
Polymorphism Identification in the Coding Sequences (ORFs) of the Porcine Pregnancy-Associated Glycoprotein 2-like Gene Subfamily in Pigs

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Genomic DNA (gDNA) Templates, and PCR Amplification
2.2. Sequencing and SNPs/InDels Identification in the ORF of the pPAG2-L
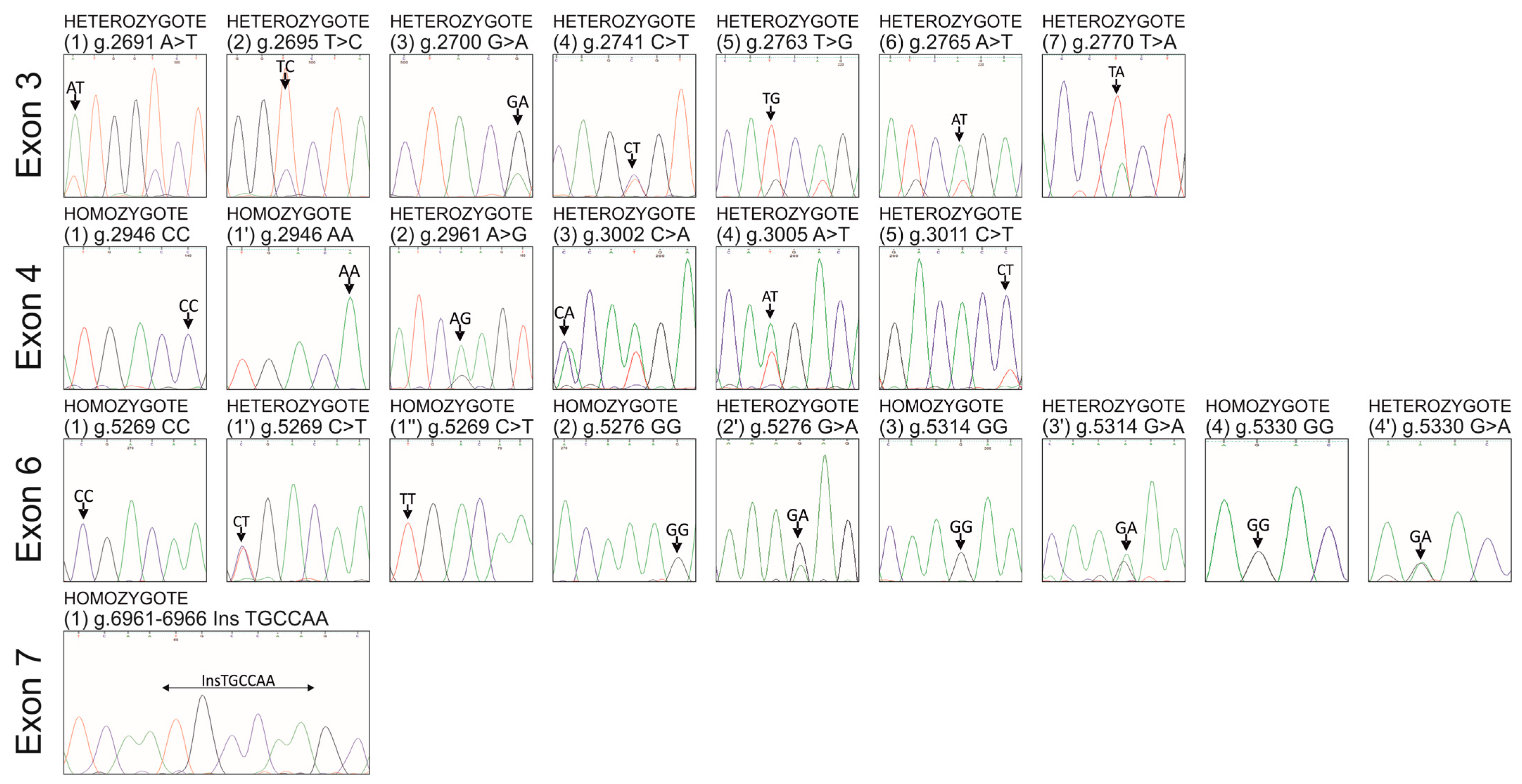
3. Results
3.1. Seven Novel SNPs in Exon 3 of the pPAG2-L in the Crossbreed Pigs
3.2. Five Novel SNPs in Exon 4 of the pPAG2-L in the Crossbreed Pigs
3.3. Four Novel SNPs in Exon 6 of the pPAG2-L in the Crossbreed Pigs
3.4. Novel InDel in Exon 7 of the pPAG2-L in the Crossbreed Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wallace, R.M.; Pohler, K.G.; Smith, M.F.; Green, J.A. Placental PAGs: Gene origins, expression patterns, and use as markers of pregnancy. Reproduction 2015, 149, R115–R126. [Google Scholar] [CrossRef]
- Hughes, A.L.; Green, J.A.; Garbayo, J.M.; Roberts, R.M. Adaptive diversification within a large family of recently duplicated, placentally expressed genes. Proc. Natl. Acad. Sci. USA 2000, 7, 3319–3323. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Green, J.A.; Piontkivska, H.; Roberts, R.M. Aspartic proteinase phylogeny and the origin of Pregnancy-Associated Glycoproteins. Mol. Biol. Evol. 2003, 20, 1940–1945. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Low, B.G.; Nagel, R.J.; Kramer, K.K.; Anthony, R.V.; Zoli, A.P.; Beckers, J.F.; Roberts, R.M. Identification of the major pregnancy specific antigens of cattle and sheep as inactive members of the aspartic proteinase family. Proc. Natl. Acad. Sci. USA 1991, 88, 10247–10251. [Google Scholar] [CrossRef]
- Xie, S.; Green, J.; Bixby, J.B.; Szafranska, B.; Demartini, J.C.; Hecht, S.; Roberts, R.M. The diversity and evolutionary relationships of the pregnancy-associated glycoproteins, an aspartic proteinase subfamily consisting of many trophoblast-expressed genes. Proc. Natl. Acad. Sci. USA 1997, 94, 12809–12816. [Google Scholar] [CrossRef] [PubMed]
- Szafranska, B.; Panasiewicz, G. The placental expression of the porcine pregnancy-associated glycoprotein (pPAG) gene family examined in situ and in vitro. Anim. Reprod. Sci. 2002, 72, 95–113. [Google Scholar] [CrossRef]
- Szafranska, B.; Xie, S.; Green, J.; Roberts, R.M. Porcine Pregnancy-Associated Glycoproteins: New members of the aspartic proteinase gene family expressed in the trophectoderm. Biol. Reprod. 1995, 53, 21–28. [Google Scholar] [CrossRef]
- Green, J.; Xie, S.; Szafranska, B.; Newman, A.; Gan, X.; McDowell, K.; Roberts, R.M. Identification of a new aspartic proteinase expressed by the outer chorionic cell layer of the equine placenta. Biol. Reprod. 1999, 60, 1069–1077. [Google Scholar] [CrossRef]
- Green, J.A.; Xie, S.; Quan, X.; Bao, B.; Gan, X.; Mathialagan, N.; Beckers, J.-F.; Roberts, R.M. Pregnancy-associated bovine and ovine glycoproteins exhibit spatially and temporally distinct expression patterns during pregnancy. Biol. Reprod. 2000, 62, 1624–1631. [Google Scholar] [CrossRef]
- Garbayo, J.M.; Green, J.; Manikkam, M.; Beckers, J.F.; Kiesling, D.O.; Ealy, A.D.; Roberts, R.M. Caprine Pregnancy-Associated Glycoproteins (PAGs): Their cloning, expression and evolutionary relationship to other PAG. Mol. Reprod. Dev. 2000, 57, 311–322. [Google Scholar] [CrossRef]
- Szafranska, B.; Panasiewicz, G.; Majewska, M.; Beckers, J.-F. Chorionic expression of heterogeneous products of the PAG (Pregnancy-Associated Glycoprotein) gene family secreted in vitro throughout embryonic and foetal development in the pig. Reprod. Nutr. Dev. 2003, 43, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Panasiewicz, G.; Majewska, M.; Szafranska, B. Trophoblastic cDNA cloning of porcine Pregnancy-Associated Glycoprotein genes (pPAG) and in silico analysis of coded polypeptide precursors. Reprod. Biol. 2004, 4, 131–141. [Google Scholar]
- Szafranska, B.; Majewska, M.; Panasiewicz, G. N-glycodiversity of the Pregnancy-Associated Glycoprotein family (PAG) produced in vitro by trophoblast and trophectoderm explants during implantation, placentation and advanced pregnancy in the pig. Reprod. Biol. 2004, 4, 67–89. [Google Scholar]
- Majewska, M.; Panasiewicz, G.; Dabrowski, M.; Gizejewski, Z.; Beckers, J.F.; Szafranska, B. Multiple forms of Pregnancy-Associated Glycoproteins released in vitro by porcine chorion or placentomal and interplacentomal explants of wild and domestic ruminants. Reprod. Biol. 2005, 5, 185–203. [Google Scholar] [PubMed]
- Majewska, M.; Panasiewicz, G.; Majewski, M.; Szafranska, B. Localization of chorionic Pregnancy-Associated Glycoprotein family in the pig. Reprod. Biol. 2006, 6, 205–230. [Google Scholar] [PubMed]
- Szafranska, B.; Panasiewicz, G.; Majewska, M. Biodiversity of multiple pregnancy-associated glycoprotein (PAG) family: Gene cloning and chorionic protein purification in domestic and wild eutherians (Placentalia)—A review. Reprod. Nutr. Dev. 2006, 5, 481–502. [Google Scholar] [CrossRef]
- Majewska, M.; Panasiewicz, G.; Louis, K.K.; Olivera, V.M.; Mamani, J.M.; Abd-Elnaeim, M.M.; Szafranska, B. Pregnancy-Associated Glycoprotein (PAG) family: Transcripts and gene amplicons in camelids. Reprod. Biol. 2009, 9, 127–150. [Google Scholar] [CrossRef]
- Telugu, B.P.; Walker, A.M.; Green, J.A. Characterization of the bovine pregnancy-associated glycoprotein gene family—Analysis of gene sequences, regulatory regions within the promoter and expression of selected genes. BMC Genom. 2009, 24, 185. [Google Scholar] [CrossRef]
- Majewska, M.; Panasiewicz, G.; Szafranska, B. Pregnancy-associated glycoprotein (PAG) family localized in chorionic cells within the epitheliochorial/diffuse placenta of the alpaca (Lama pacos). Acta Histochem. 2011, 113, 570–577. [Google Scholar] [CrossRef]
- Majewska, M.; Panasiewicz, G.; Szafranska, B. Expression of pregnancy-associated glycoprotein family in the epitheliochorial placenta of two Camelidae species (C. dromedarius and C. bactrianus). Acta Histochem. 2013, 115, 669–676. [Google Scholar] [CrossRef]
- Majewska, M.; Lipka, A.; Panasiewicz, G.; Gowkielewicz, M.; Jozwik, M.; Majewski, M.K.; Szafranska, B. Identification of Novel Placentally Expressed Aspartic Proteinase in Humans. Int. J. Mol. Sci. 2017, 8, 1227. [Google Scholar] [CrossRef]
- Lipka, A.; Panasiewicz, G.; Majewska, M.; Paukszto, L.; Bieniek-Kobuszewska, M.; Szafranska, B. Identification of Placental Aspartic Proteinase in the Eurasian Beaver (Castor fiber L.). Int. J. Mol. Sci. 2018, 19, 1229. [Google Scholar] [CrossRef]
- Chen, X.; Rosenfeld, C.S.; Roberts, R.M.; Green, J.A. An aspartic proteinase expressed in the yolk sac and neonatal stomach of the mouse. Biol. Reprod. 2001, 65, 1092–1101. [Google Scholar] [CrossRef]
- Vawter, G.A.; Parks, T.E.; Killian, G.; Ealy, A.D.; Green, J.A. Cloning and preliminary characterization of Pregnancy-Associated Glycoproteins expressed in the placenta of white-tailed deer. Biol. Reprod. 2004, 70, 153. [Google Scholar]
- Brandt, G.; Parks, T.E.; Killian, G.; Ealy, A.D.; Green, J.A. A cloning and expression analysis of pregnancy-associated glycoprotein expressed in throphoblast of the white-tail deer placenta. Molec. Reprod. Develop. 2007, 74, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.R. The structure and function of the aspartic proteinase. Annu. Rev. Biophys. Chem. 1990, 19, 189–215. [Google Scholar] [CrossRef]
- Szecsi, P.B. The aspartic proteinase. Scand. J. Clin. Lab. Investig. 1992, 52, 5–22. [Google Scholar] [CrossRef]
- Seelmeier, S.; Schmidt, H.; Turk, V.; von der Helm, K. Human immunodeficiency virus has an aspartic-type protease that can be inhibited by pepstatin A. Proc. Natl. Acad. Sci. USA 1988, 85, 6612–6616. [Google Scholar] [CrossRef]
- Kervinen, J.; Sarkkinen, P.; Kalkkinen, N.; Mikola, L.; Saarma, M. Hydrolytic specificity of the barley grain aspartic proteinase. Phytochemistry 1993, 32, 799–803. [Google Scholar] [CrossRef]
- Brouwer, E.E.; van Hellemond, J.J.; van Genderen, P.J.; Slot, E.; van Lieshout, L.; Visser, L.G.; Wismans, P.J. A case report of transfusion-transmitted Plasmodium malariae from an asymptomatic non-immune traveller. Malar J. 2013, 12, 439. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; Humphreys, M.J.; Matharu, P.; Granger, R.; Horrocks, P.; Moon, R.P.; Certa, U.; Ridley, R.G.; Bur, D.; Kay, J. A distinct member of the aspartic proteinase gene family from the human malaria parasite Plasmodium falciparum. FEBS Lett. 1999, 447, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Liu, J.; Beatty, W.; Pelosof, L.; Klemba, M.; Goldberg, D.E. Four plasmepsins are active in the Plasmodium falciparum. food vacuole, including a protease with an active-site histidine. Proc. Natl. Acad. Sci. USA 2002, 99, 990–995. [Google Scholar] [CrossRef]
- Xie, S.; Green, J.; Beckers, J.-F.; Roberts, R.M. The gene encoding bovine pregnancy-associated glycoprotein-1, an inactive member of the aspartic proteinase family. Gene 1995, 159, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Szafranska, B.; Miura, R.; Ghosh, D.; Ezashi, T.; Xie, S.; Roberts, R.M.; Green, J.A. Gene for porcine pregnancy-associated glycoprotein 2 (poPAG2): Its structural organization and analysis of its promoter. Mol. Reprod. Dev. 2001, 60, 137–146. [Google Scholar] [CrossRef]
- Guruprasad, K.; Blundell, T.L.; Xie, S.; Green, J.; Szafranska, B.; Nagel, R.J.; McDowell, K.; Baker, C.B.; Roberts, R.M. Comparative modelling and analysis of amino acid substitutions suggests that the family of pregnancy-associated glycoprotein includes both active and inactive aspartic proteinases. Prot. Eng. 1996, 9, 849–856. [Google Scholar] [CrossRef]
- Green, J.A.; Parks, T.E.; Avalle, M.P.; Telugu, B.P.; McLain, A.L.; Peterson, A.J.; McMillan, W.; Mathialagan, N.; Hook, R.R.; Xie, S.; et al. The establishment of an ELISA for the detection of pregnancy-associated glycoproteins (PAGs) in the serum of pregnant cows and heifers. Theriogenology 2005, 63, 1481–1503. [Google Scholar] [CrossRef]
- Whitlock, B.K.; Maxwell, H.S. Pregnancy-associated glycoproteins and pregnancy wastage in cattle. Theriogenology 2008, 70, 550–559. [Google Scholar] [CrossRef]
- Humblot, P. Use of pregnancy specific proteins and progesterone assays to monitor pregnancy and determine the timing, frequencies and sources of embryonic mortality in ruminants. Theriogenology 2001, 56, 1417–1433. [Google Scholar] [CrossRef]
- Jonker, F.H. Fetal death: Comparative aspects in large domestic animals. Anim. Reprod. Sci. 2004, 82–83, 415–430. [Google Scholar] [CrossRef]
- Friedrich, M.; Holtz, W. Establishment of an ELISA for measuring bovine pregnancy-associated glycoprotein in serum or milk and its application for early pregnancy detection. Reprod. Domest. Anim. 2010, 45, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Constant, F.; Camous, S.; Chavatte-Palmer, P.; Heyman, Y.; de Sousa, N.; Richardd, C.; Beckers, J.F.; Guillomot, M. Altered secretion of pregnancy-associated glycoproteins during gestation in bovine somatic clones. Theriogenology 2011, 76, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Takahaschi, T.; Hayashi, K.-G.; Hosoe, M. Biology of the placental proteins in domestic ruminants: Expression, proposed roles and practical applications. JARQ 2013, 47, 43–51. [Google Scholar] [CrossRef]
- Ropstad, E.; Veiberg, V.; Säkkinen, H.; Dahl, E.; Kinahl, H.; Holand, Ø.; Beckers, J.F.; Eloranta, E. Endocrinology of pregnancy and early pregnancy detection by reproductive hormones in reindeer (Rangifer tarandus tarandus). Theriogenology 2005, 63, 1775–1788. [Google Scholar] [CrossRef] [PubMed]
- Savela, H.; Vahtiala, S.; Lindeberg, H.; Dahl, E.; Ropstad, E.; Beckers, J.F.; Saarela, S. Comparison of accuracy of ultrasonography, progesterone, and pregnancy-associated glycoprotein tests for pregnancy diagnosis in semidomesticated reindeer. Theriogenology 2009, 72, 1229–1236. [Google Scholar] [CrossRef]
- El-Battawy, K.A.; Sousa, N.M.; Szenci, O.; Beckers, J.F. Pregnancy-associated glycoprotein profile during the first trimester of pregnancy in Egyptian buffalo cows. Reprod. Domest. Anim. 2009, 44, 161–166. [Google Scholar] [CrossRef]
- Abdulkareem, T.A.; Al-Sharifi, S.; Ishak, M.A.; Eidan, S.M.; Alnimr, M.A.; Passavant, C.W.; Branen, J.R.; Sasser, R.G. Early pregnancy detection of iraqi riverine buffalo (Bubalus bubalis) using the BioPRYN enzyme-linked immunosorbent assay for PSPB and the progesterone assay. Reprod. Domest. Anim. 2011, 46, 455–462. [Google Scholar] [CrossRef]
- Gabor, G.; Toth, F.; Ozsvari, L.; Abonyi-Toth, Z.; Sasser, R.G. Early detection of pregnancy and embryonic loss in dairy cattle by ELISA tests. Reprod. Domest. Anim. 2007, 42, 633–636. [Google Scholar] [CrossRef]
- Lopes-da-Costa, L.; Chagas e Silvaa, J.; Deloche, M.C.; Jeanguyot, N.; Humblot, P.; Horta, A.E. Effects of embryo size at transfer (whole versus demi) and early pregnancy progesterone supplementation on embryo growth and pregnancy-specific protein bovine concentrations in recipient dairy heifers. Theriogenology 2011, 76, 522–531. [Google Scholar] [CrossRef]
- García-Ispierto, I.; Almería, S.; Serrano, B.; de Sousa, N.M.; Beckers, J.F.; López-Gatius, F. Plasma concentrations of pregnancy-associated glycoproteins measured using anti-bovine PAG-2 antibodies on day 120 of gestation predict abortion in dairy cows naturally infected with Neospora caninum. Reprod. Domest. Anim. 2013, 48, 613–618. [Google Scholar] [CrossRef]
- Breukelman, S.P.; Perényi, Z.; Taverne, M.A.M.; Jonker, H.; van der Weijden, G.C.; Vos, P.L.; de Ruigh, L.; Dieleman, S.J.; Beckers, J.F.; Szenci, O. Characterisation of pregnancy losses after embryo transfer by measuring plasma progesterone and bovine pregnancy-associated glycoprotein-1concentrations. Vet. J. 2012, 194, 71–76. [Google Scholar] [CrossRef]
- Bieniek-Kobuszewska, M.; Panasiewicz, G.; Lipka, A.; Majewska, M.; Szafranska, B. Novel SNPs and InDels discovered in two promoter regions of porcine pregnancy-associated glycoprotein 2-like subfamily (pPAG2-Ls) in crossbreed pigs. Funct. Integr. Genom. 2016, 6, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Bieniek-Kobuszewska, M.; Panasiewicz, G.; Szafranska, B. Identification of 1093-bp intron A, SNPs and InDels discovered in the porcine pregnancy-associated glycoprotein 2-like (pPAG2-L) gene subfamily in pigs. Genetika 2020, 52, 851–866. [Google Scholar] [CrossRef]
- Panasiewicz, G.; Bieniek-Kobuszewska, M.; Lipka, A.; Majewska, M.; Jedryczko, R.; Szafranska, B. Novel effects of identified SNPs within porcine Pregnancy-Associated Glycoprotein gene family (pPAGs) on the major reproductive traits in Hirschmann hybrid line sows. Res. Vet. Sci. 2017, 114, 123–130. [Google Scholar] [CrossRef]
- Bidanel, J.P. Mapping of quantitative trait loci for reproductive traits in pigs. In Control of Pig Reproduction IX; Rodriguez-Martinez, H., Soede, N.M., Flowers, W.L., Eds.; Context Products, Ltd.: Leicestershire, UK, 2013; pp. 287–300. [Google Scholar]
- McLaren, D.G.; Cleveland, M.A.; Deeb, N.; Selma, F.; Mileham, A.J.; Newman, S.; Southwood, O.I.; Wang, L. Genome-wide selection for reproductive traits in swine. In Control of Pig Reproduction IX; Rodriguez-Martinez, H., Soede, N.M., Flowers, W.L., Eds.; Context Products, Ltd.: Leicestershire, UK, 2013; pp. 227–242. [Google Scholar]
- Hu, Z.L.; Park, C.A.; Reecy, J.M. Bringing the Animal QTLdb and CorrDB into the future: Meeting new challenges and providing updated services. Nucleic Acids Res. 2022, 50, D956–D961. [Google Scholar] [CrossRef]
- Aleksandrow, L.B.; Serena, N.-Z.; Wedge, D.C. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef]
- Myers, S.; Freeman, C.; Auton, A.; Donnely, P.; McVean, G. A common sequence motif associated with recombination hot spots and genome instability in humans. Nat. Genet. 2008, 40, 1124–1129. [Google Scholar] [CrossRef]
- Amos, W. Even small SNP clusters are non-randomly distributed: Is this evidence of mutational non-independence? Proc. Biol. Sci. 2010, 277, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Tenaillon, M.I.; Austerlitz, F.; Tenaillon, O. Apparent mutational hotspots and long distance linkage disequilibrium resulting from a bottleneck. J. Evol. Biol. 2008, 21, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Lohmueller, K.E.; Indap, A.R.; Schmidt, S.; Boyko, A.R.; Hernandez, R.D.; Hubisz, M.J.; Sninsky, J.J.; White, T.J.; Sunyaev, S.R.; Nielsen, R.; et al. Proportionally more deleterious genetic variation in European than in African populations. Nature 2008, 451, 994–997. [Google Scholar] [CrossRef]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Fang, M.D.; Huang, T.H.; Li, C.C.; Yu, M.; Zhao, S.H. Detection of differentially expressed genes between Erhualian and Large White placentas on day 75 and 90 of gestation. BMC Genom. 2009, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Franczak, A.; Wojciechowicz, B.; Kotwica, G. Transcriptomic analysis of the porcine endometrium during early pregnancy and the estrous cycle. Reprod. Biol. 2013, 13, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, K. Analysis of uteroplacental-specific molecules and their functions during implantation and placentation in the bovine. J. Reprod. Dev. 2007, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Exons Studied | Primer | Sequence |
|---|---|---|
| Ex1 | F: pr500se | 5′GACTGCCTGTGTCTTACT3′ |
| R: IntrAas | 5′GACTGTCAGGAATGATGGCA3′ | |
| Ex1 | F: sensATG | 5′TGGASCCAGGAAAGAAGCATG3′ |
| R: IntrAasSr | 5′GGATGAGAGATGGAGCCAGGAG3′ | |
| Ex2 | Ex2se | 5′TACAACATGATCCAGAATCT3′ |
| R: Ex3asSR | 5′GATGAGSCGGTGTCAAAKAYGAC3′ | |
| Ex3 | F: IntrBsedo3 | 5′GCTGCTCCAATGAGGCACTGGT3′ |
| R: Ex4as | 5′GTCATGGAAGGTGCTGGAGTG3′ | |
| Ex3,4 | F: Ex3seP | 5′TGGTCTACGTGGGCAACATCA3′ |
| R: IntrDasSr | 5′CCTAGAGTAAGAATGCACTTAGCAG3′ | |
| Ex4 | F: Ex4pr | 5′CCATCTACTGCAAAAGCAAG3′ |
| R: Ex5as | 5′GGGATAGGCCAGGCCCAGGA3′ | |
| Ex5 | F: IntrDsedo5 | 5′AGG AGCGGCCGTGGGTTGAAAT3′ |
| R: IntrEas | 5′CTTCCTTATCGAATCCGTCAGCG3′ | |
| Ex6 | F: IntrEsedo6 | 5′GACTCTCTCAGTAGGCTGATTGC3′ |
| R: Ex8as | 5′GACRTTGTTAATGGTGAAGA3′ | |
| Ex6 | F: IntrEsedo6 | 5′GACTCTCTCAGTAGGCTGATTGC3′ |
| R: IntrFasGap | 5′TAGCCTGGAGCAAAGTGGTAATTCATTC3′ | |
| Ex7,8,9 | F: IntrFsedo7 | 5′GAATGAATTACCACTTTGCTCCAGGCTA3′ |
| R: Ex8as | 5′GACRTTGTTAATGGTGAAGA3′ | |
| F: nEx7se | 5′GCTGCCAGGCCATCKTGGATA3′ | |
| R: pagC1155 | 5′CAGGCCAATCCTGTTCTGTCCT3′ |
| SNP Locus (IUPAC Code) * | SNP Locus acc. DNASIS ** | Genotype Frequencies (px) | |||||
|---|---|---|---|---|---|---|---|
| Genotype | px | Genotype | px | Genotype | px | ||
| Exon 3 | |||||||
| g.2691A>T (W) | g.1A>T | AA | 0 | AT | 1.0 | TT | 0 |
| g.2695T>C (Y) | g.5T>C | TT | 0 | TC | 1.0 | CC | 0 |
| g.2700G>A (R) | g.10G>A | GG | 0 | GA | 1.0 | AA | 0 |
| g.2741C>T (Y) | g.51C>T | CC | 0 | CT | 1.0 | TT | 0 |
| g.2763T>G (K) | g.73T>G | TT | 0 | TG | 1.0 | GG | 0 |
| g.2765A>T (W) | g.75A>T | AA | 0 | AT | 1.0 | TT | 0 |
| g.2770T>A (W) | g.80T>A | TT | 0 | TA | 1.0 | AA | 0 |
| Exon 4 | |||||||
| g.2946C>A (M) | g.48C>A | CC | 0.33 | AC | 0 | AA | 0.67 |
| g.2961A>G (R) | g.63A>G | AA | 0.21 | AG | 0.79 | GG | 0 |
| g.3002C>A (M) | g.104C>A | CC | 0 | AC | 1.0 | AA | 0 |
| g.3005A>T (W) | g.107A>T | AA | 0.07 | AT | 0.93 | TT | 0 |
| g.3011C>T (Y) | g.113C>T | CC | 0.07 | CT | 0.93 | TT | 0 |
| Exon 6 | |||||||
| g.5269C>T (Y) | g.1C>T | CC | 0.08 | CT | 0.63 | TT | 0.29 |
| g.5276G>A (R) | g.8G>A | GG | 0.64 | AG | 0.36 | AA | 0 |
| g.5314G>A (R) | g.46G>A | GG | 0.85 | AG | 0.15 | AA | 0 |
| g.5330G>A (R) | g.62G>A | GG | 0.9 | AG | 0.1 | AA | 0 |
| Exon 7 | |||||||
| g.6961–6966 Ins TGCCAA | g.121–126 Ins TGCCAA | TGCCAA | 1.0 | - | - | - | - |
| SNP Locus (IUPAC Code) * | pPAG2 Codon | Consensus/Code [aa] | Codon with SNP | Localization of Substitution [aa] ** | Substitution/Code [aa] |
|---|---|---|---|---|---|
| Exon 3 | |||||
| g.2691A>T (W) | ATG | Met (M) | TTG | 74 | Leu (L) |
| g.2695T>C (Y) | GTC | Val (V) | GCC | 75 | Ala (A) |
| g.2700G>A (R) | GTG | Val (V) | ATG | 77 | Met (M) |
| g.2741C>T (Y) | AGC | Ser (S) | AGT | 90 | Ser (S) |
| g.2763T>G (K) | TCA | Ser (S) | GCT | 98 | Ala (A) |
| g.2765A>T (W) | TCA | Ser (S) | GCT | 98 | Ala (A) |
| g.2770T>A (W) | CTC | Leu (L) | CAC | 100 | His (H) |
| Exon 4 | |||||
| g.2946C>A (M) | CGG | Arg (R) | AGG | 129 | Arg (R) |
| g.2961A>G (R) | AAG | Lys (K) | GAG | 134 | Glu (E) |
| g.3002C>A (M) | GGC | Gly (G) | GGA | 147 | Gly (G) |
| g.3005A>T (W) | CAA | Gln (Q) | CAT | 148 | His (H) |
| g.3011C>T (Y) | ACC | Thr (T) | ACT | 150 | Thr (T) |
| Exon 6 | |||||
| g.5269C>T (Y) | CGA | Ser (S) | TGA | 219 | Ser (S) |
| g.5276G>A (R) | GAG | Glu (E) | AAG | 222 | Lys (K) |
| g.5314G>A (R) | AAG | Lys (K) | AAA | 235 | Lys (K) |
| g.5330G>A (R) | GAC | Asp (D) | AAC | 240 | Asn (N) |
| Exon 7 | |||||
| g.6961–6966 Ins TGCCAA | - | - | TGCCAA | 298–299 | Asn (N) Ala (A) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bieniek-Kobuszewska, M.; Panasiewicz, G. Polymorphism Identification in the Coding Sequences (ORFs) of the Porcine Pregnancy-Associated Glycoprotein 2-like Gene Subfamily in Pigs. Genes 2024, 15, 1149. https://doi.org/10.3390/genes15091149
Bieniek-Kobuszewska M, Panasiewicz G. Polymorphism Identification in the Coding Sequences (ORFs) of the Porcine Pregnancy-Associated Glycoprotein 2-like Gene Subfamily in Pigs. Genes. 2024; 15(9):1149. https://doi.org/10.3390/genes15091149
Chicago/Turabian StyleBieniek-Kobuszewska, Martyna, and Grzegorz Panasiewicz. 2024. "Polymorphism Identification in the Coding Sequences (ORFs) of the Porcine Pregnancy-Associated Glycoprotein 2-like Gene Subfamily in Pigs" Genes 15, no. 9: 1149. https://doi.org/10.3390/genes15091149
APA StyleBieniek-Kobuszewska, M., & Panasiewicz, G. (2024). Polymorphism Identification in the Coding Sequences (ORFs) of the Porcine Pregnancy-Associated Glycoprotein 2-like Gene Subfamily in Pigs. Genes, 15(9), 1149. https://doi.org/10.3390/genes15091149