
Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sheep Investigated and Data Collection
2.2. Blood Sample and DNA Purification
2.3. PCR Amplification and Single-Stranded Conformational Polymorhipsm Analysis
2.4. Haplotype Determination
2.5. Statistical Analyses
3. Results
3.1. Frequencies of the Ovine ADIPOQ Haplotypes
3.2. Association of the Ovine ADIPOQ Haplotypes with Growth Traits
3.3. Association of the Ovine ADIPOQ Haplotypes with Carcass Traits
3.4. Diplotype Analyses
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shinoda, Y. Regulation of bone formation by adiponectin through autocrine/paracrine and endocrine pathways. Nippon Rinsho Jpn. J. Clin. Med. 2006, 99, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Morsci, N.S.; Schnabel, R.D.; Taylor, J.F. Association analysis of adiponectin and somatostatin polymorphisms on BTA1 with growth and carcass traits in Angus cattle. Anim. Genet. 2006, 37, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Arita, Y. Genomic structure and mutations in adipose-specific gene, adiponectin. Int. J. Obes. 2000, 24, 861–868. [Google Scholar]
- Dall’Olio, S.; Davoli, R.; Buttazzoni, L.; Zambonelli, P.; Russo, V. Study of porcine adiponectin (ADIPOQ) gene and association of a missense mutation with EBVs for production and carcass traits in Italian Duroc heavy pigs. Livest. Sci. 2009, 125, 101–104. [Google Scholar] [CrossRef]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A Novel Serum Protein Similar to C1q, Produced Exclusively in Adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, W.C.; St Jean, P.L.; Mitchell, B.D.; Pollin, T.I.; Knowler, W.C.; Ehm, M.G.; Bell, C.J.; Sakul, H.; Wagner, M.J.; Burns, D.K.; et al. Genome-wide and fine-mapping linkage studies of type 2 diabetes and glucose traits in the Old Order Amish: Evidence for a new diabetes locus on chromosome 14q11 and confirmation of a locus on chromosome 1q21-q24. Diabetes 2003, 52, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Wang, M.; Zhong, D.; Shi, D.; Ma, L.; Tong, N.; Zhang, Z. AdipoQ polymorphisms are associated with type 2 diabetes mellitus: A meta-analysis study. Diabetes 2013, 29, 532–545. [Google Scholar]
- Yang, Y.; Zhang, F.; Ding, R.; Skrip, L.; Wang, Y.; Lei, H.; Hu, D. ADIPOQ gene polymorphisms and cancer risk: A meta-analysis. Cytokine 2013, 61, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Ramya, K.; Ayyappa, K.A.; Ghosh, S.; Mohan, V.; Radha, V. Genetic association of ADIPOQ gene variants with type 2 diabetes, obesity and serum adiponectin levels in south Indian population. Gene 2013, 532, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jiang, H.; Kuang, J.; Hou, X.; Feng, Y.; Su, Z. Genetic variations in ADIPOQ gene are associated with chronic obstructive pulmonary disease. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Kearns, C.F.; Mckeever, K.H.; Roegner, V.; Brady, S.M.; Malinowski, K. Adiponectin and leptin are related to fat mass in horses. Vet. J. 2006, 172, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Du, Y.; Zhang, C.; Shi, X.; Chen, D.; Sun, J.; Jin, Q.; Lan, X.; Chen, H. Polymorphism in a microsatellite of the acrp30 gene and its association with growth traits in goats. Biochem. Genet. 2011, 49, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.H.; Xiong, Y.Z.; Deng, C.Y.; Jiang, S.W.; Zuo, B.; Zheng, R.; Li, F.E.; Lei, M.G. Association of the A-G polymorphism in porcine adiponectin gene with fat deposition and carcass traits. Asian-Aust. J. Anim. Sci. 2006, 19, 779–783. [Google Scholar] [CrossRef]
- Houde, A.A.; Murphy, B.D.; Mathieu, O.; Bordignon, V.; Palin, M.F. Characterization of swine adiponectin and adiponectin receptor polymorphisms and their association with reproductive traits. Anim. Genet. 2008, 39, 249–257. [Google Scholar] [CrossRef] [PubMed]
- An, Q.M.; Zhou, H.T.; Hu, J.; Luo, Y.Z.; Hickford, J.G. Haplotypes and Sequence Variation in the Ovine Adiponectin Gene (ADIPOQ). Genes 2015, 6, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.T.; Hickford, J.G.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.T.; Hickford, J.G. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Volk, M.G. An examination of the evidence supporting the association of dietary cholesterol and saturated fats with serum cholesterol and development of coronary heart disease. Altern. Med. Rev. 2007, 12, 228–245. [Google Scholar] [PubMed]
- Gibbs, R.A.; Belmont, J.W.; Hardenbol, P.; Willis, T.D.; Yu, F.; Yang, H.; Chang, L.Y.; Huang, W.; Liu, B.; Shen, Y.; et al. The International HapMap Project. Nature 2003, 426, 789–796. [Google Scholar]
- Snyder, M.W.; Adey, A.; Kitzman, J.O.; Shendure, J. Haplotype-resolved genome sequencing: Experimental methods and applications. Nat. Rev. Genet. 2015, 16, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Barth, N.; Langmann, T.; Schölmerich, J.; Schmitz, G.; Schäffler, A. Identification of regulatory elements in the human adipose most abundant gene transcript-1 (apM-1) promoter: Role of SP1/SP3 and TNF-α as regulatory pathways. Diabetologia 2004, 47, 153. [Google Scholar] [CrossRef]
- Menzaghi, C.; Ercolino, T.; Di, P.R.; Berg, A.H.; Warram, J.H.; Scherer, P.E.; Trischitta, V.; Doria, A. A haplotype at the adiponectin locus is associated with obesity and other features of the insulin resistance syndrome. Diabetes 2002, 51, 2306–2312. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.F.; Long, K.Z.; Hsu, C.C.; Mamun, A.A.; Chiu, Y.F.; Tu, H.P.; Chen, P.S.; Jhang, H.R.; Hwang, S.J.; Huang, M.C. Adiponectin gene (ADIPOQ) polymorphisms correlate with the progression of nephropathy in Taiwanese male patients with type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Chung, E. Novel SNPs in the bovine ADIPOQ and PPARGC1A genes are associated with carcass traits in Hanwoo (Korean cattle). Mol. Biol. Rep. 2013, 40, 4651–4660. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, J.; Flisikowska, T.; Schnieke, A.; Kind, A.; Szydlowski, M.; Switonski, M.; Flisikowski, K. Polymorphisms in the promoter region of the adiponectin (ADIPOQ) gene are presumably associated with transcription level and carcass traits in pigs. Anim. Genet. 2012, 44, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Owecki, M.; Miczke, A.; Kaczmarek, M.; Hoppe-Gołebiewska, J.; Pupek-Musialik, D.; Słomski, R.; Bryll, W.; Cymerys, M.; Nikisch, E.; Sowiński, J. The Y111 H (T415C) polymorphism in exon 3 of the gene encoding adiponectin is uncommon in Polish obese patients. Horm. Metab. Res. 2007, 39, 797–800. [Google Scholar] [CrossRef] [PubMed]
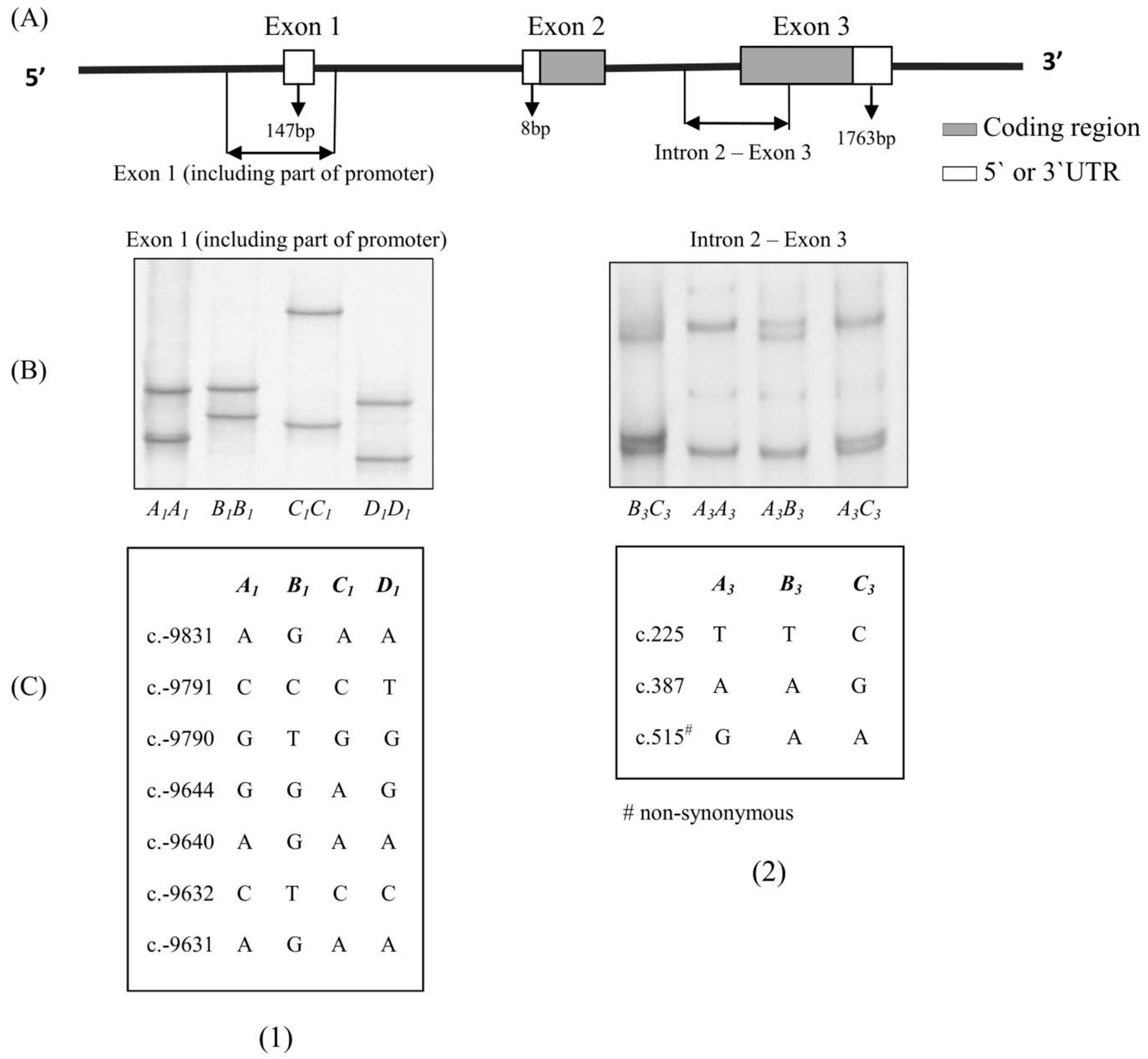

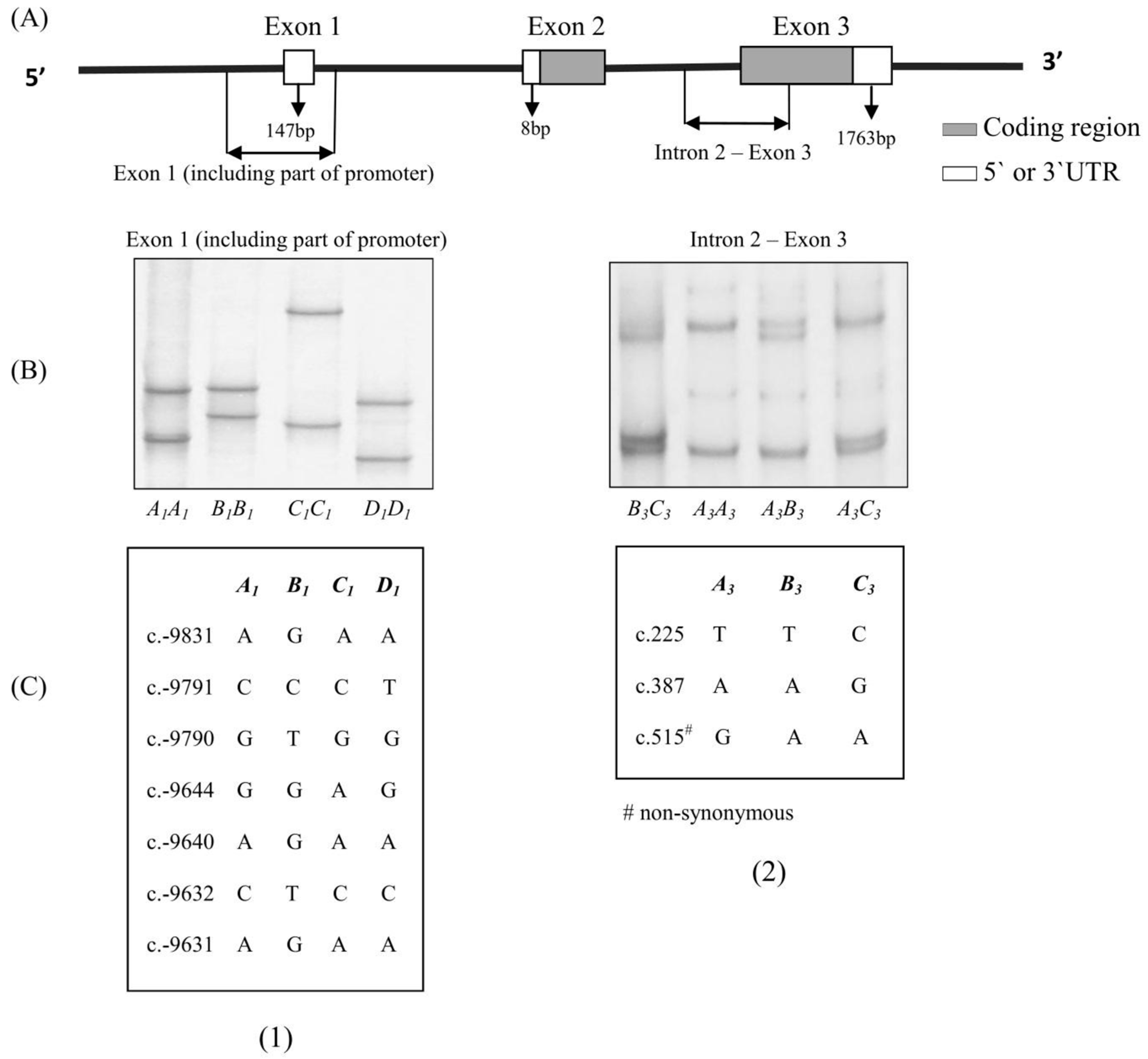{kind=link}
| Traits (unit) | Haplotype Assessed | Mean ± SE a | p-Value | |||
|---|---|---|---|---|---|---|
| Haplotype Absent (0) | n | Haplotype Present (1) | n | |||
| Birth weight (kg) | A1–A3 | 5.28 ± 0.15 | 384 | 5.30 ± 0.15 | 561 | 0.721 |
| A1–C3 | 5.31 ± 0.15 | 769 | 5.22 ± 0.16 | 176 | 0.248 | |
| B1–A3 | 5.24 ± 0.15 | 284 | 5.32 ± 0.15 | 661 | 0.203 | |
| B1–C3 | 5.30 ± 0.15 | 873 | 5.20 ± 0.17 | 72 | 0.264 | |
| Tailing weight (kg) | A1–A3 | 12.22 ± 0.25 | 384 | 12.13 ± 0.24 | 561 | 0.586 |
| A1–C3 | 12.19 ± 0.23 | 769 | 12.02 ± 0.29 | 176 | 0.436 | |
| B1–A3 | 12.15 ± 0.26 | 284 | 12.17 ± 0.23 | 661 | 0.885 | |
| B1–C3 | 12.14 ± 0.23 | 873 | 12.57 ± 0.37 | 72 | 0.293 | |
| Weaning weight (kg) | A1–A3 | 27.91 ± 0.41 | 384 | 27.46 ± 0.39 | 561 | 0.161 |
| A1–C3 | 27.63 ± 0.38 | 769 | 27.69 ± 0.49 | 176 | 0.884 | |
| B1–A3 | 27.58 ± 0.43 | 284 | 27.68 ± 0.39 | 661 | 0.745 | |
| B1–C3 | 27.62 ± 0.38 | 873 | 28.05 ± 0.61 | 72 | 0.405 | |
| Pre-weaning growth rate (g/day) | A1–A3 | 275.9 ± 8.54 | 384 | 268.2 ± 8.56 | 561 | 0.025 |
| A1–C3 | 272.6 ± 8.46 | 769 | 270.5 ± 9.32 | 176 | 0.684 | |
| B1–A3 | 271.7 ± 8.73 | 284 | 272.4 ± 8.49 | 661 | 0.844 | |
| B1–C3 | 272.0 ± 8.41 | 873 | 274.8 ± 9.97 | 72 | 0.633 | |
| Traits (unit) | Haplotype Assessed | Other Haplotype in Model | Mean ± SE a | p-Value | |||
|---|---|---|---|---|---|---|---|
| Haplotype Absent (0) | n | Haplotype Present (1) | n | ||||
| V-GR (mm) b | A1–A3 | — | 3.18 ± 0.37 | 159 | 3.10 ± 0.35 | 244 | 0.771 |
| A1–C3 | — | 2.87 ± 0.33 | 330 | 4.24 ± 0.42 | 73 | 0.001 | |
| B1–A3 | — | 3.39 ± 0.37 | 127 | 2.98 ± 0.35 | 276 | 0.136 | |
| B1–C3 | — | 2.26 ± 0.47 | 369 | 2.56 ± 0.64 | 34 | 0.738 | |
| A1–C3 | B1–A3 | 2.92 ± 0.39 | 330 | 4.43 ± 0.49 | 73 | 0.001 | |
| B1–A3 | A1–C3 | 3.56 ± 0.42 | 127 | 3.79 ± 0.45 | 276 | 0.478 | |
| Leg lean-meat yield (%) c | A1–A3 | — | 20.75 ± 0.16 | 159 | 20.30 ± 0.15 | 244 | 0.001 |
| A1–C3 | — | 20.50 ± 0.15 | 330 | 20.39 ± 0.19 | 73 | 0.450 | |
| B1–A3 | — | 20.30 ± 0.17 | 127 | 20.59 ± 0.16 | 276 | 0.023 | |
| B1–C3 | — | 20.97 ± 0.21 | 369 | 20.85 ± 0.29 | 34 | 0.585 | |
| A1–A3 | B1–A3 | 20.65 ± 0.18 | 159 | 20.18 ± 0.19 | 244 | 0.001 | |
| B1–A3 | A1–A3 | 20.36 ± 0.19 | 127 | 20.47 ± 0.20 | 276 | 0.467 | |
| Loin lean-meat yield (%) c | A1–A3 | — | 14.12 ± 0.11 | 159 | 13.94 ± 0.11 | 244 | 0.028 |
| A1–C3 | — | 14.01 ± 0.11 | 330 | 14.04 ± 0.13 | 73 | 0.723 | |
| B1–A3 | — | 13.98 ± 0.12 | 127 | 14.03 ± 0.11 | 276 | 0.598 | |
| B1–C3 | — | 14.28 ± 0.15 | 369 | 14.34 ± 0.21 | 34 | 0.565 | |
| Shoulder lean-meat yield (%) c | A1–A3 | — | 16.54 ± 0.13 | 159 | 16.46 ± 0.13 | 244 | 0.399 |
| A1–C3 | — | 16.52 ± 0.12 | 330 | 16.36 ± 0.15 | 73 | 0.183 | |
| B1–A3 | — | 16.51 ± 0.14 | 127 | 16.48 ± 0.13 | 276 | 0.786 | |
| B1–C3 | — | 17.01 ± 0.17 | 369 | 17.07 ± 0.23 | 34 | 0.808 | |
| Total lean-meat yield (%) d | A1–A3 | — | 51.42 ± 0.34 | 159 | 50.71 ± 0.32 | 244 | 0.004 |
| A1–C3 | — | 51.03 ± 0.32 | 330 | 50.79 ± 0.40 | 73 | 0.448 | |
| B1–A3 | — | 50.80 ± 0.35 | 127 | 51.10 ± 0.33 | 276 | 0.245 | |
| B1–C3 | — | 52.16 ± 0.44 | 369 | 52.26 ± 0.59 | 34 | 0.583 | |
| Proportion leg lean-meat yield (%) e | A1–A3 | — | 40.36 ± 0.16 | 159 | 40.04 ± 0.14 | 244 | 0.004 |
| A1–C3 | — | 40.17 ± 0.15 | 330 | 40.14 ± 0.18 | 73 | 0.806 | |
| B1–A3 | — | 39.96 ± 0.16 | 127 | 40.29 ± 0.15 | 276 | 0.006 | |
| B1–C3 | — | 40.00 ± 0.21 | 369 | 39.89 ± 0.29 | 34 | 0.838 | |
| A1–A3 | B1–A3 | 40.27 ± 0.17 | 159 | 40.00 ± 0.19 | 244 | 0.038 | |
| B1–A3 | A1–A3 | 39.99 ± 0.18 | 127 | 40.28 ± 0.19 | 276 | 0.046 | |
| Proportion loin lean-meat yield (%) e | A1–A3 | — | 27.47 ± 0.13 | 159 | 27.50 ± 0.12 | 244 | 0.746 |
| A1–C3 | — | 27.45 ± 0.12 | 330 | 27.75 ± 0.15 | 73 | 0.048 | |
| B1–A3 | — | 27.53 ± 0.13 | 127 | 27.46 ± 0.12 | 276 | 0.458 | |
| B1–C3 | — | 27.37 ± 0.17 | 369 | 27.43 ± 0.23 | 34 | 0.836 | |
| Proportion shoulder lean-meat yield (%) e | A1–A3 | — | 32.17 ± 0.16 | 159 | 32.46 ± 0.15 | 244 | 0.011 |
| A1–C3 | — | 32.38 ± 0.15 | 330 | 32.21 ± 0.19 | 73 | 0.257 | |
| B1–A3 | — | 32.51 ± 0.16 | 127 | 32.25 ± 0.15 | 276 | 0.037 | |
| B1–C3 | — | 32.63 ± 0.22 | 369 | 32.68 ± 0.29 | 34 | 0.713 | |
| A1–A3 | B1–A3 | 32.18 ± 0.18 | 159 | 32.37 ± 0.19 | 244 | 0.147 | |
| B1–A3 | A1–A3 | 32.45 ± 0.18 | 127 | 32.11 ± 0.19 | 276 | 0.024 | |
| Traits (unit) | Mean ± SE a | p-Value | ||||
|---|---|---|---|---|---|---|
| A1–A3/A1–A3 (n = 61) | A1–A3/A1–C3 (n = 38) | A1–A3/B1–A3 (n = 140) | B1–A3/B1–A3 (n = 89) | B1–A3/B1–C3 (n = 24) | ||
| V-GR (mm) b | 1.65 ± 0.68 b | 3.22 ± 0.74 a | 2.01 ± 0.64 a,b | 1.94 ± 0.63 a,b | 2.25 ± 0.77 a,b | 0.062 |
| Leg lean-meat yield (%) c | 20.80 ± 0.31 b | 20.86 ± 0.34 a,b | 20.96 ± 0.29 b | 21.41 ± 0.29 a | 21.11 ± 0.35 a,b | 0.017 |
| Loin lean-meat yield (%) c | 14.19 ± 0.22 b | 14.54 ± 0.25 a,b | 14.30 ± 0.21 a,b | 14.59 ± 0.21 a | 14.40 ± 0.26 a,b | 0.028 |
| Shoulder lean-meat yield (%) c | 17.21 ± 0.25 | 17.07 ± 0.28 | 17.03 ± 0.24 | 17.25 ± 0.23 | 17.22 ± 0.29 | 0.347 |
| Total lean-meat yield (%) d | 52.20 ± 0.65 a,b | 52.47 ± 0.71 a,b | 52.29 ± 0.61 b | 53.24 ± 0.60 a | 52.73 ± 0.74 a,b | 0.037 |
| Proportion leg lean-meat yield (%) e | 39.82 ± 0.31 | 39.73 ± 0.34 | 40.07 ± 0.29 | 40.19 ± 0.28 | 40.02 ± 0.35 | 0.227 |
| Proportion loin lean-meat yield (%) e | 27.18 ± 0.25 b | 27.73 ± 0.27 a | 27.35 ± 0.23 ab | 27.39 ± 0.23 a,b | 27.31 ± 0.28 a,b | 0.094 |
| Proportion shoulder lean-meat yield (%) e | 33.00 ± 0.32 a | 32.54 ± 0.35 a,b | 32.58 ± 0.30 a,b | 32.42 ± 0.29 b | 32.67 ± 0.36 a,b | 0.061 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Q.; Zhou, H.; Hu, J.; Luo, Y.; Hickford, J.G.H. Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs. Genes 2017, 8, 160. https://doi.org/10.3390/genes8060160
An Q, Zhou H, Hu J, Luo Y, Hickford JGH. Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs. Genes. 2017; 8(6):160. https://doi.org/10.3390/genes8060160
Chicago/Turabian StyleAn, Qingming, Huitong Zhou, Jiang Hu, Yuzhu Luo, and Jon G. H. Hickford. 2017. "Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs" Genes 8, no. 6: 160. https://doi.org/10.3390/genes8060160
APA StyleAn, Q., Zhou, H., Hu, J., Luo, Y., & Hickford, J. G. H. (2017). Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs. Genes, 8(6), 160. https://doi.org/10.3390/genes8060160