Genome and Epigenome Surveillance Processes Underlying UV Exposure in Plants


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Ultraviolet-Induced DNA Damage
3. Ultraviolet-Induced Genomic Changes
4. Ultraviolet-Induced Epigenome Dynamics
5. Repair of UV-Induced DNA Damage
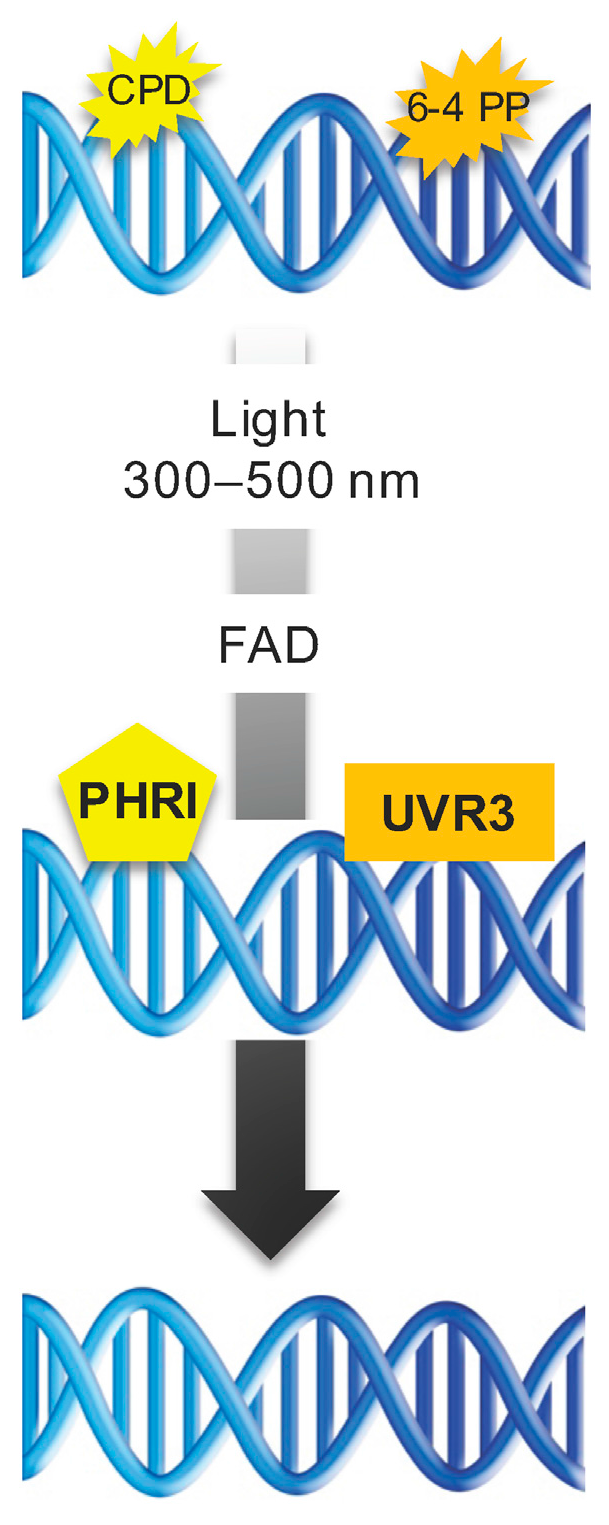
5.1. Direct Repair
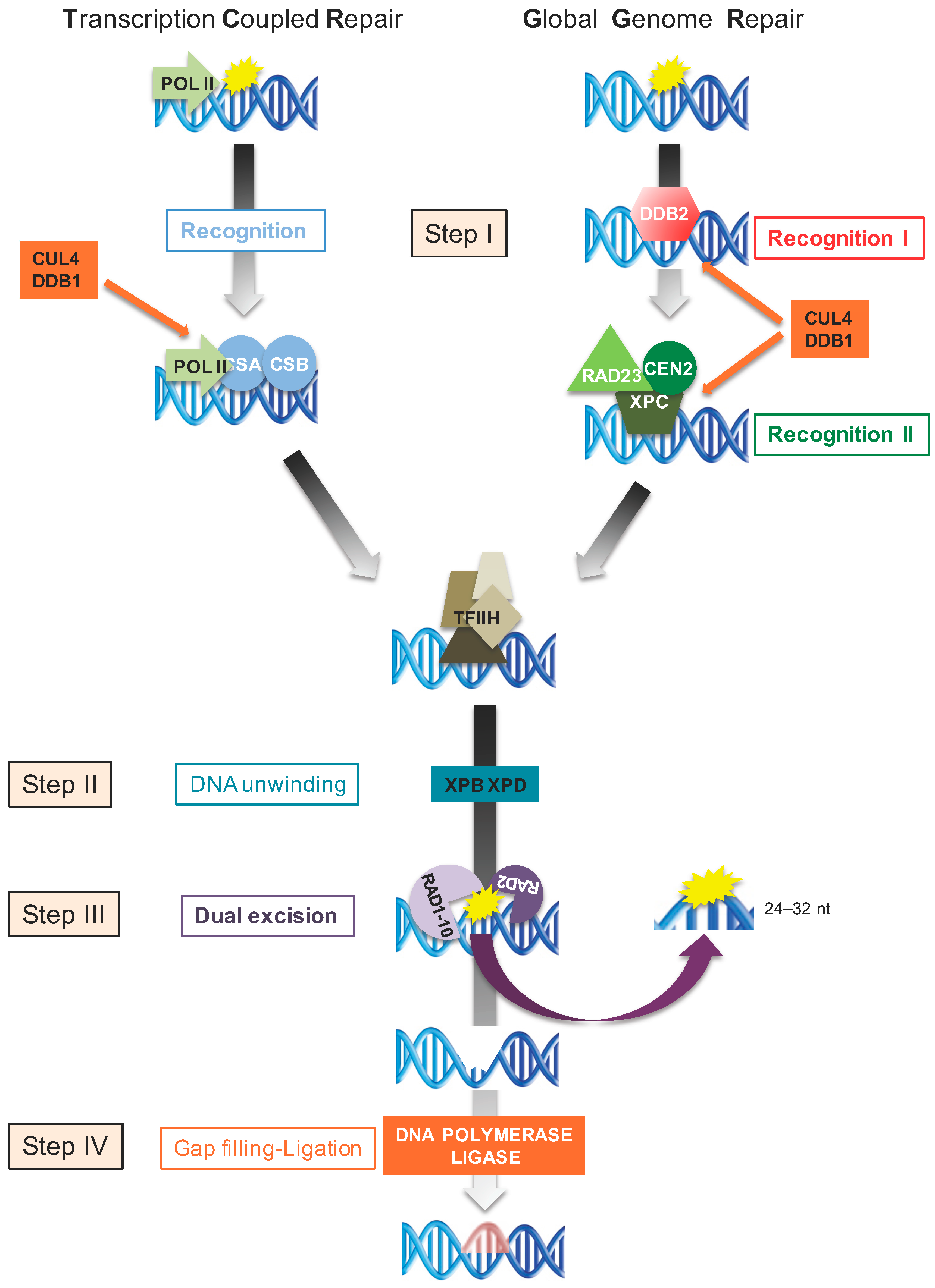
5.2. Nucleotide Excision Repair
5.3. Transcription-Coupled Repair
5.4. Global Genome Repair
5.5. Dual Incision, Repair Synthesis and Ligation
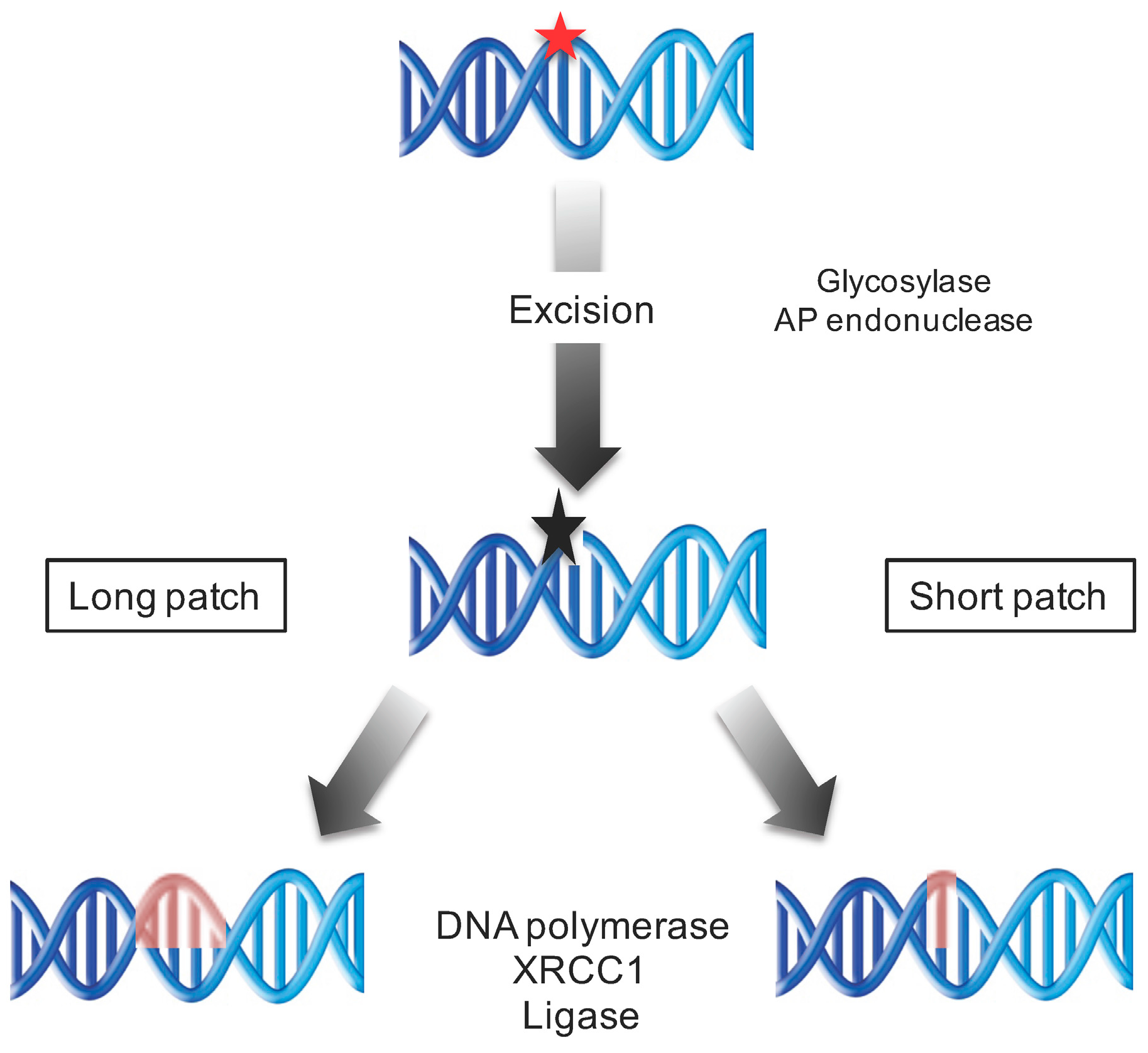
5.6. Repair of Oxidatively Induced DNA Damage by the Base Excision Repair
6. Double-Strand Break Repair
7. Interplay between Genome and Epigenome Maintenance Processes
8. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef] [PubMed]
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 5, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Schuch, A.P.; Garcia, C.C.; Makita, K.; Menck, C.F. DNA damage as a biological sensor for environmental sunlight. Photochem. Photobiol. Sci. 2013, 8, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Bourbousse, C.; Mestiri, I.; Zabulon, G.; Bourge, M.; Formiggini, F.; Koini, M.A.; Brown, S.C.; Fransz, P.; Bowler, C.; Barneche, F. Light signaling controls nuclear architecture reorganization during seedling establishment. Proc. Natl. Acad. Sci. USA 2015, 21, E2836–E2844. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Ulm, R. How plants cope with UV-B: From perception to response. Curr. Opin. Plant Biol. 2017, 37, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Rozema, J.; van de Staaij, J.; Björn, L.O.; Caldwell, M. UV-B as an environmental factor in plant life: Stress and regulation. Trends Ecol. Evol. 1997, 12, 22–28. [Google Scholar] [CrossRef]
- Shepherd, T.; Wynne Griffiths, D. The effects of stress on plant cuticular waxes. New Phytol. 2006, 3, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Mazza, C.A.; Boccalandro, H.E.; Giordano, C.V.; Battista, D.; Scopel, A.L.; Ballare, C.L. Functional significance and induction by solar radiation of UV-absorbing sunscreens in field-grown soybean crops. Plant Physiol. 2000, 122, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Hidema, J.; Tamura, K.; Takagi, S.; Hara-Nishimura, I. Plant nuclei move to escape ultraviolet-induced DNA damage and cell death. Plant Physiol. 2016, 2, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Britt, A.B. Repair of DNA damage induced by ultraviolet radiation. Plant Physiol. 1995, 108, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Schuermann, D.; Molinier, J.; Fritsch, O.; Hohn, B. The dual nature of homologous recombination in plants. Trends Genet. 2005, 3, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Sakaguchi, K. DNA repair in plants. Chem. Rev. 2006, 2, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Tadele, Z.; Hofmann, I.; Probst, A.V.; Angelis, K.J.; Kaya, H.; Araki, T.; Mengiste, T.; Mittelsten Scheid, O.; Shibahara, K.; et al. BRU1, a novel link between responses to DNA damage and epigenetic gene silencing in Arabidopsis. Genes Dev. 2004, 18, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Qüesta, J.I.; Fina, J.P.; Casati, P. DDM1 and ROS1 have a role in UV-B induced- and oxidative DNA damage in A. thaliana. Front. Plant Sci. 2013, 4, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donà, M.; Mittelsten Scheid, O. DNA damage repair in the context of plant chromatin. Plant Physiol. 2015, 4, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.L.; Dunn, J.; Rees, A.; Rünger, T.M. No formation of DNA double-strand breaks and no activation of recombination repair with UVA. J. Investig. Dermatol. 2011, 5, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.W.; Kamiyama, C.; Hidema, J.; Hikosaka, K. Ultraviolet-B-induced DNA damage and ultraviolet-B tolerance mechanisms in species with different functional groups coexisting in subalpine moorlands. Oecologia 2016, 4, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Mouret, S.; Charveron, M.; Favier, A.; Cadet, J.; Douki, T. Differential repair of UVB-induced cyclobutane pyrimidine dimers in cultured human skin cells and whole human skin. DNA Repair 2008, 5, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Adebali, O.; Adar, S.; Sancar, A. Dynamic maps of UV damage formation and repair for the human genome. Proc. Natl. Acad. Sci. USA 2017, 26, 6758–6763. [Google Scholar] [CrossRef] [PubMed]
- Schalk, C.; Cognat, V.; Graindorge, S.; Vincent, T.; Voinnet, O.; Molinier, J. Small RNA-mediated repair of UV-induced DNA lesions by the DNA damage-binding protein 2 and argonaute 1. Proc. Natl. Acad. Sci. USA 2017, 14, E2965–E2974. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, G.P.; You, Y.H.; Besaratinia, A. Mutations induced by ultraviolet light. Mutat. Res. 2005, 1–2, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Willing, E.M.; Piofczyk, T.; Albert, A.; Winkler, J.B.; Schneeberger, K.; Pecinka, A. UVR2 ensures transgenerational genome stability under simulated natural UV-B in Arabidopsis thaliana. Nat. Commun. 2016, 7, 13522. [Google Scholar] [CrossRef] [PubMed]
- Cunniffe, S.; Walker, A.; Stabler, R.; O’Neill, P.; Lomax, M.E. Increased mutability and decreased repairability of a three-lesion clustered DNA-damaged site comprised of an AP site and bi-stranded 8-oxoG lesions. Int. J. Radiat. Biol. 2014, 6, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Swoboda, P.; Gal, S.; Hohn, B.; Puchta, H. Intrachromosomal homologous recombination in whole plants. EMBO J. 1994, 2, 484–489. [Google Scholar]
- Ries, G.; Buchholz, G.; Frohnmeyer, H.; Hohn, B. UV-damage-mediated induction of homologous recombination in Arabidopsis is dependent on photosynthetically active radiation. Proc. Natl. Acad. Sci. USA 2000, 24, 13425–13429. [Google Scholar] [CrossRef] [PubMed]
- Ries, G.; Heller, W.; Puchta, H.; Sandermann, H.; Seidlitz, H.K.; Hohn, B. Elevated UV-B radiation reduces genome stability in plants. Nature 2000, 6791, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Molinier, J.; Ramos, C.; Fritsch, O.; Hohn, B. CENTRIN 2 modulates homologous recombination and nucleotide excision repair in Arabidopsis. Plant Cell 2004, 16, 1633–1643. [Google Scholar] [CrossRef] [PubMed]
- Walbot, V. UV-B damage amplified by transposons in maize. Nature 1999, 6718, 398–399. [Google Scholar] [CrossRef]
- Questa, J.; Walbot, V.; Casati, P. UV-B radiation induces Mu element somatic transposition in maize. Mol. Plant. 2013, 6, 2004–2007. [Google Scholar] [CrossRef] [PubMed]
- Ulm, R.; Baumann, A.; Oravecz, A.; Máté, Z.; Adám, E.; Oakeley, E.J.; Schäfer, E.; Nagy, F. Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 5, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Molinier, J.; Oakeley, E.J.; Niederhauser, O.; Kovalchuk, I.; Hohn, B. Dynamic response of plant genome to ultraviolet radiation and other genotoxic stresses. Mutat. Res. 2005, 1–2, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.Y.; Hsu, P.H.; Chou, D.F.; Pan, C.Y.; Wang, L.C.; Huang, W.C.; Tsai, M.D.; Lo, W.S. The histone H3K36 demethylase Rph1/KDM4 regulates the expression of the photoreactivation gene PHR1. Nucleic Acids Res. 2011, 10, 4151–4165. [Google Scholar] [CrossRef] [PubMed]
- Questa, J.I.; Rius, S.P.; Casadevall, R.; Casati, P. ZmMBD101 is a DNA-binding protein that maintains Mutator elements chromatin in a repressive state in maize. Plant Cell Environ. 2016, 1, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Van Attikum, H.; Gasser, S.M. Crosstalk between histone modifications during the DNA damage response. Trends Cell Biol. 2009, 5, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J. Cell Biol. 1999, 5, 905–916. [Google Scholar] [CrossRef]
- Friesner, J.D.; Liu, B.; Culligan, K.; Britt, A.B. Ionizing radiation-dependent γ-H2AX focus formation requires ataxia telangiectasia mutated and ataxia telangiectasia mutated and Rad3-related. Mol. Biol. Cell 2005, 5, 2566–2576. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.Z.; Li, B.; Huang, B.; Wang, Y.; Liu, X.D.; Guan, H.; Zhang, S.M.; Tang, Y.; Rang, W.Q.; Zhou, P.K. γH2AX foci formation in the absence of DNA damage: Mitotic H2AX phosphorylation is mediated by the DNA-PKcs/CHK2 pathway. FEBS Lett. 2013, 21, 3437–3443. [Google Scholar] [CrossRef] [PubMed]
- Lorković, Z.J.; Park, C.; Goiser, M.; Jiang, D.; Kurzbauer, M.T.; Schlögelhofer, P.; Berger, F. Compartmentalization of DNA damage response between heterochromatin and euchromatin is mediated by distinct H2A histone variants. Curr. Biol. 2017, 8, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, Y.; Gao, J.; Yu, F.; Dong, A.; Shen, W.H. Molecular and reverse genetic characterization of nucleosome assembly protein 1 (NAP1) genes unravels their function in transcription and nucleotide excision repair in Arabidopsis thaliana. Plant J. 2009, 1, 27–38. [Google Scholar] [CrossRef]
- Gao, J.; Zhu, Y.; Zhou, W.; Molinier, J.; Dong, A.; Shen, W.H. NAP1 family histone chaperones are required for somatic homologous recombination in Arabidopsis. Plant Cell 2012, 4, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Varas, J.; Santos, J.L.; Pradillo, M. The absence of the Arabidopsis chaperone complex CAF-1 produces mitotic chromosome abnormalities and changes in the expression profiles of genes involved in DNA repair. Front. Plant Sci. 2017, 8, 525. [Google Scholar] [CrossRef] [PubMed]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 3, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Pandey-Rai, S. Deciphering UV-B-induced variation in DNA methylation pattern and its influence on regulation of DBR2 expression in Artemisia annua L. Planta 2015, 4, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Eichten, S.R.; Springer, N.M. Minimal evidence for consistent changes in maize DNA methylation patterns following environmental stress. Front. Plant Sci. 2015, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Rochette, P.J.; Lacoste, S.; Therrien, J.P.; Bastien, N.; Brash, D.E.; Drouin, R. Influence of cytosine methylation on ultraviolet-induced cyclobutane pyrimidine dimer formation in genomic DNA. Mutat. Res. 2009, 1–2, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. Photolyase and cryptochrome blue-light photoreceptors. Adv. Protein Chem. 2004, 69, 73–100. [Google Scholar] [CrossRef] [PubMed]
- Castells, E.; Molinier, J.; Drevensek, S.; Genschik, P.; Barneche, F.; Bowler, C. det1-1-induced UV-C hyposensitivity through UVR3 and PHR1 photolyase gene over-expression. Plant J. 2010, 3, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Schärer, O.D. Nucleotide excision repair in eukaryotes. Cold Spring Harb. Perspect. Biol. 2013, 10, a012609. [Google Scholar] [CrossRef] [PubMed]
- Chu, G.; Chang, E. Xeroderma pigmentosum group E cells lack a nuclear factor that binds to damaged DNA. Science 1988, 4878, 564–567. [Google Scholar] [CrossRef]
- Molinier, J.; Lechner, E.; Dumbliauskas, E.; Genschik, P. Regulation and role of Arabidopsis CUL4-DDB1A-DDB2 in maintaining genome integrity upon UV stress. PLoS Genet. 2008, 6, e1000093. [Google Scholar] [CrossRef] [PubMed]
- Wittschieben, B.Ø.; Iwai, S.; Wood, R.D. DDB1-DDB2 (xeroderma pigmentosum group E) protein complex recognizes a cyclobutane pyrimidine dimer, mismatches, apurinic/apyrimidinic sites, and compound lesions in DNA. J. Biol. Chem. 2005, 280, 39982–39989. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Douglas, L.; Zhou, P. UV-damaged DNA-binding proteins are targets of CUL-4A-mediated ubiquitination and degradation. J. Biol. Chem. 2001, 276, 48175–48182. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Masutani, C.; Takemura, M.; Uchida, A.; Sugasawa, K.; Kondoh, J.; Ohkuma, Y.; Hanaoka, F. Centrosome protein centrin 2/caltractin 1 is part of the xeroderma pigmentosum group C complex that initiates global genome nucleotide excision repair. J. Biol. Chem. 2001, 22, 18665–18672. [Google Scholar] [CrossRef] [PubMed]
- Castells, E.; Molinier, J.; Benvenuto, G.; Bourbousse, C.; Zabulon, G.; Zalc, A.; Cazzaniga, S.; Genschik, P.; Barneche, F.; Bowler, C. The conserved factor DE-ETIOLATED 1 cooperates with CUL4-DDB1DDB2 to maintain genome integrity upon UV stress. EMBO J. 2011, 6, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.D.; Shivji, M.K. Which DNA polymerases are used for DNA-repair in eukaryotes? Carcinogenesis 1997, 4, 605–610. [Google Scholar] [CrossRef]
- Avery, S.V. Molecular targets of oxidative stress. Biochem. J. 2011, 434, 201–210. [Google Scholar] [CrossRef] [PubMed]
- García-Ortiz, M.V.; Ariza, R.R.; Roldán-Arjona, T. An OGG1 orthologue encoding a functional 8-oxoguanine DNA glycosylase/lyase in Arabidopsis thaliana. Plant Mol. Biol. 2001, 6, 795–804. [Google Scholar] [CrossRef]
- Knoll, A.; Fauser, F.; Puchta, H. DNA recombination in somatic plant cells: Mechanisms and evolutionary consequences. Chromosome Res. 2014, 2, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Devos, K.M.; Brown, J.K.; Bennetzen, J.L. Genome size reduction through illegitimate recombination counteracts genome expansion in Arabidopsis. Genome Res. 2002, 7, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Virdi, K.S.; Laurie, J.D.; Xu, Y.Z.; Yu, J.; Shao, M.R.; Sanchez, R.; Kundariya, H.; Wang, D.; Riethoven, J.J.; Wamboldt, Y.; et al. Arabidopsis MSH1 mutation alters the epigenome and produces heritable changes in plant growth. Nat. Commun. 2015, 6, 6386. [Google Scholar] [CrossRef] [PubMed]
- Schalk, C.; Drevensek, S.; Kramdi, A.; Kassam, M.; Ahmed, I.; Cognat, V.; Graindorge, S.; Bergdoll, M.; Baumberger, N.; Heintz, D.; et al. DNA damage-binding protein 2 shapes the DNA methylation landscape. Plant Cell 2016, 28, 2043–2059. [Google Scholar] [CrossRef] [PubMed]
- Naegeli, H.; Sugasawa, K. The xeroderma pigmentosum pathway: Decision tree analysis of DNA quality. DNA Repair 2011, 10, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Le May, N.; Fradin, D.; Iltis, I.; Bougnères, P.; Egly, J.M. XPG and XPF endonucleases trigger chromatin looping and DNA demethylation for accurate expression of activated genes. Mol. Cell 2012, 47, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Vu, G.T. Genome stability and evolution: Attempting a holistic view. Trends Plant Sci. 2016, 9, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Puchta, H. The repair of double-strand breaks in plants: Mechanisms and consequences for genome evolution. J. Exp. Bot. 2005, 409, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Molinier, J.; Ries, G.; Zipfel, C.; Hohn, B. Transgeneration memory of stress in plants. Nature 2006, 442, 1046–1049. [Google Scholar] [CrossRef] [PubMed]
- Pecinka, A.; Rosa, M.; Schikora, A.; Berlinger, M.; Hirt, H.; Luschnig, C.; Mittelsten Scheid, O. Transgenerational stress memory is not a general response in Arabidopsis. PLoS ONE 2009, 4, e5202. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature 2011, 7341, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Pecinka, A.; Dinh, H.Q.; Baubec, T.; Rosa, M.; Lettner, N.; Mittelsten Scheid, O. Epigenetic regulation of repetitive elements is attenuated by prolonged heat stress in Arabidopsis. Plant Cell 2010, 9, 3118–3129. [Google Scholar] [CrossRef] [PubMed]
- Tittel-Elmer, M.; Bucher, E.; Broger, L.; Mathieu, O.; Paszkowski, J.; Vaillant, I. Stress-induced activation of heterochromatic transcription. PLoS Genet. 2010, 6, e1001175. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Habu, Y. siRNA-mediated chromatin maintenance and its function in Arabidopsis thaliana. Biochim. Biophys. Acta 2011, 1809, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Rigal, M.; Mathieu, O. A “mille-feuille” of silencing: Epigenetic control of transposable elements. Biochim. Biophys. Acta 2011, 1809, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Lepère, G.; Jay, F.; Wang, J.; Bapaume, L.; Wang, Y.; Abraham, A.L.; Penterman, J.; Fischer, R.L.; Voinnet, O.; et al. Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proc. Natl. Acad. Sci. USA 2013, 110, 2389–2394. [Google Scholar] [CrossRef] [PubMed]
- Pietzenuk, B.; Markus, C.; Gaubert, H.; Bagwan, N.; Merotto, A.; Bucher, E.; Pecinka, A. Recurrent evolution of heat-responsiveness in Brassicaceae COPIA elements. Genome Biol. 2016, 1, 209. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Kaczmarek, N.; Luch, A.; Glas, A.; Carell, T.; Naegeli, H. Regulation of nucleotide excision repair by UV-DDB: Prioritization of damage recognition to internucleosomal DNA. PLoS Biol. 2011, 10, e1001183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adar, S.; Hu, J.; Lieb, J.D.; Sancar, A. Genome-wide kinetics of DNA excision repair in relation to chromatin state and mutagenesis. Proc. Natl. Acad. Sci. USA 2016, 15, E2124–E2133. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, O.; Benvenuto, G.; Bowler, C.; Molinier, J.; Hohn, B. The INO80 protein controls homologous recombination in Arabidopsis thaliana. Mol. Cell 2004, 16, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Kirik, A.; Pecinka, A.; Wendeler, E.; Reiss, B. The Chromatin assembly factor subunit FASCIATA1 is involved in homologous recombination in Pplants. Plant Cell 2006, 18, 2431–2442. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Von Harder, M.; Aiese Cigliano, R.; Schlögelhofer, P.; Mittelsten Scheid, O. The Arabidopsis SWR1 chromatin-remodeling complex is important for DNA repair, somatic recombination, and meiosis. Plant Cell 2013, 25, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, T.; Huang, S.C.; Jupe, F.; Sasaki, E.; Schmitz, R.J.; Urich, M.A.; Castanon, R.; Nery, J.R.; Barragan, C.; He, Y.; et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 2016, 2, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Niederhuth, C.E.; Bewick, A.J.; Ji, L.; Alabady, M.S.; Kim, K.D.; Li, Q.; Rohr, N.A.; Rambani, A.; Burke, J.M.; Udall, J.A.; et al. Widespread natural variation of DNA methylation within angiosperms. Genome Biol. 2016, 1, 194. [Google Scholar] [CrossRef] [PubMed]
- Pecinka, A.; Abdelsamad, A.; Vu, G.T. Hidden genetic nature of epigenetic natural variation in plants. Trends Plant Sci. 2013, 11, 625–632. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinier, J. Genome and Epigenome Surveillance Processes Underlying UV Exposure in Plants. Genes 2017, 8, 316. https://doi.org/10.3390/genes8110316
Molinier J. Genome and Epigenome Surveillance Processes Underlying UV Exposure in Plants. Genes. 2017; 8(11):316. https://doi.org/10.3390/genes8110316
Chicago/Turabian StyleMolinier, Jean. 2017. "Genome and Epigenome Surveillance Processes Underlying UV Exposure in Plants" Genes 8, no. 11: 316. https://doi.org/10.3390/genes8110316
APA StyleMolinier, J. (2017). Genome and Epigenome Surveillance Processes Underlying UV Exposure in Plants. Genes, 8(11), 316. https://doi.org/10.3390/genes8110316