
Molecular Mechanism of HINTW (HINT Gene in W-Chromosome)-Mediated UBE2I (Ubiquitin Conjugating Enzyme E2 I) Interaction in Female Differentiation of Chicken Embryos

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Expression and Detection of His-HINTW Protein
2.3. Protein Purification and Concentration
2.4. His Pull-Down Assay
2.5. Mass Spectrometry (MS) Data Acquisition and Analysis
2.6. Co-Immunoprecipitation (Co-IP)
3. Results
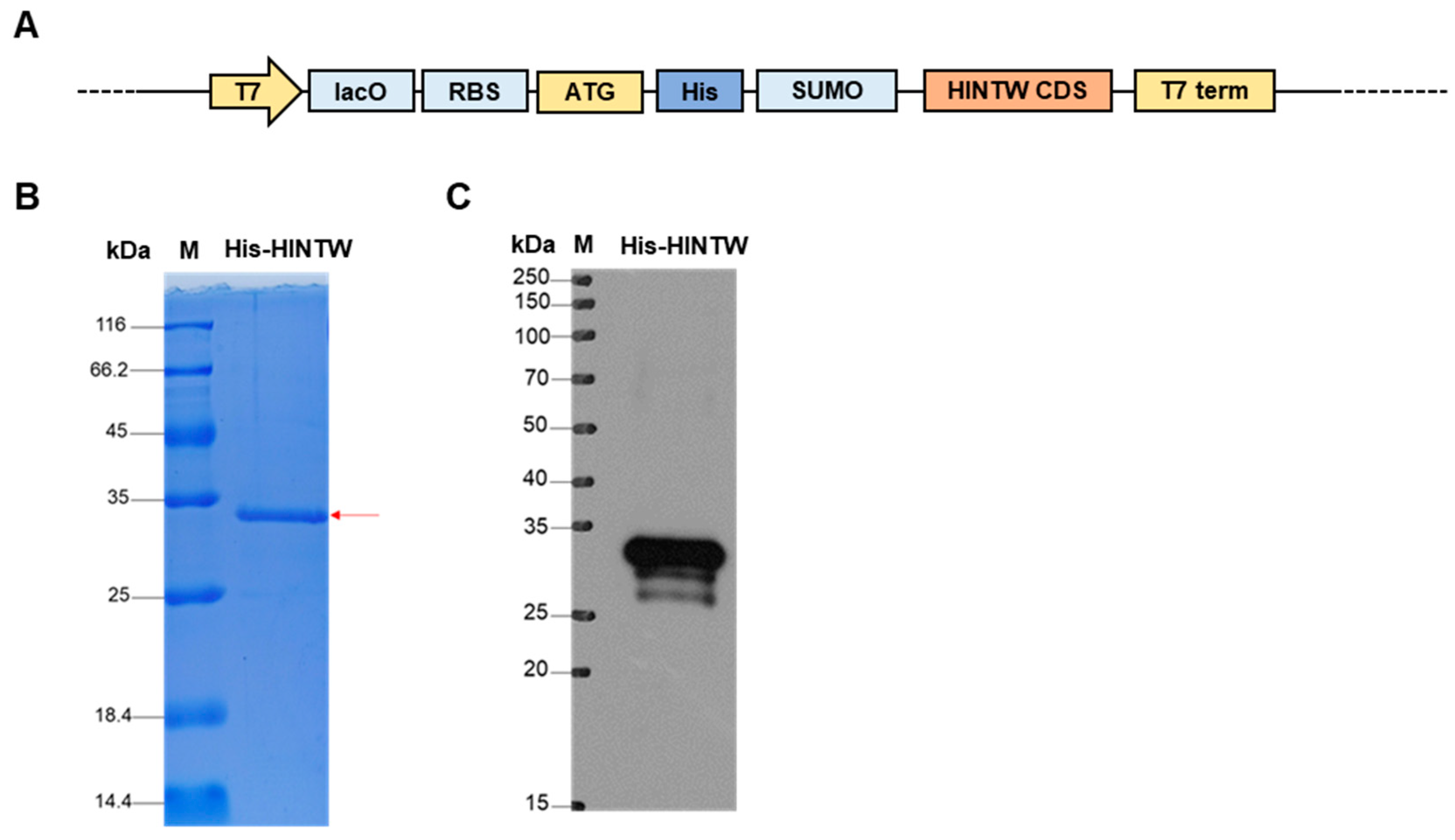
3.1. The Construction and Identification of the pET-SUMO-HINTW
3.2. The Identification of the HINTW Interaction Protein via Pull-Down
3.3. The Analysis of HINTW Interacting Proteins
3.4. Indirect Immunofluorescence Staining for PGCLCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hori, T.; Asakawa, S.; Itoh, Y.; Shimizu, N.; Mizuno, S. Wpkci, encoding an altered form ofPKCI, is conserved widely on the avian W chromosome and expressed in early female embryos: Implication of its role in female sex determination. Mol. Biol. Cell 2000, 11, 3645–3660. [Google Scholar] [CrossRef] [PubMed]
- O’neill, M.; Binder, M.; Smith, C.; Andrews, J.; Reed, K.; Smith, M.; Millar, C.; Lambert, D.; Sinclair, A. ASW: A gene with conserved avian W-linkage and female specific expression in chick embryonic gonad. Dev. Genes Evol. 2000, 210, 243–249. [Google Scholar] [CrossRef]
- Ceplitis, H.; Ellegren, H. Adaptive molecular evolution of HINTW, a female-specific gene in birds. Mol. Biol. Evol. 2003, 21, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Backström, N.; Ceplitis, H.; Berlin, S.; Ellegren, H. Gene conversion drives the evolution of HINTW, an ampliconic gene on the female-specific avian W chromosome. Mol. Biol. Evol. 2005, 22, 1992–1999. [Google Scholar] [CrossRef]
- Nagai, H.; Sezaki, M.; Bertocchini, F.; Fukuda, K.; Sheng, G. HINTW, a W-chromosome HINT gene in chick, is expressed ubiquitously and is a robust female cell marker applicable in intraspecific chimera studies. Genesis 2014, 52, 424–430. [Google Scholar] [CrossRef]
- Hirst, C.E.; Major, A.T.; Smith, C.A. Sex determination and gonadal sex differentiation in the chicken model. Int. J. Dev. Biol. 2018, 62, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Roeszler, K.N.; Sinclair, A.H. Genetic evidence against a role for W-linked histidine triad nucleotide binding protein (HINTW) in avian sex determination. Int. J. Dev. Biol. 2009, 53, 59–67. [Google Scholar] [CrossRef]
- Bakovic, V.; Höglund, A.; Cerezo, M.L.M.; Henriksen, R.; Wright, D. Genomic and gene expression associations to morphology of a sexual ornament in the chicken. G3 2022, 12, jkac174. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Chen, S.; Yang, N.; Zheng, J. Overview of Avian Sex Reversal. Int. J. Mol. Sci. 2023, 24, 8284. [Google Scholar] [CrossRef]
- Ayers, K.L.; Davidson, N.M.; Demiyah, D.; Roeszler, K.N.; Grützner, F.; Sinclair, A.H.; Oshlack, A.; Smith, C.A. RNA sequencing reveals sexually dimorphic gene expression before gonadal differentiation in chicken and allows comprehensive annotation of the W-chromosome. Genome Biol. 2013, 14, R26. [Google Scholar] [CrossRef]
- Hirst, C.E.; Major, A.T.; Ayers, K.L.; Brown, R.J.; Mariette, M.; Sackton, T.B.; A Smith, C.; Sackton, T. Sex reversal and comparative data undermine the W chromosome and support Z-linked DMRT1 as the regulator of gonadal sex differentiation in birds. Endocrinology 2017, 158, 2970–2987. [Google Scholar] [CrossRef] [PubMed]
- Ayers, K.L.; Smith, C.A.; Lambeth, L.S. The molecular genetics of avian sex determination and its manipulation. Genesis 2013, 51, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhou, Q. The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized. Genes 2020, 11, 1126. [Google Scholar] [CrossRef]
- Yazdi, H.P.; Ellegren, H. A Genetic Map of Ostrich Z Chromosome and the Role of Inversions in Avian Sex Chromosome Evolution. Genome Biol. Evol. 2018, 10, 2049–2060. [Google Scholar] [CrossRef]
- Jin, K.; Li, D.; Jin, J.; Song, J.; Zhang, Y.; Chang, G.; Chen, G.; Li, B. C1EIP functions as an activator of ENO1 to promote chicken PGCs formation via inhibition of the notch signaling pathway. Front. Genet. 2020, 11, 751. [Google Scholar] [CrossRef]
- Sun, C.; Jin, K.; Zhou, J.; Zuo, Q.; Song, J.; Yani, Z.; Chen, G.; Li, B. Role and function of the Hintw in early sex differentiation in chicken (Gallus gallus) embryo. Anim. Biotechnol. 2021, 34, 56–66. [Google Scholar] [CrossRef]
- Jin, K.; Zhou, J.; Zuo, Q.-S.; Li, J.-C.; Jiuzhou, S.; Zhang, Y.-N.; Chang, G.-B.; Chen, G.-H.; Li, B.-C. UBE2I stimulates female gonadal differentiation in chicken (Gallus gallus) embryos. J. Integr. Agric. 2021, 20, 2986–2994. [Google Scholar] [CrossRef]
- Liu, J.A.J.; Wu, M.-H.; Yan, C.H.; Chau, B.K.H.; So, H.; Ng, A.; Chan, A.; Cheah, K.S.E.; Briscoe, J.; Cheung, M. Phosphorylation of Sox9 is required for neural crest delamination and is regulated downstream of BMP and canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 2882–2887. [Google Scholar] [CrossRef]
- Guo, Y.; Cheng, L.; Li, X.; Tang, S.; Zhang, X.; Gong, Y. Transcriptional regulation of CYP19A1 expression in chickens: ESR1, ESR2 and NR5A2 form a functional network. Gen. Comp. Endocrinol. 2022, 315, 113939. [Google Scholar] [CrossRef]
- Jin, K.; Zuo, Q.; Song, J.; Zhang, Y.; Chen, G.; Li, B. CYP19A1 (aromatase) dominates female gonadal differentiation in chicken (Gallus gallus) embryos sexual differentiation. Biosci. Rep. 2020, 40, BSR20201576. [Google Scholar] [CrossRef]
- Wang, J.; Gong, Y. Transcription of CYP19A1 is directly regulated by SF-1 in the theca cells of ovary follicles in chicken. Gen. Comp. Endocrinol. 2017, 247, 1–7. [Google Scholar] [CrossRef]
- Ishikawaa, T.; Nakada-Moriya, Y.; Andoc, C.; Tandac, N.; Nishidac, S.; Minatogawaa, Y.; Nohnond, T. Expression of the JNK2-α1 gene in the developing chick brain. Biochem. Biophys. Res. Commun. 1997, 234, 489–492. [Google Scholar] [CrossRef]
- Hajihoseini, Z.; Eghbalsaied, S. Simultaneous effects of IGF1 and Fadrozole on parthenogenesis and pluripotency markers in chicken embryo. Theriogenology 2018, 114, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Jung, K.M.; Rengaraj, D.; Lee, K.Y.; Yoo, E.; Kim, T.H.; Han, J.Y. Single-cell RNA sequencing of mitotic-arrested prospermatogonia with DAZL::GFP chickens and revealing unique epigenetic reprogramming of chickens. J. Anim. Sci. Biotechnol. 2022, 13, 64. [Google Scholar] [CrossRef]
- Luo, X.; Guo, J.; Zhang, J.; Ma, Z.; Li, H. Overview of chicken embryo genes related to sex differentiation. PeerJ 2024, 12, e17072. [Google Scholar] [CrossRef]
- Hayashi, T.; Seki, M.; Maeda, D.; Wang, W.; Kawabe, Y.-I.; Seki, T.; Saitoh, H.; Fukagawa, T.; Yagi, H.; Enomoto, T. Ubc9 is essential for viability of higher eukaryotic cells. Exp. Cell Res. 2002, 280, 212–221. [Google Scholar] [CrossRef]
- Cox, A.R.; Chernis, N.; Kim, K.H.; Masschelin, P.M.; Saha, P.K.; Briley, S.M.; Sharp, R.; Li, X.; Felix, J.B.; Sun, Z.; et al. Ube2i deletion in adipocytes causes lipoatrophy in mice. Mol. Metab. 2021, 48, 101221. [Google Scholar] [CrossRef]
- Yang, H.; Gao, S.; Chen, J.; Lou, W. UBE2I promotes metastasis and correlates with poor prognosis in hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 234. [Google Scholar] [CrossRef]
- Ihara, M.; Stein, P.; Schultz, R.M. UBE2I (UBC9), a SUMO-conjugating enzyme, localizes to nuclear speckles and stimulates transcription in mouse oocytes. Biol. Reprod. 2008, 79, 906–913. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Song, J.; Ali, M.A.; Sun, H.; Niu, Y.; Zuo, Q.; Han, W.; Li, B.; Jin, K. Molecular Mechanism of HINTW (HINT Gene in W-Chromosome)-Mediated UBE2I (Ubiquitin Conjugating Enzyme E2 I) Interaction in Female Differentiation of Chicken Embryos. Genes 2025, 16, 366. https://doi.org/10.3390/genes16040366
Sun C, Song J, Ali MA, Sun H, Niu Y, Zuo Q, Han W, Li B, Jin K. Molecular Mechanism of HINTW (HINT Gene in W-Chromosome)-Mediated UBE2I (Ubiquitin Conjugating Enzyme E2 I) Interaction in Female Differentiation of Chicken Embryos. Genes. 2025; 16(4):366. https://doi.org/10.3390/genes16040366
Chicago/Turabian StyleSun, Changhua, Jiuzhou Song, Malik Ahsan Ali, Hongyan Sun, Yingjie Niu, Qisheng Zuo, Wei Han, Bichun Li, and Kai Jin. 2025. "Molecular Mechanism of HINTW (HINT Gene in W-Chromosome)-Mediated UBE2I (Ubiquitin Conjugating Enzyme E2 I) Interaction in Female Differentiation of Chicken Embryos" Genes 16, no. 4: 366. https://doi.org/10.3390/genes16040366
APA StyleSun, C., Song, J., Ali, M. A., Sun, H., Niu, Y., Zuo, Q., Han, W., Li, B., & Jin, K. (2025). Molecular Mechanism of HINTW (HINT Gene in W-Chromosome)-Mediated UBE2I (Ubiquitin Conjugating Enzyme E2 I) Interaction in Female Differentiation of Chicken Embryos. Genes, 16(4), 366. https://doi.org/10.3390/genes16040366