song, J. An Interpretable Prediction Model for Identifying N7-Methylguanosine Sites Based on XGBoost and SHAP. Mol. Ther. Nucleic Acids 2020, 22, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Jin, D.; Jia, C. EnsemPseU: Identifying Pseudouridine Sites With an Ensemble Approach. IEEE Access 2020, 8, 79376–79382. [Google Scholar] [CrossRef]
- Song, B.; Tang, Y.; Wei, Z.; Liu, G.; Su, J.; Meng, J.; Chen, K. PIANO: A Web Server for Pseudouridine-Site (Ψ) Identification and Functional Annotation. Front. Genet. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Chen, K.; Tang, Y.; Ma, J.; Meng, J.; Wei, Z. PSI-MOUSE: Predicting Mouse Pseudouridine Sites From Sequence and Genome-Derived Features. Evol. Bioinforma. Online 2020, 16, 1176934320925752. [Google Scholar] [CrossRef]
- Liu, K.; Chen, W.; Lin, H. XG-PseU: An eXtreme Gradient Boosting Based Method for Identifying Pseudouridine Sites. Mol. Genet. Genomics MGG 2020, 295, 13–21. [Google Scholar] [CrossRef]
- Lv, Z.; Zhang, J.; Ding, H.; Zou, Q. RF-PseU: A Random Forest Predictor for RNA Pseudouridine Sites. Front. Bioeng. Biotechnol. 2020, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Hong, K.J.; Tayara, H.; Chong, K. to m6A-NeuralTool: Convolution Neural Tool for RNA N6-Methyladenosine Site Identification in Different Species. IEEE Access 2021, 9, 17779–17786. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, X.; Liu, M.; Xu, Z.; Liu, G. DNN-m6A: A Cross-Species Method for Identifying RNA N6-Methyladenosine Sites Based on Deep Neural Network with Multi-Information Fusion. Genes 2021, 12, 354. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Feng, P.; Cui, L.; Su, R.; Chen, W.; Wei, L. Iterative Feature Representation Algorithm to Improve the Predictive Performance of N7-Methylguanosine Sites. Brief. Bioinform. 2021, 22, bbaa278. [Google Scholar] [CrossRef] [PubMed]
- Ning, Q.; Sheng, M. m7G-DLSTM: Intergrating Directional Double-LSTM and Fully Connected Network for RNA N7-Methlguanosine Sites Prediction in Human. Chemom. Intell. Lab. Syst. 2021, 217, 104398. [Google Scholar] [CrossRef]
- Li, H.; Chen, L.; Huang, Z.; Luo, X.; Li, H.; Ren, J.; Xie, Y. DeepOMe: A Web Server for the Prediction of 2’-O-Me Sites Based on the Hybrid CNN and BLSTM Architecture. Front. Cell Dev. Biol. 2021, 9, 686894. [Google Scholar] [CrossRef] [PubMed]
- Begik, O.; Lucas, M.C.; Pryszcz, L.P.; Ramirez, J.M.; Medina, R.; Milenkovic, I.; Cruciani, S.; Liu, H.; Vieira, H.G.S.; Sas-Chen, A.; et al. Quantitative Profiling of Pseudouridylation Dynamics in Native RNAs with Nanopore Sequencing. Nat. Biotechnol. 2021, 39, 1278–1291. [Google Scholar] [CrossRef]
- Feng, P.; Chen, W. iRNA-m5U: A Sequence Based Predictor for Identifying 5-Methyluridine Modification Sites in Saccharomyces Cerevisiae. Methods San Diego Calif 2022, 203, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Rehman, H.U.; Habib, U.; Ijaz, U. m6A-Finder: Detecting m6A Methylation Sites from RNA Transcriptomes Using Physical and Statistical Properties Based Features. Comput. Biol. Chem. 2022, 97, 107640. [Google Scholar] [CrossRef]
- Shoombuatong, W.; Basith, S.; Pitti, T.; Lee, G.; Manavalan, B. THRONE: A New Approach for Accurate Prediction of Human RNA N7-Methylguanosine Sites. J. Mol. Biol. 2022, 434, 167549. [Google Scholar] [CrossRef]
- Ao, C.; Zou, Q.; Yu, L. NmRF: Identification of Multispecies RNA 2’-O-Methylation Modification Sites from RNA Sequences. Brief. Bioinform. 2022, 23, bbab480. [Google Scholar] [CrossRef] [PubMed]
- Hassan, D.; Acevedo, D.; Daulatabad, S.V.; Mir, Q.; Janga, S.C. Penguin: A Tool for Predicting Pseudouridine Sites in Direct RNA Nanopore Sequencing Data. Methods 2022, 203, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Ao, C.; Ye, X.; Sakurai, T.; Zou, Q.; Yu, L. m5U-SVM: Identification of RNA 5-Methyluridine Modification Sites Based on Multi-View Features of Physicochemical Features and Distributed Representation. BMC Biol. 2023, 21, 93. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chung, C.-R.; Huang, H.-D.; Lee, T.-Y. Identification of Species-Specific RNA N6-Methyladinosine Modification Sites from RNA Sequences. Brief. Bioinform. 2023, 24, bbac573. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Tayara, H.; Chong, K.T. DL-m6A: Identification of N6-Methyladenosine Sites in Mammals Using Deep Learning Based on Different Encoding Schemes. IEEE/ACM Trans. Comput. Biol. Bioinform. 2023, 20, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Zhao, Y.; Jin, J.; Qiao, J.; Wang, D.; Wang, Y.; Wei, L. Rm-LR: A Long-Range-Based Deep Learning Model for Predicting Multiple Types of RNA Modifications. Comput. Biol. Med. 2023, 164, 107238. [Google Scholar] [CrossRef] [PubMed]
- Soylu, N.N.; Sefer, E. BERT2OME: Prediction of 2’-O-Methylation Modifications From RNA Sequence by Transformer Architecture Based on BERT. IEEE/ACM Trans. Comput. Biol. Bioinform. 2023, 20, 2177–2189. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Ma, C.-Y.; Gao, D.; Liu, X.-W.; Yuan, S.-S.; Ding, H. i2OM: Toward a Better Prediction of 2’-O-Methylation in Human RNA. Int. J. Biol. Macromol. 2023, 239, 124247. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.T.; Rakkiyapan, R.; Park, J.; Malik, A.; Manavalan, B. H2Opred: A Robust and Efficient Hybrid Deep Learning Model for Predicting 2’-O-Methylation Sites in Human RNA. Brief. Bioinform. 2023, 25, bbad476. [Google Scholar] [CrossRef]
- Tu, G.; Wang, X.; Xia, R.; Song, B. m6A-TCPred: A Web Server to Predict Tissue-Conserved Human m6A Sites Using Machine Learning Approach. BMC Bioinform. 2024, 25, 127. [Google Scholar] [CrossRef]
- Wang, H.; Huang, T.; Wang, D.; Zeng, W.; Sun, Y.; Zhang, L. MSCAN: Multi-Scale Self- and Cross-Attention Network for RNA Methylation Site Prediction. BMC Bioinform. 2024, 25, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, Y.; Liang, Y. TMSC-m7G: A Transformer Architecture Based on Multi-Sense-Scaled Embedding Features and Convolutional Neural Network to Identify RNA N7-Methylguanosine Sites. Comput. Struct. Biotechnol. J. 2024, 23, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jin, J.; Gao, W.; Qiao, J.; Wei, L. Moss-m7G: A Motif-Based Interpretable Deep Learning Method for RNA N7-Methlguanosine Site Prediction. J. Chem. Inf. Model. 2024. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).