
The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants Materials and DNA Sequencing
2.2. Genome Annotation, Genome Mapping and Sequence Analysis
2.3. Repeat Structure
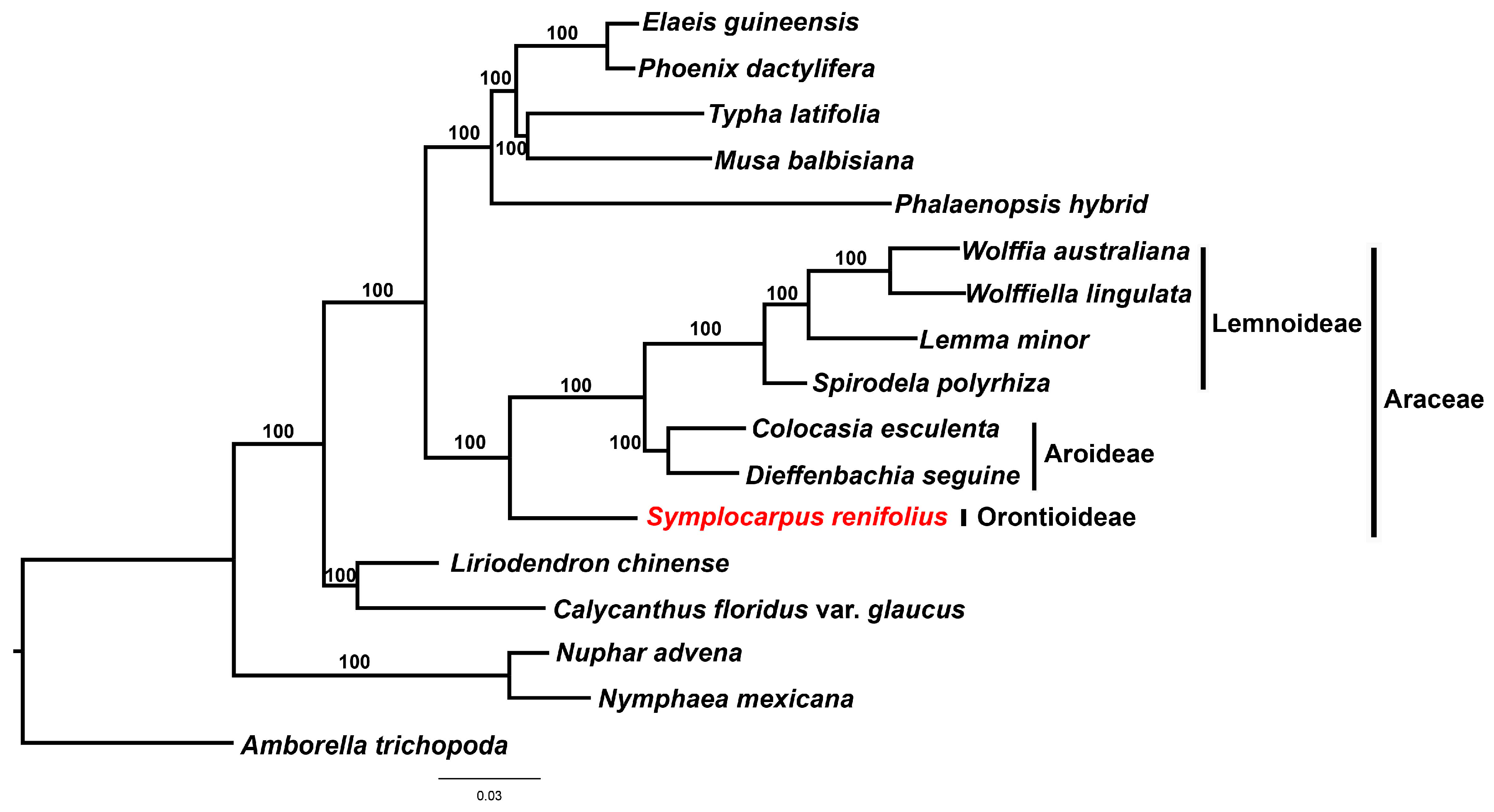
2.4. Phylogenetic Analysis
3. Results
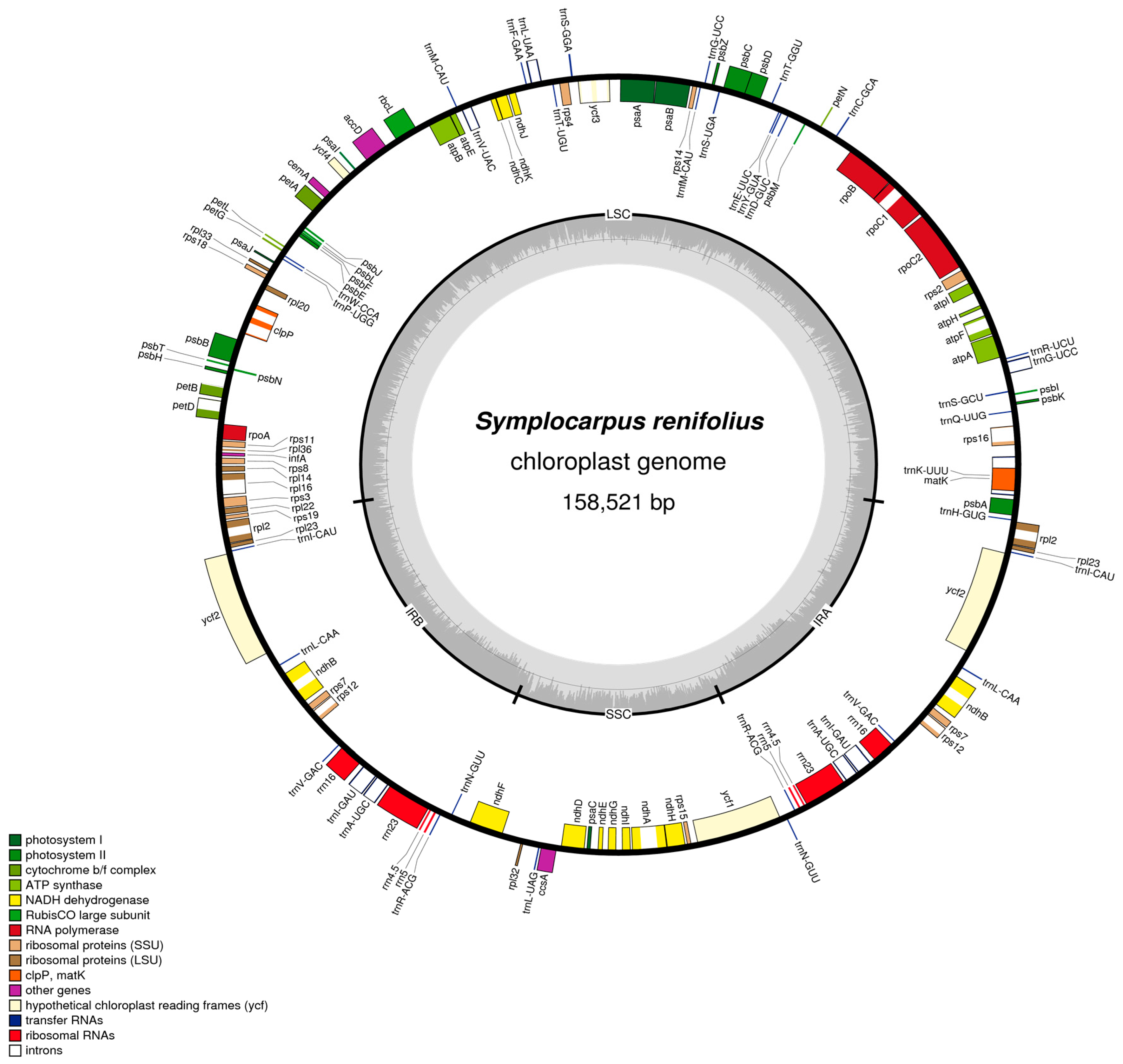
3.1. Characteristics of the Symplocarpus renifolius and Araceae cp Genomes
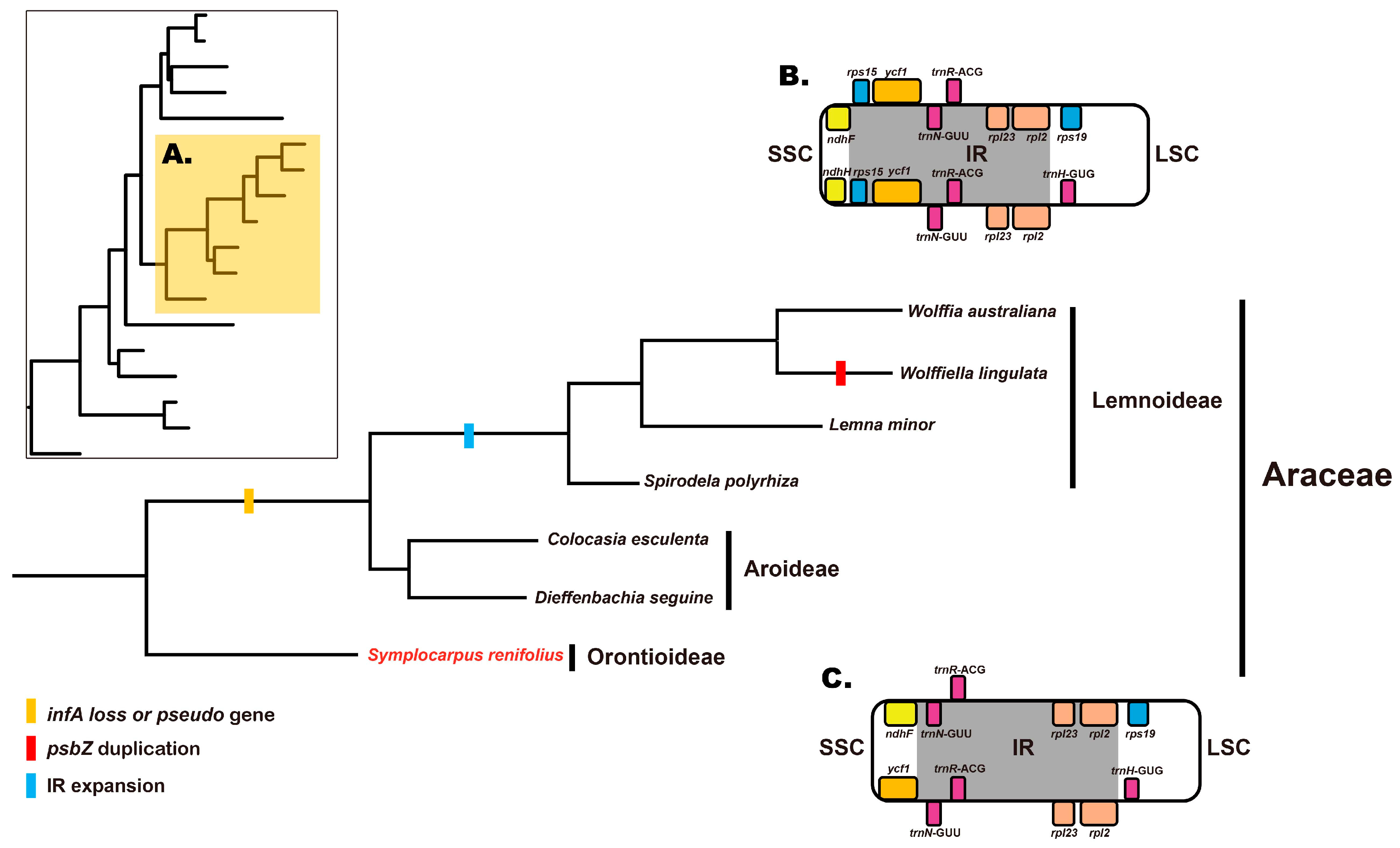
3.2. infA and psbZ Genes in Araceae
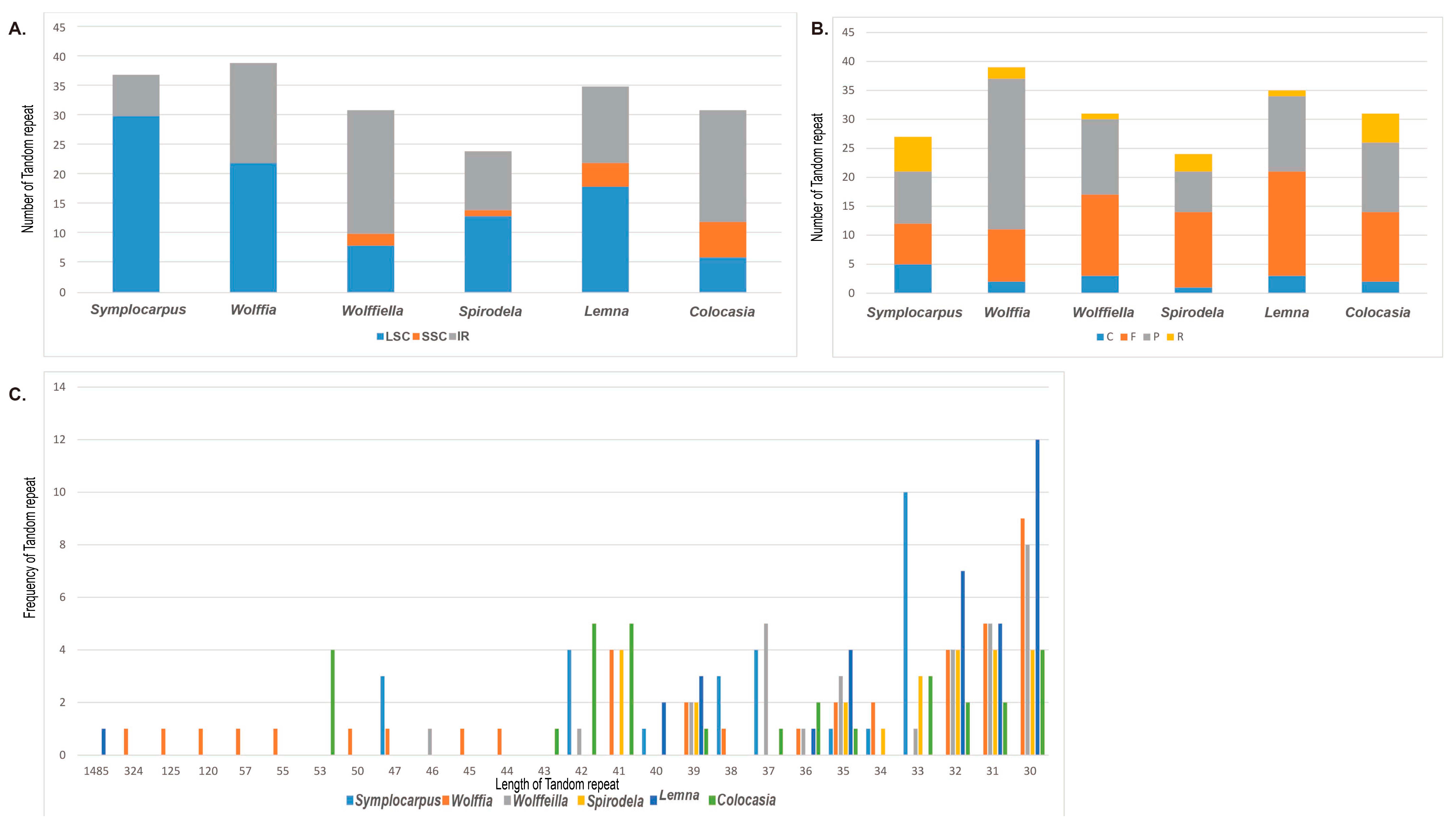
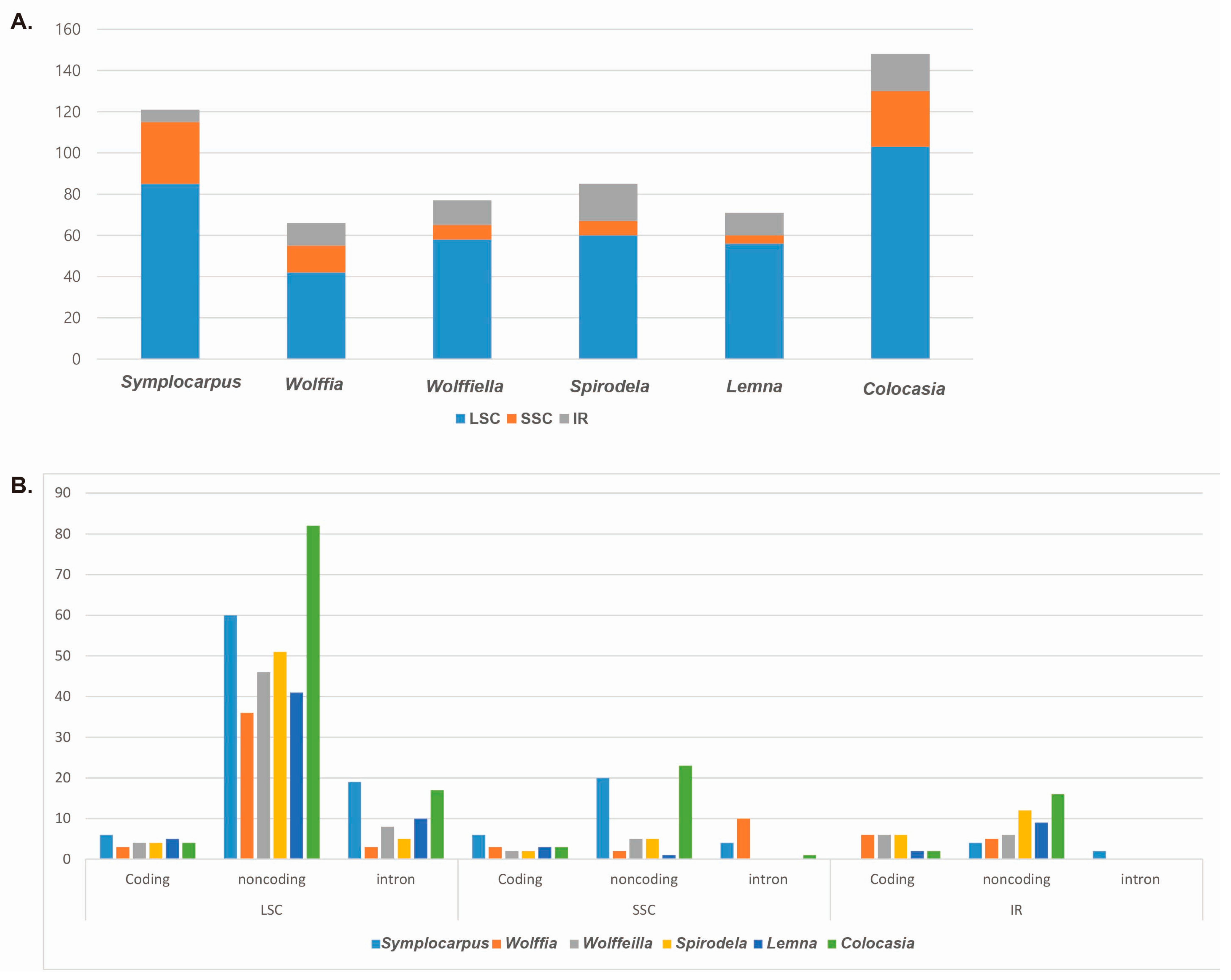
3.3. Simple-Sequence Repeats Loci of Araceae cp Genomes
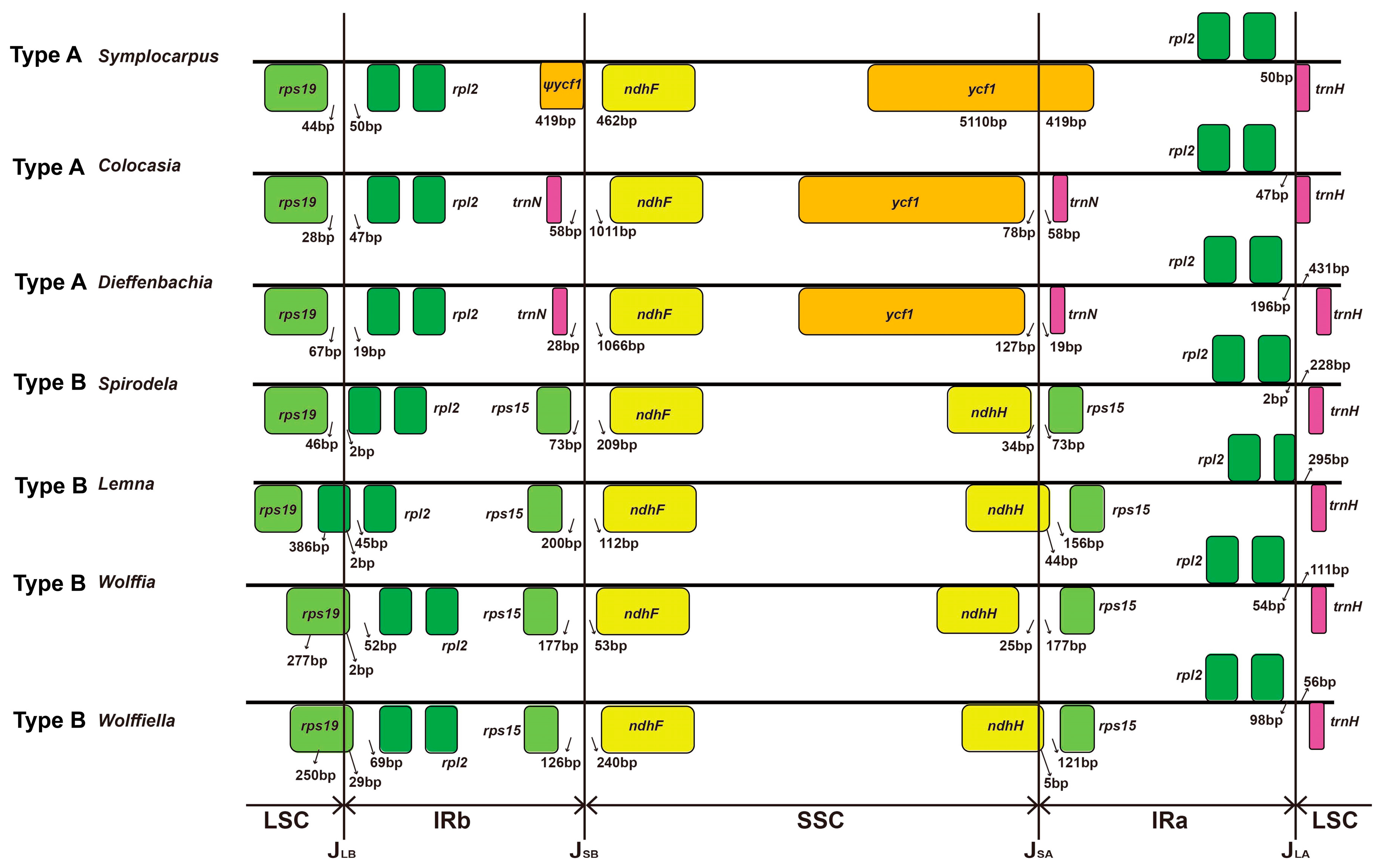
3.4. Two Types of Inverted Repeat Regions in Araceae
3.5. Phylogenetic Analysis of Araceae
4. Discussion
4.1. Gene Loss of infA and Gene Duplication of psbZ in Araceae
4.2. Phylogenetic Analysis and cp Structure in Araceae
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grayum, M.H. Evolution and phylogeny of the Araceae. Ann. Mo. Bot. Gard. 1990, 77, 628–697. [Google Scholar] [CrossRef]
- Les, D.H.; Crawford, D.J.; Landolt, E.; Gabel, J.D.; Kimball, R.T. Phylogeny and systematics of Lemnaceae, the duckweed family. Syst. Bot. 2002, 27, 221–240. [Google Scholar]
- Cusimano, N.; Bogner, J.; Mayo, S.J.; Boyce, P.C.; Wong, S.Y.; Hesse, M.; Hetterscheid, W.L.; Keating, R.C.; French, J.C. Relationships within the Araceae: Comparison of morphological patterns with molecular phylogenies. Am. J. Bot. 2011, 98, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Schott, H.W. Prodromus Systematis Aroidearum; Congretationis Mechitharisticae: Vienna, Austria, 1860. [Google Scholar]
- Engler, A. Vergleichende Untersuchungen über die morphologischen Verhältnisse der Araceae. I. Natürliches System der Araceae. In Nova Acta Academiae Caesareae Leopino-Carolinae Germanicae Naturae Curiosorum; E. Blochmann & Sohn: Dresden, Germany, 1876; Volume 39, pp. 133–155. [Google Scholar]
- Engler, A. Araceae-Pars generalis et index familiae generalis. In Das Pflanzenreich; Engler, A., Ed.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1920; Volume 74, (IV.23A); pp. 1–71. [Google Scholar]
- Barabé, D.; Lacroix, C.; Bruneau, A.; Forest, F.; Lacroix, C. The correlation between development of atypical bisexual flowers and phylogeny in the Aroideae (Araceae). Plant Syst. Evol. 2002, 232, 1–19. [Google Scholar] [CrossRef]
- Rothwell, G.W.; Van Atta, M.R.; Ballard, H.W., Jr.; Stockey, R.A. Molecular phylogenetic relationships among Lemnaceae and Araceae using the chloroplast trnL-trnF spacer. Mol. Phylogenet. Evol. 2004, 30, 378–385. [Google Scholar] [CrossRef]
- Mayo, S.J.; Bogner, J.; Bouce, P.C. The Genera of Araceae; Royal Botanic Gardens, Kew: London, UK, 1997. [Google Scholar]
- Wen, J.; Jansen, R.K.; Kilgore, K. Evolution of the eastern Asian and eastern North American disjunct genus Symplocarpus (Araceae): Insights from chloroplast DNA restriction site data. Biochem. Syst. Ecol. 1996, 24, 735–747. [Google Scholar] [CrossRef]
- Nie, Z.-L.; Sun, H.; Li, H.; Wen, J. Intercontinental biogeography of subfamily Orontioideae (Symplocarpus, Lysichiton and Orontium) of Araceae in eastern Asia and North America. Mol. Phylogenet. Evol. 2006, 40, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Contrasting modes and tempos of genome evolution in land plant organelles. Trends Genet. 1990, 6, 115–120. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; de Pamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; dePamphilis, C.W.; Leebens-Mack, J.; Müller, K.F.; Gusinger-Bellian, M.; Haberte, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Palmer, J.D. A chloroplast DNA inversion markers and ancient evolutionary split in the sunflower family (Asteraceae). Proc. Natl. Acad. Sci. USA 1987, 84, 5818–5822. [Google Scholar] [CrossRef] [PubMed]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The complete chloroplast genome sequence of Pelargonium X hortorum: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D.; Osorio, B.; Aldrich, J.; Thompson, W.F. Chloroplast DNA evolution among legumes: Loss of a large inverted repeat occurred prior to other sequence rearrangements. Curr. Genet. 1987, 11, 275–286. [Google Scholar] [CrossRef]
- Wang, R.J.; Cheng, C.L.; Chang, C.C.; Wu, C.L.; Su, T.M.; Chaw, S.M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Mardanov, A.V.; Ranin, N.V.; Kuznestsov, B.B.; Samigullin, T.H.; Antonov, A.S.; Kolganova, T.V.; Skyabin, K.G. Complete sequence of the Duckweed (Lemna minor) chloroplast genome: Structural organization and phylogenetic relationships to other Angiosperms. J. Mol. Evol. 2008, 66, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Messing, J. High-throughput sequencing of three Lemnoideae (Duckweeds) chloroplast genomes from total DNA. PLoS ONE 2011, 6, e24670. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Biggs, P.G.; Mattews, P.J.; Collins, L.J.; Hendy, M.D.; Lockhart, P.J. Mutational dynamics of Aroid chloroplast genomes. Genome Biol. Evol. 2012, 4, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Schleiermacher, C. REPuter: Fast computation of maximal repeats in complete genomes. Bioinformatics 1999, 15, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Kavanagh, T.A.; Hibberd, J.M.; Grey, J.C.; Morden, C.W.; et al. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jansen, R.K.; Park, S. Complete plastome sequence of Thalictrum coreanum (Ranunculaceae) and transfer of the rpl32 gene to the nucleus in the ancestor of the subfamily Thalictroideae. BMC Plant Biol. 2015, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Son, O.G.; Park, S.J. The chloroplast genome of Elaeagnus macrophylla and trnH duplication event in Elaeagnaceae. PLoS ONE 2015, 10, e0138727. [Google Scholar] [CrossRef] [PubMed]
- Ruhlman, T.A.; Jansen, R.K. The plastid genomes of flowering plants. In Chloroplast Biotechnology: Methods and Protocols; Maliga, P., Ed.; Springer Science and Business Media, LLC: New York, NY, USA, 2014; Volume 1132, pp. 3–38. [Google Scholar]
- Swiatek, M.; Kuras, R.; Sokolenko, A.; Higgs, D.; Olive, J.; Cinque, G.; Müller, B.; Eichacker, L.A.; Stem, D.B.; Bassi, R.; et al. The chloroplast gene ycf9 encodes a photosystem II (PSII) core subunit, PsbZ, that participates in PSII supramolecular architecture. Plant Cell 2001, 13, 1347–1367. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Yocum, C.F. Structure and function of photosystems I and II. Annu. Rev. Plant Biol. 2006, 57, 521–565. [Google Scholar] [CrossRef] [PubMed]
- McNeal, J.R.; Kuehl, J.; Boore, J.; Leebens-Mack, J.H.; de Pamphilis, C.W. Complete plastid genome sequences suggest strong selection for retention of photosynthetic genes in the parasitic plant genus Cuscuta. BMC Plant Biol. 2007, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Wickett, N.J.; Fan, Y.; Lewis, P.; Goffinet, B. Distribution and evolution of pseudogenes, gene losses and a gene rearrangement in the plastid genome of the nonphotosynthetic liverwort, Aneura mirabilis (Metzgeriales, Jungermanniopsida). J. Mol. Evol. 2008, 67, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA 1992, 89, 10648–10652. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, L.I.; Salazar, G.A.; Chase, M.W.; Mayo, S.J.; Bogner, J.; Dávila, P. Phylogenetic relationships of aroids and duckweeds (Araceae) inferred from coding and noncoding plastid DNA. Am. J. Bot. 2008, 95, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, G.M.; Dawnie, S.R. Expansion and contraction of the chloroplast inverted repeat in Apiaceae subfamily Apioideae. Syst. Bot. 2000, 25, 648–667. [Google Scholar] [CrossRef]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.R.; Meng, B.Y.; et al. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distint tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol. Gen. Genet. 1989, 217, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.M.; Neckermann, K.; Igloi, G.L.; Kössel, H. Complete sequence of the Maize chloroplast genome: Gene content, hotspots of divergence and fine tuning of genetic information by transcript editing. J. Mol. Biol. 1995, 241, 614–628. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symplocarpus | Colocasia | Dieffenbachia | Lemna | Spirodela | Wolffiella | Wolffia | |
|---|---|---|---|---|---|---|---|
| Total length | 158,521 bp | 162,424 bp | 163,699 bp | 165,955 bp | 168,788 bp | 169,337 bp | 168,704 bp |
| LSC | 86,620 bp | 86,670 bp | 90,780 bp | 89,906 bp | 91,222 bp | 92,015 bp | 91,454 bp |
| SSC | 20,299 bp | 22,208 bp | 22,440 bp | 13,603 bp | 14,056 bp | 13,956 bp | 13,394 bp |
| IR | 25,801 bp | 25,273 bp | 25,235 bp | 31,223 bp | 31,755 bp | 31,683 bp | 31,930 bp |
| % AT content | 62.7% | 63.8% | 63.6% | 64.3% | 64.3% | 64.2% | 64.1% |
| Genes | |||||||
| Coding genes | 80 | 79 | 79 | 79 | 79 | 79 | 79 |
| tRNA | 30 | 30 | 30 | 30 | 30 | 30 | 30 |
| rRNA | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| IR duplication genes | 17 | 17 | 17 | 17 | 19 | 19 | 19 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, K.S.; Park, K.T.; Park, S. The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae. Genes 2017, 8, 324. https://doi.org/10.3390/genes8110324
Choi KS, Park KT, Park S. The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae. Genes. 2017; 8(11):324. https://doi.org/10.3390/genes8110324
Chicago/Turabian StyleChoi, Kyoung Su, Kyu Tae Park, and SeonJoo Park. 2017. "The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae" Genes 8, no. 11: 324. https://doi.org/10.3390/genes8110324
APA StyleChoi, K. S., Park, K. T., & Park, S. (2017). The Chloroplast Genome of Symplocarpus renifolius: A Comparison of Chloroplast Genome Structure in Araceae. Genes, 8(11), 324. https://doi.org/10.3390/genes8110324