Clavine Alkaloids Gene Clusters of Penicillium and Related Fungi: Evolutionary Combination of Prenyltransferases, Monooxygenases and Dioxygenases



Abstract
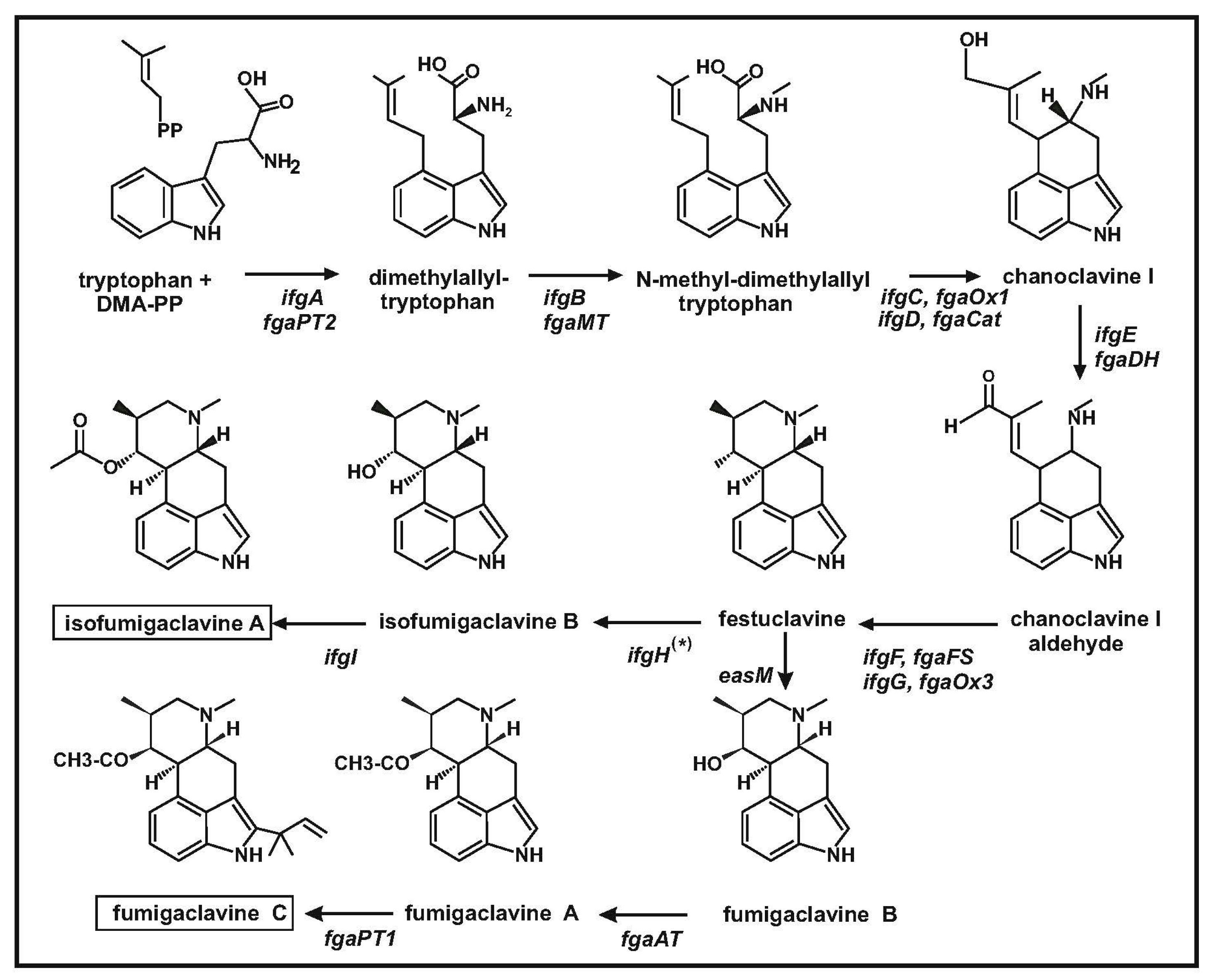
1. Clavine and Ergot Alkaloids: An Overview
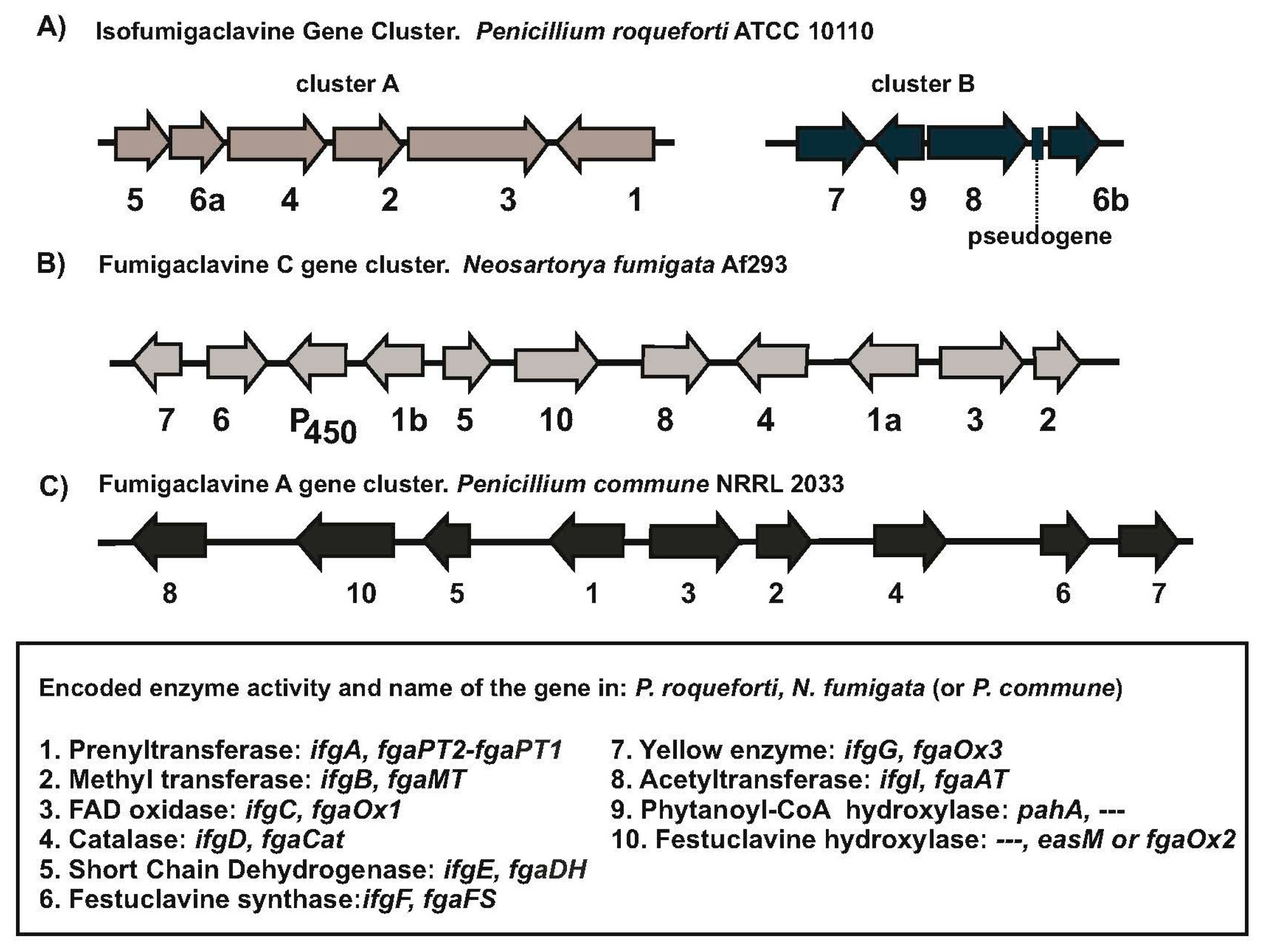
2. The Clavine Gene Cluster in Penicillium roqueforti: Comparison with Neosartorya fumigata
2.1. The Clavine Biosynthetic Genes of Penicillium roqueforti Are Located into Two Clusters
2.2. Two Yellow Enzymes Genes Exist in Penicillium roqueforti and Neosartorya fumigata
2.3. Penicillium roqueforti Synthesize Both Festuclavine and Agroclavine: The Role of Tyrosine versus Phenylalanine in the Yellow Enzyme Active Center
2.4. A Fe2+ and α-Ketoglutarate-Dependent Dioxygenase Is Encoded in Cluster B of Penicillium roqueforti
3. A Fumigaclavine Gene Cluster in Penicillium commune
4. A Cycloclavine Gene Cluster in Aspergillus japonicus and Byssochlamys spectabilis
5. A Chanoclavine I Gene Cluster Exist in Members of the Arthrodermataceae Family
6. Role of Non-Clustered Non-Ribosomal Peptide Synthetases on Fumigaclavine C Biosynthesis
7. The Penicillium and Other Aspergillaceae and Arthrodermataceae Fungi Lack the Conversion of Agroclavine to Lysergic Acid
8. Evolutionary Relationship between Clavine and Ergot Alkaloids: A Recruiting Play of Prenyltransferases, Monooxygenases and Dioxygenases
8.1. Gene Gains, Gene Losses and Gene Rearrangement in the Clavine Alkaloid Producers
8.2. Plasticity of the Clusters: Clavine Gene Clusters in Penicillium Species
8.2.1. Clavine Gene Cluster in Penicillium camemberti
8.2.2. Clavine Gene Cluster in Penicillium expansum
8.2.3. Clavine Gene Cluster in Penicillium steckii
8.2.4. Clavine Gene Cluster in Penicillium griseofulvum
8.3. Role and Evolution of Phytanoyl-CoA Hydroxylase Enzymes in Clavine and Ergot Alkaloid Biosynthesis
8.4. Vertical Gene Diversification versus Horizontal Gene Transfer
Acknowledgments
Conflicts of Interest
References
- Xu, J.; Song, Y.C.; Guo, Y.; Mei, Y.N.; Tan, R.X. Fumigaclavines D-H, new ergot alkaloids from endophytic Aspergillus fumigatus. Planta Med. 2014, 80, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.M.; Yu, Z.G.; Zhang, J.; Wu, J.H.; Tan, R.X. Bioactive alkaloids from endophytic Aspergillus fumigatus. J. Nat. Prod. 2009, 72, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Coyle, C.M.; Cheng, J.Z.; O’Connor, S.E.; Panaccione, D.G. An old yellow enzyme gene controls the branch point between Aspergillus fumigatus and Claviceps purpurea ergot alkaloid pathways. Appl. Environ. Microbiol. 2010, 76, 3898–3903. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Matuschek, M.; Xie, X.L.; Li, S.M. Ergot alkaloid biosynthesis in Aspergillus fumigatus: Conversion of chanoclavine-I aldehyde to festuclavine by the festuclavine synthase FgaFS in the presence of the old yellow enzyme FgaOx3. Org. Biomol. Chem. 2010, 8, 3500–3508. [Google Scholar] [CrossRef] [PubMed]
- Bilovol, Y.; Panaccione, D.G. Functional analysis of the gene controlling hydroxylation of festuclavine in the ergot alkaloid pathway of Neosartorya fumigata. Curr. Genet. 2016, 62, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Unsöld, I.A. Molecular Biological and Biochemical Investigations on the Biosynthesis of Fumigaclavines in Aspergillus fumigatus AF 293/B 5233 and Penicillium commune NRRL2033. Ph.D. Dissertation, University of Tubingen, Tübingen, Germany, 2006. [Google Scholar]
- Jakubczyk, D.; Caputi, L.; Hatsch, A.I.; Nielsen, C.A.F.; Diefenbacher, M.; Klein, J.; Molt, A.; Schröder, H.; Cheng, H.J.; Naesby, M.; et al. Discovery and Reconstitution of the Cycloclavine Biosynthetic Pathway. Enzymatic Formation of a Cyclopropyl Group. Angew. Chem. Int. Ed. 2015, 54, 5117–5121. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, D.; Caputi, L.; Stevenson, C.E.; Lawson, D.M.; O’Connor, S.E. Structural characterization of EasH (Aspergillus japonicus)—An oxidase involved in cycloclavine biosynthesis. Chem. Commun. 2016, 52, 14306–14309. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Heddergott, C.; Xie, X.; Brakhage, A.A.; Li, S.-M. Genome mining reveals the presence of a conserved gene cluster for the biosynthesis of ergot alkaloid precursors in the fungal family Arthrodermataceae. Microbiology 2012, 158, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bodega, Á.; Álvarez-Álvarez, R.; Liras, P.; Martín, J.F. Silencing of a second dimethylallyltryptophan synthase of Penicillium roqueforti reveals a novel clavine alkaloid gene cluster. Appl. Microbiol. Biotechnol. 2017, 101, 6111–6121. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.L.; Panaccione, D.G. Diversification of ergot alkaloids in natural and modified fungi. Toxins 2015, 7, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. Ergot alkaloids—Biology and molecular biology. Alkaloids Chem. Biol. 2006, 63, 45–86. [Google Scholar] [PubMed]
- Florea, S.; Panaccione, D.G.C.L. Ergot Alkaloids of the Family Clavicipitaceae. Phytopathology 2017, 107, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Li, S.M. Ergot alkaloids: structure diversity, biosynthetic gene clusters and functional proof of biosynthetic genes. Nat. Prod. Rep. 2011, 28, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Ohmomo, S.; Sato, T.; Utagawa, T.; Abe, M. Production of alkaloids and related substances by fungi. Isolation of festuclavine and three new indole alkaloids, roquefortine A, B, and C from cultures of Penicillium roqueforti. Nippon Nogei Kagaku Kaishi 1975, 49, 615–623. [Google Scholar] [CrossRef]
- O’Brien, M.; Nielsen, K.F.; O’Kiely, P.; Forristal, P.D.; Fuller, H.T.; Frisvad, J.C. Mycotoxins and other secondary metabolites produced in vitro by Penicillium paneum Frisvad and Penicillium roqueforti Thom isolated from baled grass silage in Ireland. J. Agric. Food Chem. 2006, 54, 9268–9276. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Sumarah, M.W.; Frisvad, J.C.; Miller, J.D. Production of metabolites from the Penicillium roqueforti complex. J. Agric. Food Chem. 2006, 54, 3756–3763. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Liras, P. Secondary Metabolites in Cheese Fungi. In Reference Series in Phytochemistry: Fungal Metabolites; Merillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 293–315. [Google Scholar]
- García-Estrada, C.; Martín, J.F. Biosynthetic gene clusters for relevant secondary metabolites produced by Penicillium roqueforti in blue cheeses. Appl. Microbiol. Biotechnol. 2016, 100, 8303–8313. [Google Scholar] [CrossRef] [PubMed]
- Kosalková, K.; Domínguez-Santos, R.; Coton, M.; Coton, E.; García-Estrada, C.; Liras, P.; Martín, J.F. A natural short pathway synthesizes roquefortine C but not meleagrin in three different Penicillium roqueforti strains. Appl. Microbiol. Biotechnol. 2015, 99, 7601–7612. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-M. Evolution of aromatic prenyltransferases in the biosynthesis of indole derivatives. Phytochemistry 2009, 70, 1746–1757. [Google Scholar] [CrossRef] [PubMed]
- Unsöld, I.A.; Li, S.M. Overproduction, purification and characterization of FgaPT2, a dimethylallyltryptophan synthase from Aspergillus fumigatus. Microbiology 2005, 151, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Unsöld, I.A.; Li, S.M. Reverse prenyltransferase in the biosynthesis of fumigaclavine C in Aspergillus fumigatus: Gene expression, purification, and characterization of fumigaclavine C synthase FGAPT1. Chembiochem 2006, 7, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Fan, A.; Li, S.-M. Saturation mutagenesis on Arg244 of the tryptophan C4-prenyltransferase FgaPT2 leads to enhanced catalytic ability and different preferences for tryptophan-containing cyclic dipeptides. Appl. Microbiol. Biotechnol. 2016, 100, 5389–5399. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.L.; Panaccione, D.G. Heterologous expression of lysergic acid and novel ergot alkaloids in Aspergillus fumigatus. Appl. Environ. Microbiol. 2014, 80, 6465–6472. [Google Scholar] [CrossRef] [PubMed]
- Rigbers, O.; Li, S.M. Ergot alkaloid biosynthesis in Aspergillus fumigatus. Overproduction and biochemical characterization of a 4-dimethylallyltryptophan N-methyltransferase. J. Biol. Chem. 2008, 283, 26859–26868. [Google Scholar] [CrossRef] [PubMed]
- Goetz, K.E.; Coyle, C.M.; Cheng, J.Z.; O’Connor, S.E.; Panaccione, D.G. Ergot cluster-encoded catalase is required for synthesis of chanoclavine-I in Aspergillus fumigatus. Curr. Genet. 2011, 57, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.A.F.; Folly, C.; Hatsch, A.; Molt, A.; Schröder, H.; O’Connor, S.E.; Naesby, M. The important ergot alkaloid intermediate chanoclavine-I produced in the yeast Saccharomyces cerevisiae by the combined action of EasC and EasE from Aspergillus japonicus. Microb. Cell Fact. 2014, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Matuschek, M.; Li, S.M. Ergot alkaloid biosynthesis in Aspergillus fumigatus: Conversion of chanoclavine-I to chanoclavine-I aldehyde catalyzed by a short-chain alcohol dehydrogenase FgaDH. Arch. Microbiol. 2010, 192, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gerhards, N.; Li, S.M. A bifunctional old yellow enzyme from Penicillium roqueforti is involved in ergot alkaloid biosynthesis. Org. Biomol. Chem. 2017, 15, 8059–8071. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.Z.; Coyle, C.M.; Panaccione, D.G.; O’Connor, S.E. Controlling a structural branch point in ergot alkaloid biosynthesis. J. Am. Chem. Soc. 2010, 22, 12835–12837. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, K.; Ropars, J.; Renault, P.; Dupont, J.; Gouzy, J.; Branca, A.; Abraham, A.L.; Ceppi, M.; Conseiller, E.; Debuchy, R.; et al. Multiple recent horizontal transfers of a large genomic region in cheese making fungi. Nat. Commun. 2014, 5, 2876. [Google Scholar] [CrossRef] [PubMed]
- Matuschek, M.; Wallwey, C.; Xie, X.; Li, S.M. New insights into ergot alkaloid biosynthesis in Claviceps purpurea: An agroclavine synthase EasG catalyses, via a non-enzymatic adduct with reduced glutathione, the conversion of chanoclavine-I aldehyde to agroclavine. Org. Biomol. Chem. 2011, 9, 4328–4335. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.R.; López-Nieto, M.J.; Martín, J.F. Isopenicillin N synthetase of Penicillium chrysogenum, an enzyme that converts delta-(l-alpha-aminoadipyl)-l-cysteinyl-d-valine to isopenicillin N. Antimicrob. Agents Chemother. 1985, 27, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Liras, P. Insights into the structure and Molecular Mechanisms of β-lactam synthesizing enzymes in fungi. In Biotechnology of Microbial Enzymes; Brahmachati, G., Demain, A.L., Adrio, J.L., Eds.; Elsevier: New York, NY, USA, 2016; pp. 215–241. [Google Scholar]
- Hausinger, R.P. FeII/alpha-ketoglutarate-dependent hydroxylases and related enzymes. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 21–68. [Google Scholar] [CrossRef] [PubMed]
- Havemann, J.; Vogel, D.; Loll, B.; Keller, U. Cyclolization of d-lysergic acid alkaloid peptides. Chem. Biol. 2014, 21, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, M.; Schofield, C.J.; Wierzbicki, A.S.; Jansen, G.A.; Wanders, R.J.; Lloyd, M.D. The chemical biology of branched-chain lipid metabolism. Prog. Lipid Res. 2003, 42, 359–376. [Google Scholar] [CrossRef]
- McDonough, M.A.; Kavanagh, K.L.; Butler, D.; Searls, T.; Oppermann, U.; Schofield, C.J. Structure of Human Phytanoyl-CoA 2-Hydroxylase Identifies Molecular Mechanisms of Refsum Disease. J. Biol. Chem. 2005, 280, 41101–41110. [Google Scholar] [CrossRef] [PubMed]
- Vinokurova, N.G.; Boichenko, L.V.; Arinbasarov, M.U. Production of alkaloids by fungi of the genus Penicillium grown on wheat grain. Appl. Biochem. Microbiol. 2003, 39, 403–406. [Google Scholar] [CrossRef]
- Gerhards, N.; Matuschek, M.; Wallwey, C.; Li, S.M. Genome mining of ascomycetous fungi reveals their genetic potential for ergot alkaloid production. Arch. Microbiol. 2015, 197, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Stauffacher, D.; Niklaus, P.; Tscherter, H.; Weber, H.P.; Hofmann, A. Cycloclavin, a new alkaloid from Ipomoea hildebrandtii Vatke. Tetrahedron 1969, 25, 5879–5887. [Google Scholar] [CrossRef]
- Furuta, T.; Koike, M.; Abe, M. Isolation of cycloclavine from the culture broth of Aspergillus japonicus SAITO. Agric. Biol. Chem. 1982, 46, 1921–1922. [Google Scholar] [CrossRef]
- Oka, T.; Ekino, K.; Fukuda, K.; Nomura, Y. Draft Genome Sequence of the Formaldehyde resistant Fungis Byssochlamys spectabilis No. 5 (Anamorph Paecelomyces variotii No. 5) (NBRC109023). Genome Announc. 2014, 2, e01162-13. [Google Scholar] [CrossRef] [PubMed]
- Burmester, A.; Shelest, E.; Glöckner, G.; Heddergott, C.; Schindler, S.; Staib, P.; Heidel, A.; Felder, M.; Petzold, A.; Szafranski, K. Comparative and functional genomics provide insights into the pathogenicity of dermatophytic fungi. Genome Biol. 2011, 12, R7. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Liras, P. Novel Antimicrobial and other Bioactive Metabolites obtained from Silent Gene Clusters. In Antibiotics: Current Innovations and Future Trends; Demain, A.L., Sanchez, S., Eds.; Horizon Scientific Press and Caister Academic Press: Poole, UK, 2014; pp. 275–292. [Google Scholar]
- O’Hanlon, K.A.; Gallagher, L.; Schretti, M.; Jöchl, C.; Kananagh, K.; Larsen, T.O.; Doyle, S. Non ribosomal peptide synthetase genes pesL and pes1 are essential for fumigaclavine C production in Aspergillus fumigatus. Appl. Envon. Microbiol. 2012, 78, 3166–3176. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.D.; Haynes, S.W.; Gao, X.; Evans, B.S.; Kelleher, N.L.; Tang, Y.; Walsh, C.T. Complexity Generation in Fungal Peptidyl Alkaloid Biosynthesis: Oxidation of Fumiquinazoline A to the Heptacyclic Hemiaminal Fumiquinazoline C by the Flavoenzyme Af12070 from Aspergillus fumigatus. Biochemistry 2011, 50, 8756–8769. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.; Funk, A.N.; Scherlach, K.; Schroeckh, V.; Shelest, E.; Horn, U.; Hertweck, C.; Brakhage, A.A. Activation of a silent fungal polyketide biosynthesis pathway through regulatory cross talk with a cryptic nonribosomal peptide synthetase gene cluster. Appl. Environ. Microbiol. 2010, 76, 8143–8149. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, P.I.; Ullán, R.V.; Albillos, S.M.; Montero, O.; Fernández-Bodega, M.Á.; García-Estrada, C.; Fernández-Aguado, M.; Martín, J.F. Molecular characterization of the PR-toxin gene cluster in Penicillium roqueforti and Penicillium chrysogenum: Cross talk of secondary metabolite pathways. Fungal Genet. Biol. 2014, 62, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, T.; Ortel, I.; Tudzynski, P.; Keller, U. Identification of the cytochrome P450 monooxygenase that bridges the clavine and ergoline alkaloid pathways. ChemBioChem 2006, 7, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.C.; Grijseels, S.; Pringent, S.; Ji, B.; Dainat, J.; Nielsen, K.F.; Frisvad, J.C.; Workman, M.; Nielsen, J. Global analysis of biosynthetic gene clusters reveals vast potential of secondary metabolite production in Penicillium species. Nat. Microbiol. 2017, 2, 17044. [Google Scholar] [CrossRef] [PubMed]
- Frisvasd, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Ballester, A.R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, Transcriptome, and Functional Analyses of Penicillium expansum Provide New Insights into Secondary Metabolism and Pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Banani, H.; Marcet-Houben, M.; Ballester, A.R.; Abbruscato, P.; González-Candelas, L.; Gabaldón, T.; Spadaro, D. Genome sequencing and secondary metabolism of the postharvest pathogen Penicillium griseofulvum. BMC Genom. 2016, 5, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Acklin, W.; Fehre, T.; Arigoni, D. The stereochemistry of chanoclavine I and isochanoclavine I. J. Chem. Soc. Chem. Commun. 1966, 1966, 799–800. [Google Scholar] [CrossRef]
- Fitzpatrick, D.A. Horizontal gene transfer in fungi. FEMS Microbiol. Lett. 2012, 329, 1–8. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name in P. roqueforti 1 | Function | N. fumigata (%) 2 | A. japonicus (%) | P. camemberti (%) | P. expansum (%) | P. steckii (%) | P. griseofulvum (%) |
|---|---|---|---|---|---|---|---|
| ifgA, CDM36678 | DMAT | fgaPT2, dmaW (64%) | dmaW (66%) | CRL19777 (84%) | XP_16600812 (60%) | OQE13746 (83%) | KXG48664 (60%) |
| ifgB, CDM36676 | Methyl Transferase | fgaMT, easF (63%) | easF (66%) | CRL19775 (85%) | XP_16604736 (53%) | OQE13752 (80%) | |
| ifgC, CDM36677 | FAD oxidase | fgaOx1, easE (49%) | easE (48%) | CRL19776 (77%) | XP_16600807 (47%) | OQE13752 (69%) | KXG48659 (44%) |
| ifgD, CDM36675 | Catalase | fgaCat, easC (62%) | easC (61%) | CRL19774 (90%) | XP_16600810 (63%) | OQE13751 (84%) | KXG48662 (60%) |
| ifgE, CDM36673 | Short Chain DH | fgaDH, easD (72%) | easD (67%) | CRL19778 (66%) | XP_16604733 (67%) | OQE13747 (68%) | |
| ifgG, CDM30151 | Yellow Enzyme | fgaOx3, easA (67%) | easA (64%) | CRL20441 (50%) | |||
| ifgF1, CDM36674 | Festuclavine Synthase I | fgaSF, easG (60%) | easG (54%) | XP_16604735 (49%) | OQE13753 (58%) | ||
| ifgF2, CDM30155 | Festuclavine Synthase II 3 | ||||||
| ifgI, CDM30153 | Acetyltransferase | fgaAT, easN (32%) | KXG 48657 (30%) | ||||
| Festuclavine Hydroxylase (P450) 4 | fgaOx2, easM | ||||||
| pahA, CDM30152 | Phytanoyl-CoA Hydroxylase | easH (34%) | CRL19773 (36%) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, J.F.; Álvarez-Álvarez, R.; Liras, P. Clavine Alkaloids Gene Clusters of Penicillium and Related Fungi: Evolutionary Combination of Prenyltransferases, Monooxygenases and Dioxygenases. Genes 2017, 8, 342. https://doi.org/10.3390/genes8120342
Martín JF, Álvarez-Álvarez R, Liras P. Clavine Alkaloids Gene Clusters of Penicillium and Related Fungi: Evolutionary Combination of Prenyltransferases, Monooxygenases and Dioxygenases. Genes. 2017; 8(12):342. https://doi.org/10.3390/genes8120342
Chicago/Turabian StyleMartín, Juan F., Rubén Álvarez-Álvarez, and Paloma Liras. 2017. "Clavine Alkaloids Gene Clusters of Penicillium and Related Fungi: Evolutionary Combination of Prenyltransferases, Monooxygenases and Dioxygenases" Genes 8, no. 12: 342. https://doi.org/10.3390/genes8120342
APA StyleMartín, J. F., Álvarez-Álvarez, R., & Liras, P. (2017). Clavine Alkaloids Gene Clusters of Penicillium and Related Fungi: Evolutionary Combination of Prenyltransferases, Monooxygenases and Dioxygenases. Genes, 8(12), 342. https://doi.org/10.3390/genes8120342