Intercompartmental Piecewise Gene Transfer

Abstract
1. Introduction
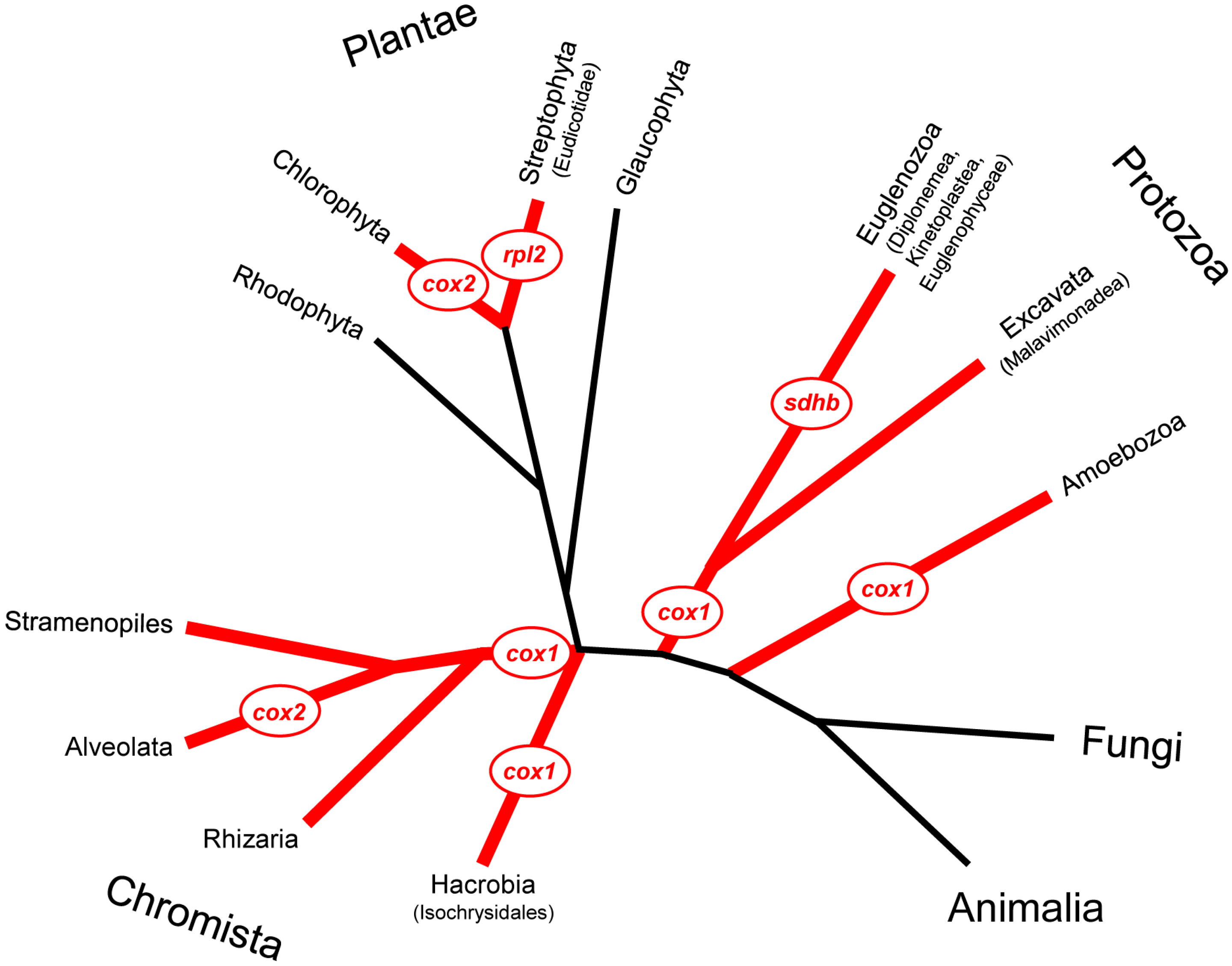
2. Cases of Piecewise Mitochondrial Gene Relocation to the Nucleus
2.1. The Cox2 Gene
2.1.1. Endosymbiotic Transfer of Cox2 in Chlorophyceae (Chlorophyta) and Alveolata
2.1.2. Mitochondrial Cox2 Fission in Wasps Campsomeris (Arthropoda: Insecta)
2.2. Transfer to the Nucleus of rpl2 in Eudicots (Tracheophyta: Angiospermae)
2.3. Nuclear Relocation of the 3’ End-Fragment of cox1 in the Majority of Protists
2.4. Transfer of sdhB in Euglenozoa
3. Fragmented RNA Genes
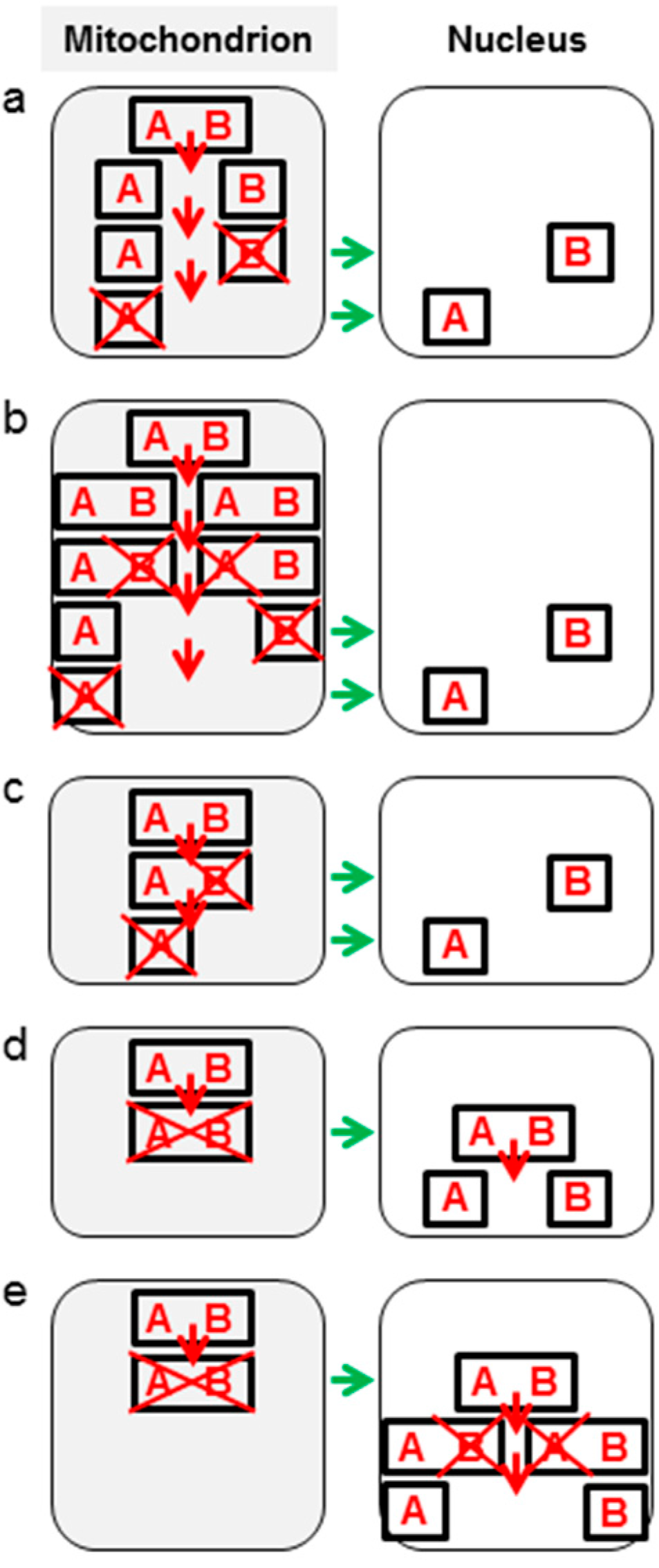
4. Mechanisms of Piecewise Intercompartmental Gene Transfer
4.1. General Models
4.2. Transfer and Integration of Nucleic Acids
5. The Hydrophobicity-Importability Hypothesis
6. Partial Gene Conversion
6.1. Chimeric Mitochondrial Genes
6.2. Mitochondrial-to-Nuclear Gene Conversion
7. Concluding Remarks
Conflicts of Interest
References
- Adams, K.L.; Palmer, J.D. Evolution of mitochondrial gene content: gene loss and transfer to the nucleus. Mol. Phylogenet. Evol. 2003, 29, 380–395. [Google Scholar] [CrossRef]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W. Endosymbiotic gene transfer: organellar genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Origin, evolution and genetic effects of nuclear insertions of organellar DNA. Trends Genet. 2005, 21, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Kleine, T.; Maier, U.G.; Leister, D. DNA transfer from organellars to the nucleus: The idiosyncratic genetics of endosymbiosis. Annu. Rev. Plant. Biol. 2009, 60, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Witnessing genome evolution: Experimental reconstruction of endosymbiotic and horizontal gene transfer. Annu. Rev. Genet. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.; Nelson-Sathi, S.; Roettger, M.; Sousa, F.L.; Lockhart, P.J.; Bryant, D.; Hazkani-Covo, E.; McInerney, J.O.; Landan, G.; Martin, W.F. Endosymbiotic origin and differential loss of eukaryotic genes. Nature 2015, 524, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Harish, A.; Kurland, C.G. Mitochondria are not captive bacteria. J. Theor. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bock, R.; Timmis, J.N. Reconstructing evolution: Gene transfer from plastids to the nucleus. Bioessays 2008, 30, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Bensasson, D.; Zhang, D.; Hartl, D.L.; Hewitt, G.M. Mitochondrial pseudogenes: Evolution’s misplaced witnesses. Trends Ecol. Evol. 2001, 16, 314–321. [Google Scholar] [CrossRef]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W. Molecular poltergeists: Mitochondrial DNA copies (NUMTs) in sequenced nuclear genomes. PLoS Genet. 2010, 6, e1000834. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.S. Intracellular mitochondrial DNA transfers to the nucleus in human cancer cells. Curr. Opin. Genet. Dev. 2016, 38, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Noutsos, C.; Kleine, T.; Armbruster, U.; DalCorso, G.; Leister, D. Nuclear insertions of organellar DNA can create novel patches of functional exon sequences. Trends Genet. 2007, 23, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. The relation of recombination to mutational advance. Mutat. Res. 1964, 1, 2–9. [Google Scholar] [CrossRef]
- Lynch, M. Mutation accumulation in transfer RNAs: Molecular evidence for Muller’s rachet in mitochondrial genomes. Mol. Biol. Evol. 1996, 13, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Berg, O.G.; Kurland, C.G. Why mitochondrial genes are most often found in nuclei. Mol. Biol. Evol. 2000, 17, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F.; Raven, J.A. Free-radical-induced mutation vs redox regulation: Costs and benefits of genes in organellars. J. Mol. Evol. 1996, 42, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.; Albert, B.; Godelle, B. Reducing the genome size of organellars favours gene transfer to the nucleus. Trends Ecol. Evol. 2001, 16, 135–141. [Google Scholar] [CrossRef]
- Gemmell, N.J.; Braischer, T.L. Organellar genome evolution. Trends Ecol. Evol. 2001, 16, 489–490. [Google Scholar] [CrossRef]
- Gray, M.W.; Schnare, M.N. Evolution of rRNA gene organization. In Ribosomal RNA: Structure, Evolution, Processing, and Function in Protein Biosynthesis; Zimmermann, R., Dahlberg, A., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 49–69. [Google Scholar]
- Evguenieva-Hackenberg, E. Bacterial ribosomal RNA in pieces. Mol. Microbiol. 2005, 57, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Kanai, A. Disrupted tRNA Genes and tRNA fragments: A perspective on tRNA gene evolution. Life 2015, 5, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Shiba, K.; Schimmel, P. Functional assembly of a randomly cleaved protein. Proc. Natl. Acad. Sci. USA 1992, 89, 1880–1884. [Google Scholar] [CrossRef] [PubMed]
- Magliery, T.J.; Wilson, C.G.; Pan, W.; Mishler, D.; Ghosh, I.; Hamilton, A.D.; Regan, L. Detecting protein-protein interactions with a green fluorescent protein fragment reassembly trap: scope and mechanism. J. Am. Chem. Soc. 2005, 127, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Fukutani, Y.; Ishii, J.; Kondo, A.; Ozawa, T.; Matsunami, H.; Yohda, M. Split luciferase complementation assay for the analysis of G protein-coupled receptor ligand response in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2017, 114, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Dolan, M.J.; Luan, H.; Shropshire, W.C.; Sutcliffe, B.; Cocanougher, B.; Scott, R.L.; Frechter, S.; Zlatic, M.; Jefferis, G.S.X.E.; White, B.H. Facilitating neuron-specific genetic manipulations in Drosophila melanogaster using a split GAL4 repressor. Genetics 2017, 206, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Ishibashi, K.; Toki, S. A split Staphylococcus aureus Cas9 as a compact genome-editing tool in plants. Plant. Cell Physiol. 2017, 58, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Snel, B.; Bork, P.; Huynen, M. Genome evolution. Gene fusion versus gene fission. Trends Genet. 2000, 16, 9–11. [Google Scholar] [CrossRef]
- Kummerfeld, S.K.; Teichmann, S.A. Relative rates of gene fusion and fission in multi-domain proteins. Trends Genet. 2005, 21, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Marianayagam, N.J.; Sunde, M.; Matthews, J.M. The power of two: Protein dimerization in biology. Trends Biochem. Sci. 2004, 29, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Nishi, H.; Bryant, S.; Panchenko, A.R. Caught in self-interaction: evolutionary and functional mechanisms of protein homooligomerization. Phys. Biol. 2011, 8, 035007. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M. The evolution of multimeric protein assemblages. Mol. Biol. Evol. 2012, 29, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Kück, U.; Jekosch, K.; Holzamer, P. DNA sequence analysis of the complete mitochondrial genome of the green alga Scenedesmus obliquus: Evidence for UAG being a leucine and UCA being a non-sense codon. Gene 2000, 253, 13–18. [Google Scholar] [CrossRef]
- Nedelcu, A.M.; Lee, R.W.; Lemieux, C.; Gray, M.W.; Burger, G. The complete mitochondrial DNA sequence of Scenedesmus obliquus reflects an intermediate stage in the evolution of the green algal mitochondrial genome. Genome Res. 2000, 10, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, X.; Antaramian, A.; Vazquez-Acevedo, M.; Funes, S.; Tolkunova, E.; d’Alayer, J.; Claros, M.G.; Davidson, E.; King, M.P.; González-Halphen, D. Subunit II of cytochrome c oxidase in Chlamydomonad algae is a heterodimer encoded by two independent nuclear genes. J. Biol. Chem. 2001, 276, 11302–11309. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, M.A.; Gordon, D.P.; Orrell, T.M.; Bailly, N.; Bourgoin, T.; Brusca, R.C.; Cavalier-Smith, T.; Guiry, M.D.; Kirk, P.M. A higher level classification of all living organisms. PLoS ONE 2015, 10, e0119248. [Google Scholar]
- Burger, G.; Zhu, Y.; Littlejohn, T.G.; Greenwood, S.J.; Schnare, M.N.; Lang, B.F.; Gray, M.W. Complete sequence of the mitochondrial genome of Tetrahymena pyriformis and comparison with Paramecium aurelia mitochondrial DNA. J. Mol. Biol. 2000, 297, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Edqvist, J.; Burger, G.; Gray, M.W. Expression of mitochondrial protein-coding genes in Tetrahymena pyriformis. J. Mol. Biol. 2000, 297, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Swart, E.C.; Nowacki, M.; Shum, J.; Stiles, H.; Higgins, B.P.; Doak, T.G.; Schotanus, K.; Magrini, V.J.; Minx, P.; Mardis, E.R.; et al. The Oxytricha trifallax mitochondrial genome. Genome Biol. Evol. 2012, 4, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Handa, H.; Bonnard, G.; Grienenberger, J.M. The rapeseed mitochondrial gene encoding a homologue of the bacterial protein Ccl1 is divided into two independently transcribed reading frames. Mol. Gen. Genet. 1996, 252, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Unseld, M.; Marienfeld, J.R.; Brandt, P.; Brennicke, A. The mitochondrial genome of Arabidopsis thaliana contains 57 genes in 366,924 nucleotides. Nat. Genet. 1997, 15, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Rayapuram, N.; Hagenmuller, J.; Grienenberger, J.M.; Bonnard, G.; Giegé, P. The three mitochondrial encoded CcmF proteins form a complex that interacts with CCMH and c-type apocytochromes in Arabidopsis. J. Biol. Chem. 2008, 283, 25200–25208. [Google Scholar] [CrossRef] [PubMed]
- Szafranski, P. Evolutionarily recent, insertional fission of mitochondrial cox2 into complementary genes in bilaterian Metazoa. BMC Genom. 2017, 18, 269. [Google Scholar] [CrossRef] [PubMed]
- Funes, S.; Davidson, E.; Reyes-Prieto, A.; Magallón, S.; Herion, P.; King, M.P.; González-Halphen, D. A green algal apicoplast ancestor. Science 2002, 298, 2155. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Salinas, E.; Riveros-Rosas, H.; Li, Z.; Fucíková, K.; Brand, J.J.; Lewis, L.A.; González-Halphen, D. Lineage-specific fragmentation and nuclear relocation of the mitochondrial cox2 gene in chlorophycean green algae (Chlorophyta). Mol. Phylogenet. Evol. 2012, 64, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.F.; Keeling, P.J.; van Dooren, G.G.; McFadden, G.I. Comment on ´A green algal apicoplast ancestor´. Science 2003, 301, 49. [Google Scholar] [CrossRef] [PubMed]
- Waller, R.F.; Keeling, P.J. Alveolate and chlorophycean mitochondrial cox2 genes split twice independently. Gene 2006, 383, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Ong, H.C.; Palmer, J.D. Mitochondrial gene transfer in pieces: Fission of the ribosomal protein gene rpl2 and partial or complete gene transfer to the nucleus. Mol. Biol. Evol. 2001, 18, 2289–2297. [Google Scholar] [CrossRef] [PubMed]
- Gawryluk, R.M.; Gray, M.W. An ancient fission of mitochondrial cox1. Mol. Biol. Evol. 2010, 27, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Gawryluk, R.M.; Gray, M.W. A split and rearranged nuclear gene encoding the iron-sulfur subunit of mitochondrial succinate dehydrogenase in Euglenozoa. BMC Res. Notes 2009, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [Google Scholar] [CrossRef] [PubMed]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Prochnik, S.E.; Umen, J.; Nedelcu, A.M.; Hallmann, A.; Miller, S.M.; Nishii, I.; Ferris, P.; Kuo, A.; Mitros, T.; Fritz-Laylin, L.K.; et al. Genomic analysis of organismal complexity in the multicellular green alga Volvox carteri. Science. 2010, 329, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Forget, L.; Ustinova, J.; Wang, Z.; Huss, V.A.; Lang, B.F. Hyaloraphidium curvatum: A linear mitochondrial genome, tRNA editing, and an evolutionary link to lower fungi. Mol. Biol. Evol. 2002, 19, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, T.Y.; Schnare, M.N.; Young, P.G.; Gray, M.W. Rearranged coding segments, separated by a transfer RNA gene, specify the two parts of a discontinuous large subunit ribosomal RNA in Tetrahymena pyriformis mitochondria. J. Biol. Chem. 1987, 262, 2879–2887. [Google Scholar] [PubMed]
- Boer, P.H.; Gray, M.W. Scrambled ribosomal RNA gene pieces in Chlamydomonas reinhardtii mitochondrial DNA. Cell 1988, 55, 399–411. [Google Scholar] [CrossRef]
- Denovan-Wright, E.M.; Lee, R.W. Comparative structure and genomic organization of the discontinuous mitochondrial ribosomal RNA genes of Chlamydomonas eugametos and Chlamydomonas reinhardtii. J. Mol. Biol. 1994, 241, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Denovan-Wright, E.M.; Sankoff, D.; Spencer, D.F.; Lee, R.W. Evolution of fragmented mitochondrial ribosomal RNA genes in Chlamydomonas. J. Mol. Evol. 1996, 42, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Kairo, A.; Fairlamb, A.H.; Gobright, E.; Nene, V. A 7.1 kb linear DNA molecule of Theileria parva has scrambled rDNA sequences and open reading frames for mitochondrially encoded proteins. EMBO J. 1994, 13, 898–905. [Google Scholar] [PubMed]
- Feagin, J.E.; Gardner, M.J.; Williamson, D.H.; Wilson, R.J. The putative mitochondrial genome of Plasmodium falciparum. J. Protozool. 1991, 38, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Slamovits, C.H.; Saldarriaga, J.F.; Larocque, A.; Keeling, P.J. The highly reduced and fragmented mitochondrial genome of the early-branching dinoflagellate Oxyrrhis marina shares characteristics with both apicomplexan and dinoflagellate mitochondrial genomes. J. Mol. Biol. 2007, 372, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Kamikawa, R.; Inagaki, Y.; Sako, Y. Fragmentation of mitochondrial large subunit rRNA in the dinoflagellate Alexandrium catenella and the evolution of rRNA structure in alveolate mitochondria. Protist 2007, 158, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Dellaporta, S.L.; Xu, A.; Sagasser, S.; Jakob, W.; Moreno, M.A.; Buss, L.W.; Schierwater, B. Mitochondrial genome of Trichoplax adhaerens supports placozoa as the basal lower metazoan phylum. Proc. Natl. Acad. Sci. USA 2006, 103, 8751–8756. [Google Scholar] [CrossRef] [PubMed]
- Signorovitch, A.Y.; Buss, L.W.; Dellaporta, S.L. Comparative genomics of large mitochondria in placozoans. PLoS Genet. 2007, 3, e13. [Google Scholar] [CrossRef] [PubMed]
- Milbury, C.A.; Gaffney, P.M. Complete mitochondrial DNA sequence of the eastern oyster Crassostrea virginica. Mar. Biotechnol. 2005, 7, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wei, Z.; Kong, X.; Shi, W. Complete mitochondrial DNA sequence of oyster Crassostrea hongkongensis—A case of ‘Tandem duplication-random loss’ for genome rearrangement in Crassostrea? BMC Genom. 2008, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Liu, X.; Zhang, G.; Liu, B.; Guo, X. ´Tandem duplication-random loss´ is not a real feature of oyster mitochondrial genomes. BMC Genom. 2009, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Milbury, C.A.; Lee, J.C.; Cannone, J.J.; Gaffney, P.M.; Gutell, R.R. Fragmentation of the large subunit ribosomal RNA gene in oyster mitochondrial genomes. BMC Genom. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.J.; Norman, J.E.; Schnare, M.N.; Gray, M.W.; Keeling, P.J.; Waller, R.F. Broad genomic and transcriptional analysis reveals a highly derived genome in dinoflagellate mitochondria. BMC Biol. 2007, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Randau, L.; Münch, R.; Hohn, M.J.; Jahn, D.; Söll, D. Nanoarchaeum equitans creates functional tRNAs from separate genes for their 5’- and 3’-halves. Nature 2005, 433, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Fujishima, K.; Sugahara, J.; Tomita, M.; Kanai, A. Sequence evidence in the archaeal genomes that tRNAs emerged through the combination of ancestral genes as 5’ and 3’ tRNA halves. PLoS ONE 2008, 3, e1622. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Cozen, A.E.; Lowe, T.M. Discovery of permuted and recently split transfer RNAs in Archaea. Genome Biol. 2011, 12, R38. [Google Scholar] [CrossRef] [PubMed]
- Masta, S.E.; Boore, J.L. Parallel evolution of truncated transfer RNA genes in arachnid mitochondrial genomes. Mol. Biol. Evol. 2008, 25, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Leroux, M.; Jani, N.; Sandler, S.J. A priA mutant expressed in two pieces has almost full activity in Escherichia coli K-12. J. Bacteriol. 2017, 199, e00267-17. [Google Scholar] [CrossRef] [PubMed]
- Nugent, J.M.; Palmer, J.D. RNA-mediated transfer of the gene coxII from the mitochondrion to the nucleus during flowering plant evolution. Cell 1991, 66, 473–481. [Google Scholar] [CrossRef]
- Adams, K.L.; Daley, D.O.; Qiu, Y.L.; Whelan, J.; Palmer, J.D. Repeated, recent and diverse transfers of a mitochondrial gene to the nucleus in flowering plants. Nature 2000, 408, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sloan, D.B.; Brown, C.W.; Rosenblueth, M.; Palmer, J.D.; Ong, H.C. Mitochondrial retroprocessing promoted functional transfers of rpl5 to the nucleus in grasses. Mol. Biol. Evol. 2017, 34, 2340–2354. [Google Scholar] [CrossRef] [PubMed]
- Thorsness, P.E.; White, K.H.; Fox, T.D. Inactivation of YME1, a member of the ftsH-SEC18-PAS1-CDC48 family of putative ATPase-encoding genes, causes increased escape of DNA from mitochondria in Saccharomyces cerevisiae. Mol. Cell. Biol. 1993, 13, 5418–5426. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, H.; Jagendorf, A.T. DNA strand-transfer activity in pea (Pisum sativum L.) chloroplasts. Plant Physiol. 1993, 102, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Thorsness, P.E. Escape of mitochondrial DNA to the nucleus in yme1 yeast is mediated by vacuolar-dependent turnover of abnormal mitochondrial compartments. J. Cell Sci. 1998, 111, 2455–2464. [Google Scholar] [PubMed]
- Thorsness, P.E.; Fox, T.D. Escape of DNA from mitochondria to the nucleus in Saccharomyces cerevisiae. Nature 1990, 346, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Thorsness, P.E.; Weber, E.R. Escape and migration of nucleic acids between chloroplasts, mitochondria, and the nucleus. Int. Rev. Cytol. 1996, 165, 207–234. [Google Scholar] [PubMed]
- Brandes, D.; Schofield, B.H.; Anton, E. Nuclear mitochondria? Science 1965, 149, 1373–1374. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Russell, S.D. Occurrence of mitochondria in the nuclei of tobacco sperm cells. Plant Cell 1994, 6, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Takemura, G.; Takatsu, Y.; Sakaguchi, H.; Fujiwara, H. Intranuclear mitochondria in human myocardial cells. Pathol. Res. Pract. 1997, 193, 305–311. [Google Scholar] [CrossRef]
- Bakeeva, L.E.; Skulachev, V.P.; Sudarikova, Y.V.; Tsyplenkova, V.G. Mitochondria enter the nucleus (one further problem in chronic alcoholism). Biochemistry 2001, 66, 1335–1341. [Google Scholar] [PubMed]
- Bilewitch, J.P.; Degnan, S.M. A unique horizontal gene transfer event has provided the octocoral mitochondrial genome with an active mismatch repair gene that has potential for an unusual self-contained function. BMC Evol. Biol. 2011, 11, 228. [Google Scholar] [CrossRef] [PubMed]
- Ricchetti, M.; Tekaia, F.; Dujon, B. Continued colonization of the human genome by mitochondrial DNA. PLoS Biol. 2004, 2, e273. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.H.; Timmis, J.N. The origin and characterization of new nuclear genes originating from a cytoplasmic organellar genome. Mol. Biol. Evol. 2011, 28, 2019–2028. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Timmis, J.N. Cytoplasmic organellar DNA preferentially inserts into open chromatin. Genome Biol. Evol. 2013, 5, 1060–1064. [Google Scholar] [CrossRef] [PubMed]
- Von Heijne, G. Why mitochondria need a genome. FEBS Lett. 1986, 198, 1–4. [Google Scholar] [CrossRef]
- Popot, J.L.; de Vitry, C. On the microassembly of integral membrane proteins. Annu. Rev. Biophys. Biophys. Chem. 1990, 19, 369–403. [Google Scholar] [CrossRef] [PubMed]
- Claros, M.G.; Perea, J.; Shu, Y.; Samatey, F.A.; Popot, J.L.; Jacq, C. Limitations to in vivo import of hydrophobic proteins into yeast mitochondria. The case of a cytoplasmically synthesized apoapocytochrome b. Eur. J. Biochem. 1995, 228, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.O.; Whelan, J. Why genes persist in organelle genomes. Genome Biol. 2005, 6, 110. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Johnston, I.G.; Williams, B.P. Evolutionary inference across eukaryotes identifies specific pressures favoring mitochondrial gene retention. Cell Syst. 2016, 2, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Björkholm, P.; Ernst, A.M.; Hagström, E.; Andersson, S.G. Why mitochondria need a genome revisited. FEBS Lett. 2017, 591, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Song, K.; Roessler, P.G.; Nugent, J.M.; Doyle, J.L.; Doyle, J.J.; Palmer, J.D. Intracellular gene transfer in action: Dual transcription and multiple silencings of nuclear and mitochondrial cox2 genes in legumes. Proc. Natl. Acad. Sci. USA 1999, 96, 13863–13868. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.O.; Clifton, R.; Whelan, J. Intracellular gene transfer: Reduced hydrophobicity facilitates gene transfer for subunit 2 of cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2002, 99, 10510–10515. [Google Scholar] [CrossRef] [PubMed]
- Oca-Cossio, J.; Kenyon, L.; Hao, H.; Moraes, C.T. Limitations of allotopic expression of mitochondrial genes in mammalian cells. Genetics 2003, 165, 707–720. [Google Scholar] [PubMed]
- Supekova, L.; Supek, F.; Greer, J.E.; Schultz, P.G. A single mutation in the first transmembrane domain of yeast COX2 enables its allotopic expression. Proc. Natl. Acad. Sci. USA 2010, 107, 5047–5052. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Why chloroplasts and mitochondria retain their own genomes and genetic systems: Colocation for redox regulation of gene expression. Proc. Natl. Acad. Sci. USA 2015, 112, 10231–10238. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Richardson, A.O.; Zheng, Y.; Palmer, J.D. Gorgeous mosaic of mitochondrial genes created by horizontal transfer and gene conversion. Proc. Natl. Acad. Sci. USA 2010, 107, 21576–21581. [Google Scholar] [CrossRef] [PubMed]
- Bergthorsson, U.; Adams, K.L.; Thomason, B.; Palmer, J.D. Widespread horizontal transfer of mitochondrial genes in flowering plants. Nature 2003, 424, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Khakhlova, O.; Bock, R. Elimination of deleterious mutations in plastid genomes by gene conversion. Plant J. 2006, 46, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Barkman, T.J.; McNeal, J.R.; Lim, S.H.; Coat, G.; Croom, H.B.; Young, N.D.; Depamphilis, C.W. Mitochondrial DNA suggests at least 11 origins of parasitism in angiosperms and reveals genomic chimerism in parasitic plants. BMC Evol. Biol. 2007, 7, 248. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Palmer, J.D. Fine-scale mergers of chloroplast and mitochondrial genes create functional, transcompartmentally chimeric mitochondrial genes. Proc. Natl. Acad. Sci. USA 2009, 106, 16728–16733. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, P.; León, G.; Gómez, I.; Carmona, R.; Figueroa, P.; Holuigue, L.; Araya, A.; Jordana, X. Transfer of RPS14 and RPL5 from the mitochondrion to the nucleus in grasses. Gene 2004, 324, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Liu, Z.; Adams, K.L. Evolutionary transfers of mitochondrial genes to the nucleus in the Populus lineage and coexpression of nuclear and mitochondrial Sdh4 genes. New Phytol. 2006, 172, 429–439. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Heterodimeric or Trimeric Protein | Gene | Taxonomy | Reference | |
|---|---|---|---|---|
| Mitochondrial | Nuclear | |||
| NAD1 | nad1a, nad1b | - | Paramecium and other ciliates (Chromista: Alveolata) | [37,38,39] |
| NAD2 | nad2a, nad2b | - | ||
| RPS3 | rps3a, rps3b | - | ||
| CCMF | ccmFN, ccmFC1, ccmFC2 | - | liverwort Marchantia (Plantae: Marchantiophyta) | [40,41,42] |
| ccmFN (=ccb577), ccmFC | - | Triticum and most of the land plants (Plantae: Tracheophyta: Angiospermae) | ||
| ccmFN1, ccmFN2, ccmFC (=ccb452) | - | Brassica, Arabidopsis (Plantae: Tracheophyta: Angiospermae: Brassicaceae) | ||
| COXII | cox2a, cox2b | - | Campsomeris (Animalia: Arthropoda: Insecta) | [43] |
| cox2a | cox2b | Scenedesmus and other Sphaeropleales, Chaetophorales (Plantae: Chlorophyta: Chlorophyceae) | ||
| - | cox2a, cox2b | Chlamydomonas and other Chlamydomonadales (Plantae: Chlorophyta: Chlorophyceae); Plasmodium and other apicomplexan parasites, Karlodinium and other dinoflagellates, Perkinsus (Chromista: Alveolata) | [33,34,35,44,45,46,47,48] | |
| RPL2 | 5’rpl2 | 3’rpl2 | tomato, Arabidopsis, cotton (Plantae: Angiospermae: Eudicotidae) | [1,49] |
| - | 5’rpl2, 3’rpl2 | legumes, lettuce (Plantae: Angiospermae: Eudicotidae) | ||
| COXI | cox1(-) | cox1-c (=Dictiostelium coxIV) | Trypanosoma, Euglena (Protozoa: Euglenozoa); Malawimonas (Protozoa: Loucozoa); Acanthamoeba, Dictiostelium, Physarum (Protozoa: Amoebozoa); Phytophtora, Thalassiosira (Chromista: Stramenopiles); Plasmodium, Tetrahymena (Chromista: Alveolata); Bigielowiella (Chromista: Rhizaria); Emiliania (Chromista: Hacrobia) | [50] |
| SDHB | - | sdhb-n, sdhB-c | Diplonema, Trypanosoma, Euglena (Protozoa: Euglenozoa) | [51] |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szafranski, P. Intercompartmental Piecewise Gene Transfer. Genes 2017, 8, 260. https://doi.org/10.3390/genes8100260
Szafranski P. Intercompartmental Piecewise Gene Transfer. Genes. 2017; 8(10):260. https://doi.org/10.3390/genes8100260
Chicago/Turabian StyleSzafranski, Przemyslaw. 2017. "Intercompartmental Piecewise Gene Transfer" Genes 8, no. 10: 260. https://doi.org/10.3390/genes8100260
APA StyleSzafranski, P. (2017). Intercompartmental Piecewise Gene Transfer. Genes, 8(10), 260. https://doi.org/10.3390/genes8100260