Effects of Cellular Methylation on Transgene Expression and Site-Specific Integration of Adeno-Associated Virus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Reagents
2.2. AAV Production and Purification
2.3. Polymerase Chain Reaction(PCR) Analysis
2.4. Southern Hybridization
2.5. Biotinylation and Streptavidin Labeling of rAAV-2
3. Results
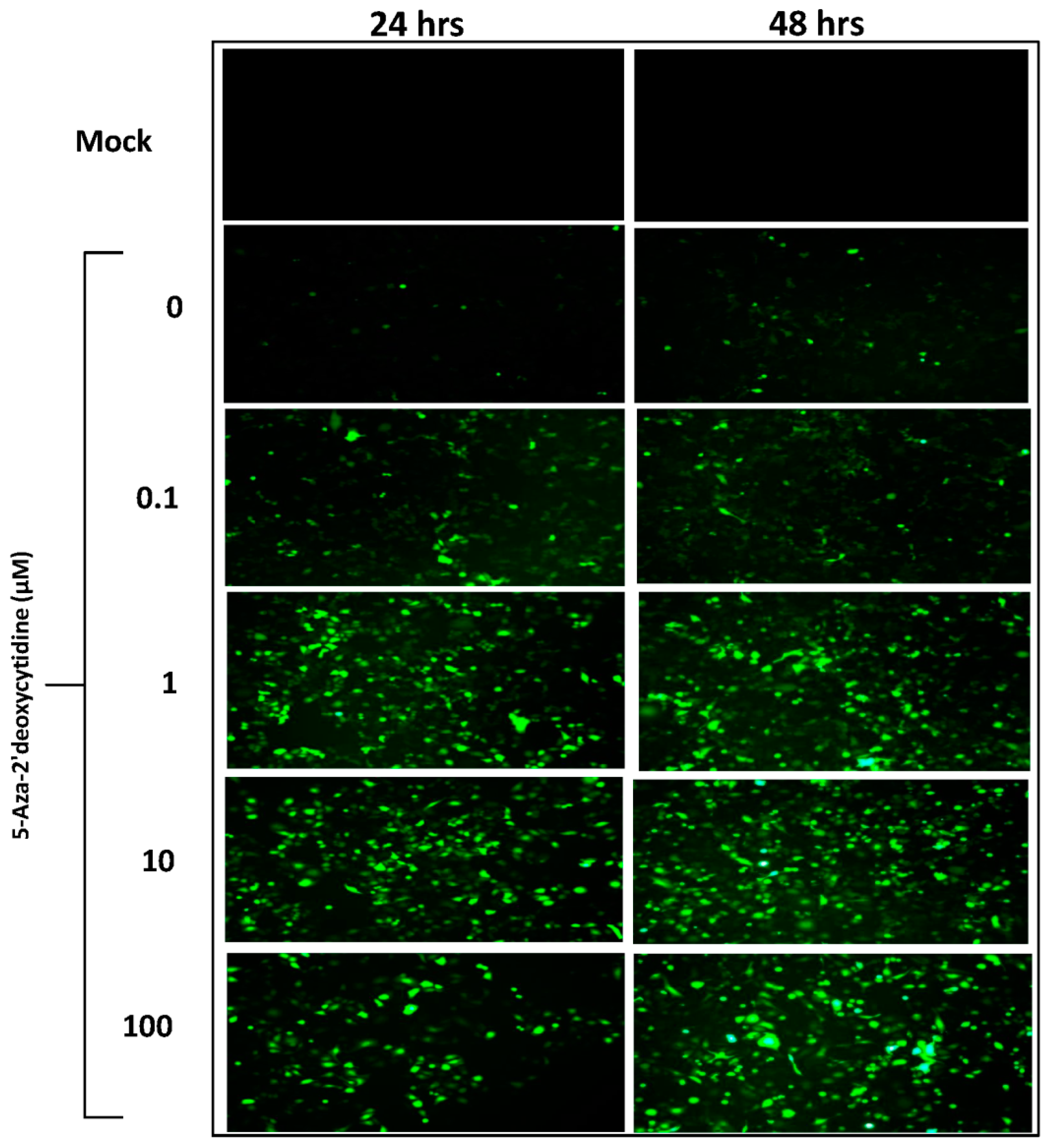
3.1. Effect of Hypo-Methylation on AAV-Transgene Expression
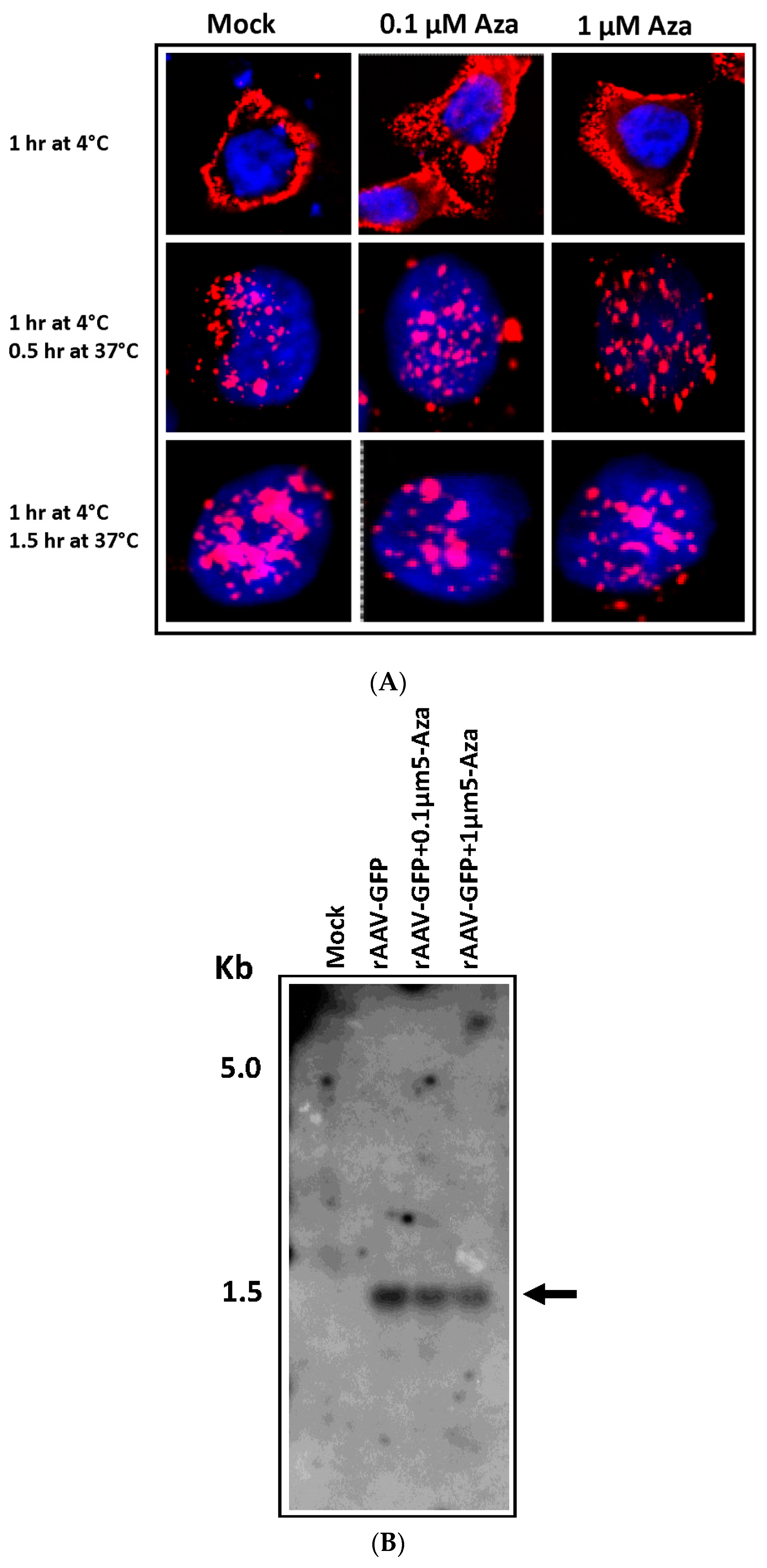
3.2. Enhanced GFP Expression in rAAV-GFP Infected Hypo-Methylated HeLa Cells Is Not Due to Enhanced Viral Transduction Following Aza Treatment
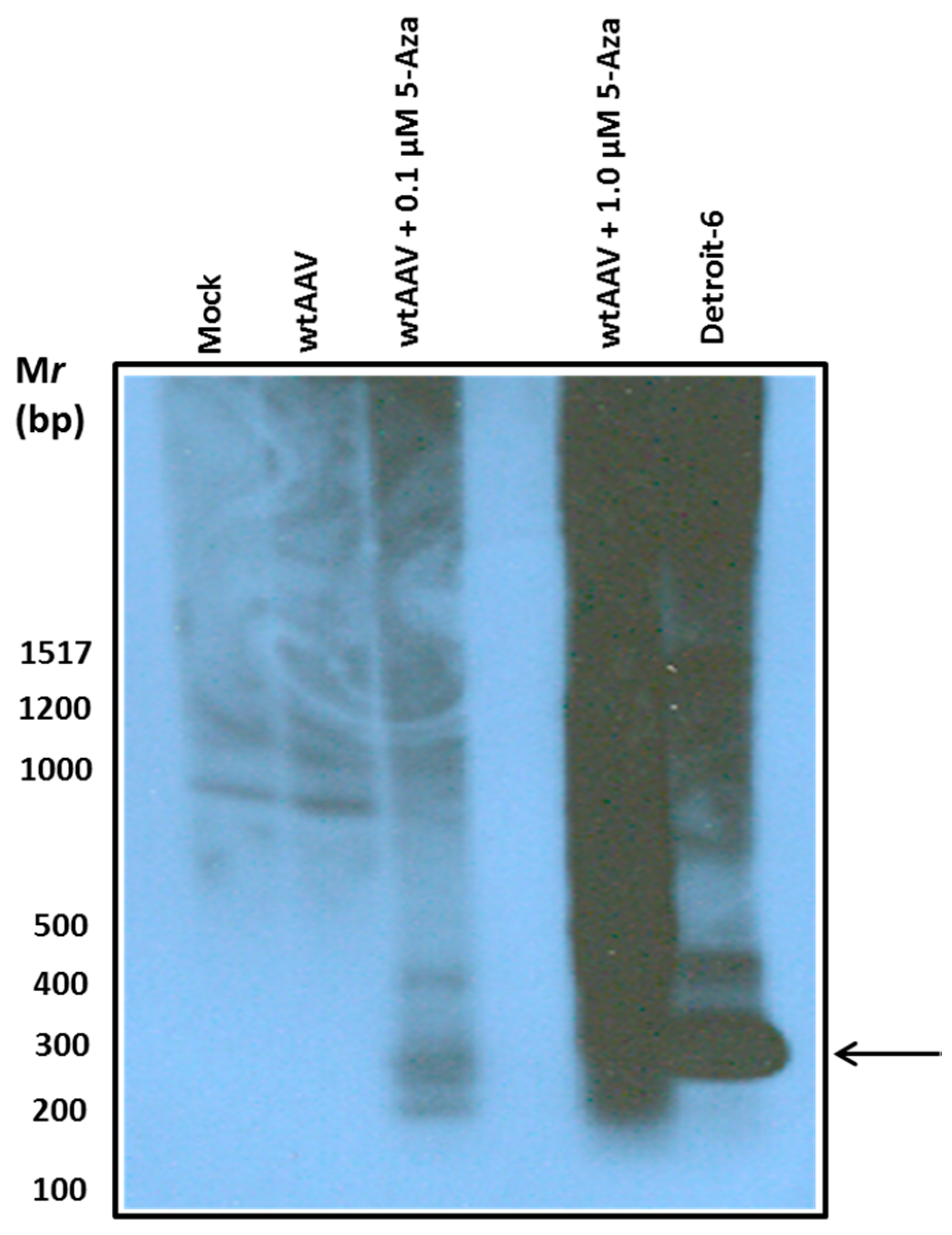
3.3. Effect of Hypo-Methylation on Site-Specific Integration of wtAAV
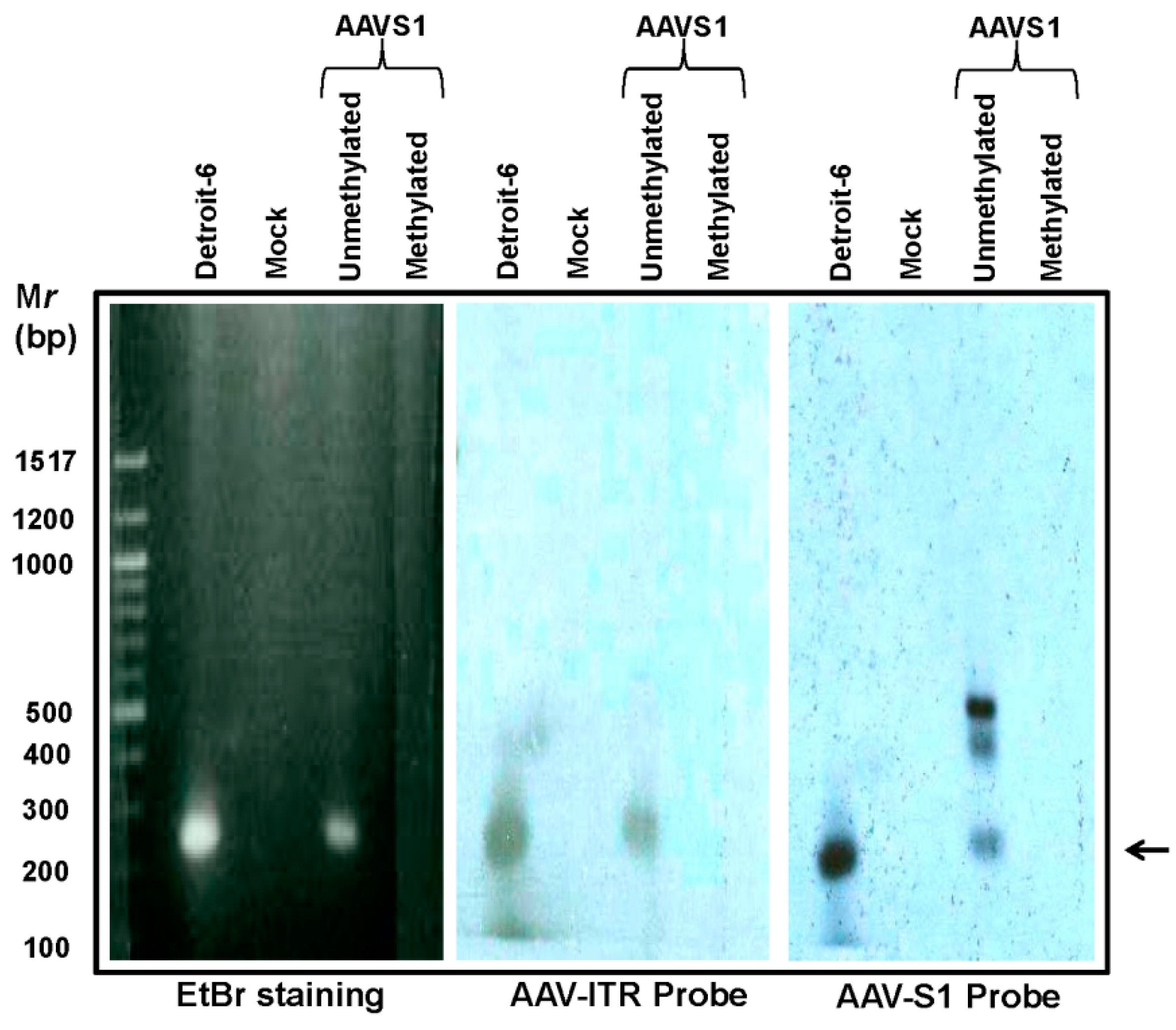
3.4. Hypo-Methylation Increases Site-Specific Integration of wtAAV Affecting AAVS1
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Doerfler, W. Epigenetic consequences of foreign DNA insertions: De novo methylation and global alterations of methylation patterns in recipient genomes. Rev. Med. Virol. 2011, 21, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.H.; Zolotukhin, S.; Muzyczka, N. Sequence requirements for binding of rep68 to the adeno-associated virus terminal repeats. J. Virol. 1996, 70, 1542–1553. [Google Scholar] [PubMed]
- Krieg, A.M. Direct immunologic activities of CpG DNA and implications for gene therapy. J. Gene Med. 1999, 1, 56–63. [Google Scholar] [CrossRef]
- Bauer, S.; Kirschning, C.J.; Hacker, H.; Redecke, V.; Hausmann, S.; Akira, S.; Wagner, H.; Lipford, G.B. Human Tlr9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci. USA 2001, 98, 9237–9242. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.D.; Smith, A.J.; De Gobbi, M.; Flenley, M.; Hughes, J.R.; Vernimmen, D.; Ayyub, H.; Sharpe, J.A.; Sloane-Stanley, J.A.; Sutherland, L.; et al. An interspecies analysis reveals a key role for unmethylated CpG dinucleotides in vertebrate polycomb complex recruitment. EMBO J. 2012, 31, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Pannell, D.; Ellis, J. Silencing of gene expression: Implications for design of retrovirus vectors. Rev. Med. Virol. 2001, 11, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.R.; Harkins, R.N.; Wang, P.; Qian, H.S.; Liu, P.; Rubanyi, G.M. Transcriptional silencing is associated with extensive methylation of the CMV promoter following adenoviral gene delivery to muscle. J. Gene Med. 2004, 6, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Schnieke, A.; Harbers, K. Treatment of mice with 5-azacytidine efficiently activates silent retroviral genomes in different tissues. Proc. Natl. Acad. Sci. USA 1985, 82, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Ortolano, S.; Spuch, C.; Navarro, C. Present and future of adeno associated virus based gene therapy approaches. Recent Pat. Endocr. Metab. Immune Drug Discov. 2012, 6, 47–66. [Google Scholar] [CrossRef] [PubMed]
- Hermonat, P.L. Adeno-associated virus inhibits human papillomavirus type 16: A viral interaction implicated in cervical cancer. Cancer Res. 1994, 54, 2278–2281. [Google Scholar] [PubMed]
- Li, L.C.; Dahiya, R. Methprimer: Designing primers for methylation pcrs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Linden, R.M.; Ward, P.; Giraud, C.; Winocour, E.; Berns, K.I. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA 1996, 93, 11288–11294. [Google Scholar] [CrossRef] [PubMed]
- Giraud, C.; Winocour, E.; Berns, K.I. Site-specific integration by adeno-associated virus is directed by a cellular DNA sequence. Proc. Natl. Acad. Sci. USA 1994, 91, 10039–10043. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- McCarty, D.M. Self-complementary AAV vectors; advances and applications. Mol. Ther. 2008, 16, 1648–1656. [Google Scholar] [CrossRef] [PubMed]
- Finn, J.D.; Hui, D.; Downey, H.D.; Dunn, D.; Pien, G.C.; Mingozzi, F.; Zhou, S.; High, K.A. Proteasome inhibitors decrease AAV2 capsid derived peptide epitope presentation on MHC class I following transduction. Mol. Ther. 2010, 18, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Mingozzi, F.; High, K.A. Immune responses to AAV in clinical trials. Curr. Gene Ther. 2011, 11, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Zolotukhin, S.; Byrne, B.J.; Mason, E.; Zolotukhin, I.; Potter, M.; Chesnut, K.; Summerford, C.; Samulski, R.J.; Muzyczka, N. Recombinant adeno-associated virus purification using novel methods improves infectious titer and yield. Gene Ther. 1999, 6, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Lamartina, S.; Roscilli, G.; Rinaudo, D.; Delmastro, P.; Toniatti, C. Lipofection of purified adeno-associated virus rep68 protein: Toward a chromosome-targeting nonviral particle. J. Virol. 1998, 72, 7653–7658. [Google Scholar] [PubMed]
- Ren, C.; Kumar, S.; Shaw, D.R.; Ponnazhagan, S. Genomic stability of self-complementary adeno-associated virus 2 during early stages of transduction in mouse muscle in vivo. Hum. Gene Ther. 2005, 16, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Ponnazhagan, S.; Mahendra, G.; Kumar, S.; Thompson, J.A.; Castillas, M., Jr. Conjugate-based targeting of recombinant adeno-associated virus type 2 vectors by using avidin-linked ligands. J. Virol. 2002, 76, 12900–12907. [Google Scholar] [CrossRef] [PubMed]
- Christman, J.K. 5-azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [PubMed]
- Hirt, B. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 1967, 26, 365–369. [Google Scholar] [CrossRef]
- Karlin, S.; Doerfler, W.; Cardon, L.R. Why is CpG suppressed in the genomes of virtually all small eukaryotic viruses but not in those of large eukaryotic viruses? J. Virol. 1994, 68, 2889–2897. [Google Scholar] [PubMed]
- Schlehofer, J.R.; Ehrbar, M.; zur Hausen, H. Vaccinia virus, herpes simplex virus, and carcinogens induce DNA amplification in a human cell line and support replication of a helpervirus dependent parvovirus. Virology 1986, 152, 110–117. [Google Scholar] [CrossRef]
- Ferrari, F.K.; Samulski, T.; Shenk, T.; Samulski, R.J. Second-strand synthesis is a rate-limiting step for efficient transduction by recombinant adeno-associated virus vectors. J. Virol. 1996, 70, 3227–3234. [Google Scholar] [PubMed]
- McCarty, D.M.; Young, S.M., Jr.; Samulski, R.J. Integration of adeno-associated virus (AAV) and recombinant AAV vectors. Ann. Rev. Genet. 2004, 38, 819–845. [Google Scholar] [CrossRef] [PubMed]
- Luszczek, W.; Cheriyath, V.; Mekhail, T.M.; Borden, E.C. Combinations of DNA methyltransferase and histone deacetylase inhibitors induce DNA damage in small cell lung cancer cells: Correlation of resistance with IFN-stimulated gene expression. Mol. Cancer Ther. 2010, 9, 2309–2321. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chanda, D.; Hensel, J.A.; Higgs, J.T.; Grover, R.; Kaza, N.; Ponnazhagan, S. Effects of Cellular Methylation on Transgene Expression and Site-Specific Integration of Adeno-Associated Virus. Genes 2017, 8, 232. https://doi.org/10.3390/genes8090232
Chanda D, Hensel JA, Higgs JT, Grover R, Kaza N, Ponnazhagan S. Effects of Cellular Methylation on Transgene Expression and Site-Specific Integration of Adeno-Associated Virus. Genes. 2017; 8(9):232. https://doi.org/10.3390/genes8090232
Chicago/Turabian StyleChanda, Diptiman, Jonathan A. Hensel, Jerome T. Higgs, Rajat Grover, Niroop Kaza, and Selvarangan Ponnazhagan. 2017. "Effects of Cellular Methylation on Transgene Expression and Site-Specific Integration of Adeno-Associated Virus" Genes 8, no. 9: 232. https://doi.org/10.3390/genes8090232
APA StyleChanda, D., Hensel, J. A., Higgs, J. T., Grover, R., Kaza, N., & Ponnazhagan, S. (2017). Effects of Cellular Methylation on Transgene Expression and Site-Specific Integration of Adeno-Associated Virus. Genes, 8(9), 232. https://doi.org/10.3390/genes8090232