Association of Candidate Genes with Response to Heat and Newcastle Disease Virus

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
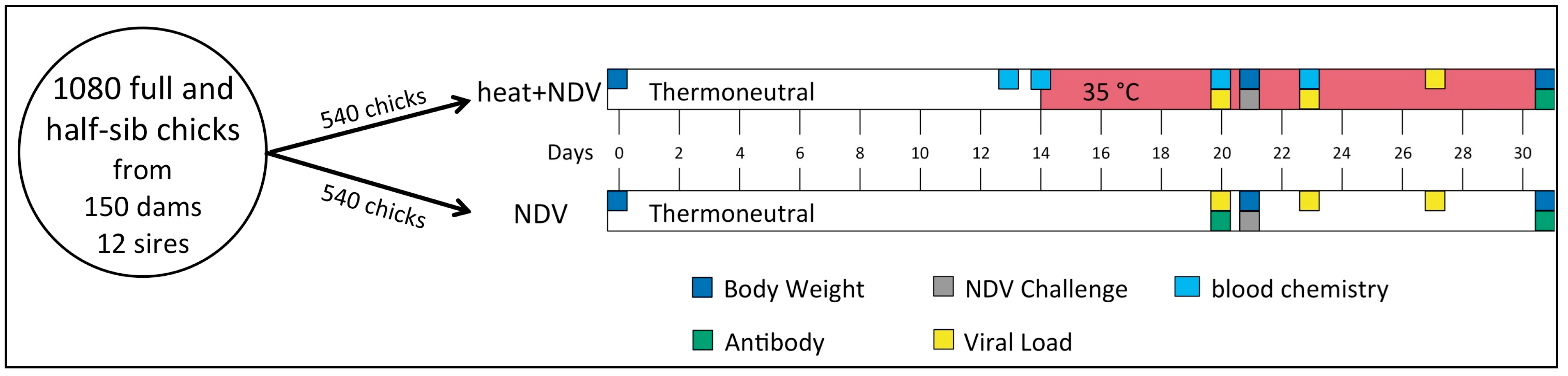
2.1. Animals and Husbandry
2.2. Experimental Design
2.3. Viral Load
2.4. Antibody
2.5. Growth Rate
2.6. Heat-Related Phenotypes
2.7. Selection of Genes and Genotyping
2.8. Data Analysis
3. Results and Discussion
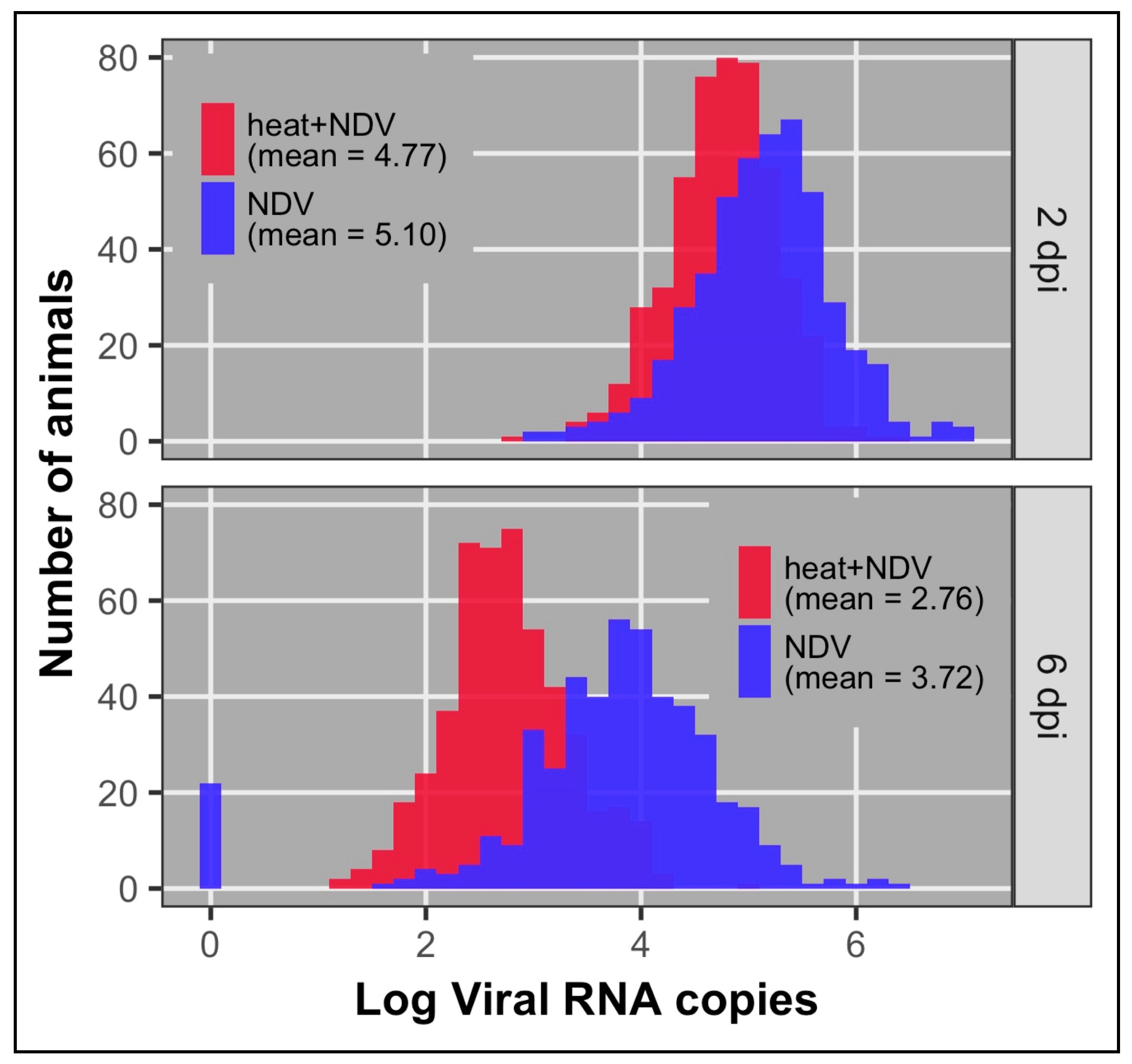
3.1. Viral Load
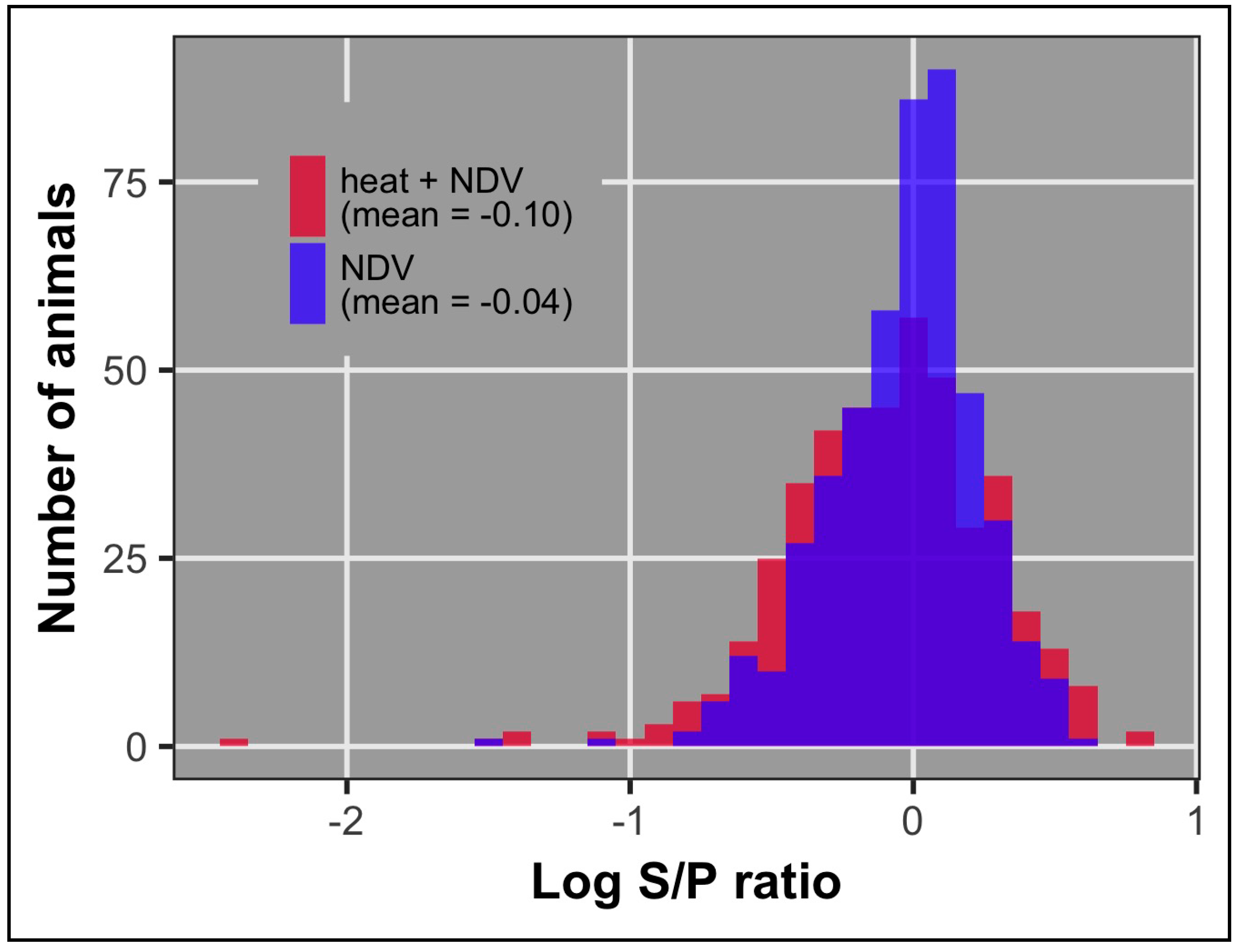
3.2. Antibody
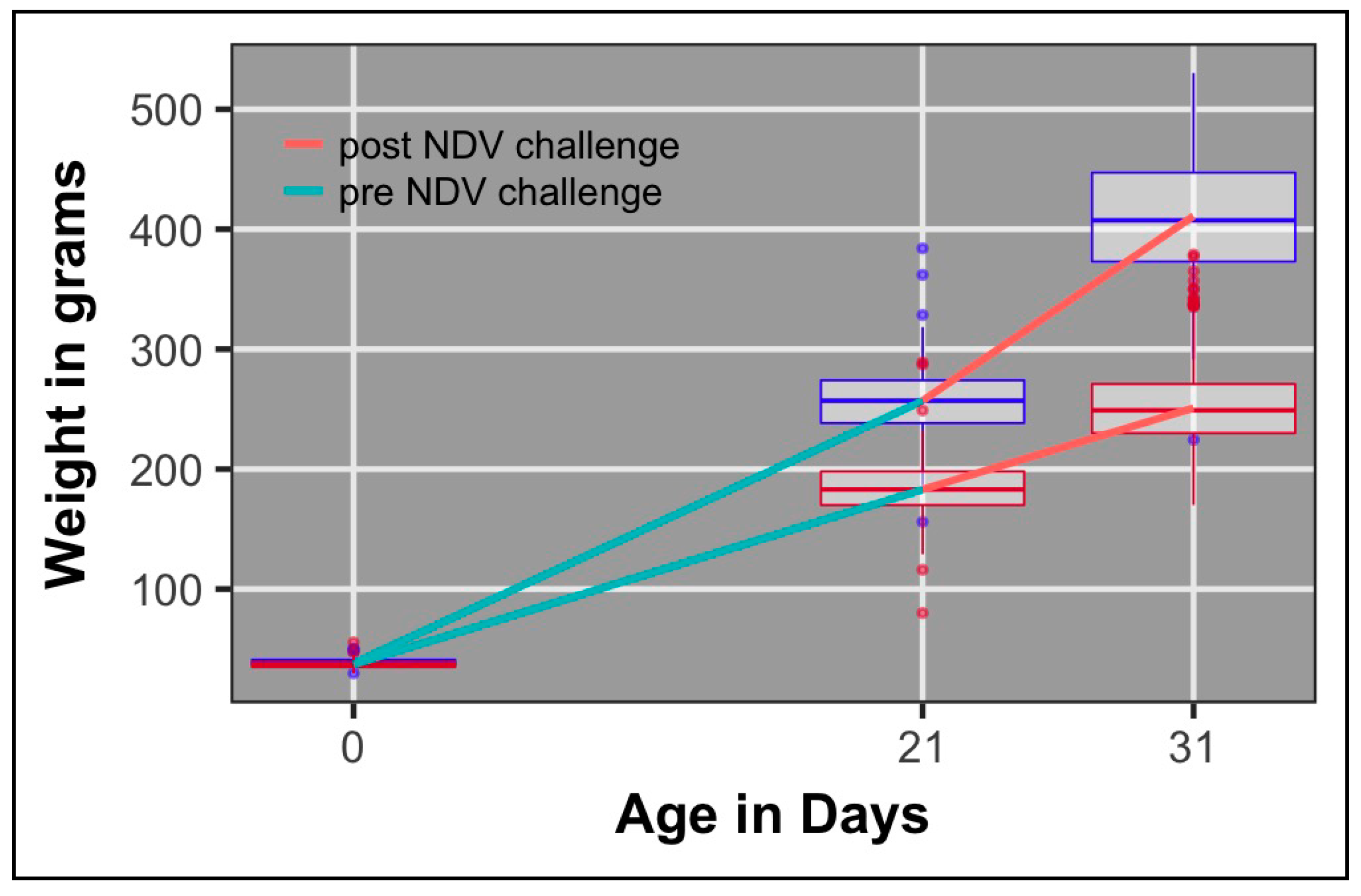
3.3. Growth Rate
3.4. Heat-Related Phenotypes
3.5. Haplotype Effects
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kitalyi, A.J. Village Chicken Production Systems in Rural Africa: Household Food Security and Gender Issues; FAO: Rome, Italy, 1998. [Google Scholar]
- Nyoni, N.M.B.; Grab, S.; Archer, E.R.M. Heat stress and chickens: Climate risk effects on rural poultry farming in low-income countries. Clim. Dev. 2018. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Havenstein, G.B.; Ferket, P.R.; Qureshi, M.A. Growth, livability, and feed conversion of 1957 vs. 1991 broilers when fed “typical” 1957 and 1991 broiler diets. Poult. Sci. 1994, 73, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.K.; Hutt, F.B. Genetic differences in resistance to Newcastle disease. Avian Dis. 1961, 5, 205–214. [Google Scholar] [CrossRef]
- Gordon, C.D.; Beard, C.W.; Hopkins, S.R.; Siegel, H.S. Chick mortality as a criterion for selection toward resistance or susceptibility to Newcastle disease. Poult. Sci. 1970, 50, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Peleg, B.A.; Soller, M.; Ron, N.; Hornstein, K.; Brody, T.; Kalmar, E. Familial differences in antibody response of broiler chickens to vaccination with attenuated and inactivated Newcastle disease virus vaccine. Avian Dis. 1976, 20, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Pitcovski, J.; Heller, D.E.; Cahaner, A.; Peleg, B.A. Selection for early responsiveness of chicks to Escherichia coli and Newcastle disease virus. Poult. Sci. 1987, 66, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Soller, M.; Heller, D.; Peleg, B.; Ron-Kuper, N.; Hornstein, K. Genetic and phenotypic correlations between immune response to Escherichia coli and to Newcastle disease virus vaccines. Poult. Sci. 1981, 60, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.R.; Wilcox, C.J.; Voitle, R.A.; Baird, C.D.; Dorminey, R.W. Characteristics of White Leghorn chickens selected for heat tolerance. Poult. Sci. 1975, 54, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Mack, L.A.; Felver-Gant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variations alter production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washburn, K.W.; Peavey, R.; Renwick, G.M. Relationship of strain variation and feed restriction to variation in blood pressure and response to heat stress. Poult. Sci. 1980, 59, 2586–2588. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Hu, J.; Hu, S.; Song, Q.; Ding, P.; Zhu, J.; Liu, X.; Wang, X.; Liu, X. High levels of virus replication and an intense inflammatory response contribute to the severe pathology in lymphoid tissues caused by Newcastle disease virus genotype VIId. Arch. Virol. 2015, 160, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Moeini, H.; Ideris, A.; Bejo, M.H.; Alitheen, N.B.M.; Kaiser, P.; Omar, A.R. Alteration in lymphocyte responses, cytokine and chemokine profiles in chickens infected with genotype VII and VIII velogenic Newcastle disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional innate immune response of the developing chicken embryo to Newcastle disease virus infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Cheng, C.Y.; Tang, P.C.; Chen, C.F.; Chen, H.H.; Lee, Y.P.; Huang, S.Y. Differential gene expressions in testes of L2 strain Taiwan country chicken in response to acute heat stress. Theriogenology 2013, 79, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.Q.; Wang, X.L.; Wang, X.W.; Yan, C.Q.; Lv, C.J.; Li, X.Q.; Chu, Z.L.; Adam, F.E.A.; Xiao, S.; Zhang, S.X.; et al. Common microRNA–mRNA interactions in different Newcastle disease virus-infected chicken embryonic visceral tissues. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, L.; Cui, D.; Ruan, W.; Liu, F.; Li, H. Differential expression of the Toll-like receptor pathway and related genes of chicken bursa after experimental infection with infectious bursa disease virus. Arch. Virol. 2012, 157, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczynski, D.R.; Suarez, D.L.; King, D.J.; Miller, P.J.; Afonso, C.L. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Feng, M.; Zhao, X.; Dai, X.; Xiang, B.; Gao, P.; Li, Y.; Li, Y.; Ren, T. Newcastle disease virus infection in chicken embryonic fibroblasts but not duck embryonic fibroblasts is associated with elevated host innate immune response. Virol. J. 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Sun, Y.; Zhang, X.; Zhang, F.; Zhang, S.; Yu, S.; Qiu, X.; Tan, L.; Song, C.; Gao, S.; et al. Toll-like receptor 3 inhibits Newcastle disease virus replication through activation of pro-inflammatory cytokines and the type-1 interferon pathway. Arch. Virol. 2014, 159, 2937–2948. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Li, W.; Huang, Y.; He, J.; Tian, Y. The effect of selenium and polysaccharide of Atractylodes macrocephala Koidz (PAMK) on immune response in chicken spleen under heat stress. Biol. Trace Elem. Res. 2014, 160, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Lamont, S.J.; Cooksey, A.M.; Mccarthy, F.; Tudor, C.O.; Vijay-Shanker, K.; Derita, R.M.; Rothschild, M.; Ashwell, C.; Persia, M.E.; et al. Transcriptome response to heat stress in a chicken hepatocellular carcinoma cell line. Cell Stress Chaperones 2015, 20, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reemers, S.S.; van Haarlem, D.A.; Groot Koerkramp, M.J.; Vervelde, L. Differential gene-expression and host-response profiles against avian influenza virus within the chicken lung due to anatomy and airflow. J. Gen. Virol. 2009, 90, 2134–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micheli, L.; Leonardi, L.; Conti, F.; Maresca, G.; Colazingari, S.; Mattei, E.; Lira, S.A.; Farioli-Vecchioli, S.; Caruso, M.; Tirone, F. PC4/Tis7/IFRD1 stimulates skeletal muscle regeneration and is involved in myoblast differentiation as a regulator of MyoD and NF-κB. J. Biol. Chem. 2011. [Google Scholar] [CrossRef] [PubMed]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel analysis of the Harderian gland transcriptome response to Newcastle disease virus in two inbred chicken lines. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscott, J.; Nguyen, T.L.A.; Arguello, M.; Nakhaei, P.; Paz, S. Manipulation of the nuclear factor-κB pathway and the innate immune response by viruses. Oncogene 2006, 25, 6844–6867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byland, R.; Vance, P.J.; Hoxie, J.A.; Marsh, M. A conserved dileucine motif mediates clathrin and AP-2-dependent endocytosis of the HIV-1 envelope protein. Mol. Biol. Cell 2007, 18, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Gytz, H.; Hansen, M.F.; Skovbjerg, S.; Kristensen, A.C.M.; Hørlyck, S.; Jensen, M.B.; Fredborg, M.; Markert, L.D.; McMillan, N.A.; Christensen, E.I.; et al. Apoptotic properties of the type 1 interferon induced family of human mitochondrial membrane ISG12 proteins. Biol. Cell 2017, 109, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Ganar, K.; Das, M.; Sinha, S.; Kumar, S. Newcastle disease virus: Current status and our understanding. Virus Res. 2014, 184, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Menoret, A.; Srivastava, P. Roles of heat-shock proteins in antigen presentation and cross-presentation. Curr. Opin. Immunol. 2002, 14, 45–51. [Google Scholar] [CrossRef]
- Kilic, I.; Simsek, E. The effects of heat stress on egg production and quality of laying hens. J. Anim. Vet. Adv. 2013, 12, 42–47. [Google Scholar]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamonta, S.J. Novel mechanisms revealed in the trachea transcriptome of resistant and susceptible chicken lines following infection with Newcastle disease virus. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef] [PubMed]
- Rowland, K.; Wolc, A.; Gallardo, R.A.; Kelly, T.; Zhou, H.; Dekkers, J.C.M.; Lamont, S.J. Genetic analysis of a commercial egg laying line challenged with Newcastle disease virus. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Saelao, P.; Wang, Y.; Chanthavixay, G.; Gallardo, R.A.; Wolc, A.; Dekkers, J.C.M.; Lamont, S.J.; Kelly, T.; Zhou, H. Genomic regions affecting response to Newcastle disease virus infection in heat stressed Hy-Line Brown layers using chicken 600 K SNP array. BMC Genet. 2018. submitted. [Google Scholar]
- Fulton, J.E.; McCarron, A.M.; Lund, A.R.; Pinegar, K.N.; Wolc, A.; Chazara, O.; Bed’Hom, B.; Berres, M.; Miller, M.M. A high-density SNP panel reveals extensive diversity, frequent recombination and multiple recombination hotspots within the chicken major histocompatibility complex B region between BG2 and CD1A1. Genet. Sel. Evol. 2016, 48, 1. [Google Scholar] [CrossRef] [PubMed]
- Kranis, A.; Gheyas, A.A.; Boschiero, C.; Turner, F.; Yu, L.; Smith, S.; Talbot, R.; Pirani, A.; Brew, F.; Kaiser, P.; et al. Development of a high density 600K SNP genotyping array for chicken. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33. [Google Scholar] [CrossRef]
- Wang, Y.; Saelao, P.; Rowland, K.; Chanthavixay, G.; Gallardo, R.A.; Wolc, A.; Dekkers, J.M.; Lamont, S.J.; Kelly, T.; Zhou, H. Genomic regions affecting response to heat stress in Hy-Line Brown layers using chicken 600 K SNP array. Unpublished word. 2018. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Welham, S.J.; Thompson, R. ASReml User Guide Release 4.1 Functional Specification; VSN International Ltd.: Hemel Hempstead, UK, 2015. [Google Scholar]
- VanRaden, P.M. Efficient methods to compute genomic predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Lwelamira, J.; Kifaro, G.C.; Gwakisa, P.S. Genetic parameters for body weights, egg traits and antibody response against Newcastle Disease Virus (NDV) vaccine among two Tanzania chicken ecotypes. Trop. Anim. Health Prod. 2009, 41, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Van Goor, A.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Quantitative trait loci identified for blood chemistry components of an advanced intercross line of chickens under heat stress. BMC Genom. 2016, 17, 287. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Wang, Y.; Tian, K.; Ye, F.; Yin, H.; Zhao, X.; Xu, H.; Huang, Y.; Liu, H.; Hsieh, J.C.F.; et al. Integrated host and viral transcriptome analyses reveal pathology and inflammatory response mechanisms to ALV-J injection in SPF chickens. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Cong, F.; Liu, X.; Han, Z.; Shao, Y.; Kong, X.; Liu, S. Transcriptome analysis of chicken kidney tissues following coronavirus avian infectious bronchitis virus infection. BMC Genom. 2013. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Bao, Y.; Ye, J.; Shao, H.; Qian, K.; Qin, A. Transcriptional profiling of host gene expression in chicken embryo fibroblasts infected with reticuloendotheliosis virus strain HA1101. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, L.; Zhang, Y.; Xue, C.; Cao, Y. A novel method for genome-wide profiling of dynamic host-pathogen interactions using 3′ end enriched RNA-seq. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panayotova-Heiermann, M.; Loo, D.D.F.; Lostao, M.P.; Wright, E.M. Sodium/D-glucose cotransporter charge movements involve polar residues. J. Biol. Chem. 1994, 269, 21016–21020. [Google Scholar] [PubMed]
- Garriga, C.; Hunter, R.R.; Amat, C.; Planas, J.M.; Mitchell, M.A.; Moretó, M. Heat stress increases apical glucose transport in the chicken jejunum. Am. J. Physiol. Integr. Comp. Physiol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, M.; Cota, M.; Arce, N.; Castillo, G.; Avelar, E.; Espinoza, S.; Morales, A. Effect of heat stress on performance and expression of selected amino acid and glucose transporters, HSP90, leptin and ghrelin in growing pigs. J. Therm. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Iezaki, T.; Onishi, Y.; Ozaki, K.; Fukasawa, K.; Takahata, Y.; Nakamura, Y.; Fujikawa, K.; Takarada, T.; Yoneda, Y.; Yamashita, Y.; et al. The transcriptional modulator interferon-related developmental regulator 1 in osteoblasts suppresses bone formation and promotes bone resorption. J. Bone Miner. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- ChickenQTLdb. Available online: https://www.animalgenome.org/cgi-bin/gbrowse/gallus/ (accessed on 15 June 2018).
- Xie, J.; Tang, L.; Lu, L.; Zhang, L.; Xi, L.; Liu, H.C.; Odle, J.; Luo, X. Differential expression of heat shock transcription factors and heat shock proteins after acute and chronic heat stress in laying chickens (Gallus gallus). PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Takemoto, S.; Takakura, Y. Heat shock protein derivatives for delivery of antigens to antigen presenting cells. Int. J. Pharm. 2008, 354, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kaiser, M.G.; Deist, M.S.; Gallardo, R.A.; David, A.B.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Transcriptome analysis in spleen reveals differential regulation of response to Newcastle disease virus in two chicken lines. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Resistant and susceptible chicken lines show distinctive responses to Newcastle disease virus infection in the lung transcriptome. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Number of Haplotypes | Base Pairs 1 | Number of SNPs |
|---|---|---|---|---|
| TLR3 | Toll like receptor 3 | 3 | 4819 | 21 |
| TLR7 | Toll like receptor 7 | 3 | 1227 | 5 |
| MX | MX dynamin like GTPase 1 | 5 | 20,492 | 34 |
| MHC-B | Major histocompatibility complex B region 2 | 9 | 210,744 | 90 |
| IFI27L2 | Interferon alpha inducible protein 27 like 1 | 4 | 446 | 7 |
| SLC5A1 | Solute carrier family 5 member 1 | 5 | 9763 | 8 |
| HSPB1 | Heat shock protein family B (small) member 1 | 2 | 1739 | 5 |
| HSPA2 | Heat shock 70 kDa protein 2 | 4 | 1364 | 5 |
| HSPA8 | Heat shock 70 kDa protein 8 | 7 | 5747 | 6 |
| IFRD1 | Interferon related developmental regulator 1 | 3 | 3774 | 3 |
| IL18R1 | Interleukin 18 receptor 1 | 3 | 6563 | 3 |
| IL1R1 | Interleukin 1 receptor type 1 | 4 | 7979 | 7 |
| AP2A2 | Adaptor related protein complex 2 subunit alpha 2 | 9 | 45,285 | 15 |
| TOLLIP | Toll interacting protein | 4 | 26,625 | 14 |
| Trait | Both Treatment Groups | NDV 3 | Heat + NDV 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| h2 (SE) | N | Mean | h2 (SE) | N | Mean | h2 (SE) | N | Mean | |
| Viral load 2 dpi 1 | 0.24 (0.06) | 969 | 4.94 | 0.32 (0.10) | 468 | 5.10 | 0.17 (0.10) | 501 | 4.77 |
| Viral load 6 dpi 1 | 0.09 (0.04) | 965 | 3.22 | 0.18 (0.10) | 470 | 3.72 | 0.11 (0.08) | 495 | 2.76 |
| Antibody 10 dpi 1 | 0.14 (0.05) | 916 | −0.07 | 0.24 (0.09) | 448 | −0.04 | 0.04 (0.06) | 468 | −0.10 |
| Growth rate pre-challenge 2 | 0.40 (0.06) | 991 | 8.62 | 0.46 (0.11) | 473 | 10.4 | 0.27 (0.09) | 518 | 6.93 |
| Growth rate post-challenge 2 | 0.16 (0.05) | 969 | 10.97 | 0.21 (0.09) | 470 | 15.4 | 0.11 (0.06) | 499 | 6.7 |
| Trait | Time Point | Analysis (Treatment Group) | Gene | p-Value | Adjusted p-Value | Haplotype with Significant Additive Effect | p-Value for Additive Effect | Adjusted p-Value for Additive Effect |
|---|---|---|---|---|---|---|---|---|
| Viral load | 6 days post NDV | combined | SLC5A1 | 0.001 | 0.014 | |||
| NDV | SLC5A1 | 0.01 | 0.14 | |||||
| Antibody | 10 days post NDV | combined | IFI27L2 | 0.002 | 0.028 | H03 1 | 0.003 | 0.18 |
| NDV | IFI27L2 | 0.01 | 0.14 | H03 1 | 0.001 | 0.06 | ||
| Growth rate | post heat and NDV | combined | HSPA2 | 0.006 | 0.084 | |||
| NDV | IFRD1 | 0.002 | 0.028 | H01 2 | 0.001 | 0.06 | ||
| heat + NDV | HSPA2 | 0.001 | 0.014 | |||||
| BE | 9 days post heat; 2 days post NDV | heat + NDV | HSPA2 | 0.001 | 0.014 | |||
| heat + NDV | TLR7 | 0.021 | 0.098 | |||||
| heat + NDV | MX | 0.017 | 0.098 | |||||
| pH | 9 days post heat; 2 days post NDV | heat + NDV | MX | 0.004 | 0.056 | |||
| heat + NDV | IL1RL1 | 0.011 | 0.077 | |||||
| HCO3 | 9 days post heat; 2 days post NDV | heat + NDV | HSPA2 | 0.002 | 0.028 | |||
| TCO2 | 9 days post heat; 2 days post NDV | heat + NDV | HSPA2 | 0.003 | 0.042 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowland, K.; Saelao, P.; Wang, Y.; Fulton, J.E.; Liebe, G.N.; McCarron, A.M.; Wolc, A.; Gallardo, R.A.; Kelly, T.; Zhou, H.; et al. Association of Candidate Genes with Response to Heat and Newcastle Disease Virus. Genes 2018, 9, 560. https://doi.org/10.3390/genes9110560
Rowland K, Saelao P, Wang Y, Fulton JE, Liebe GN, McCarron AM, Wolc A, Gallardo RA, Kelly T, Zhou H, et al. Association of Candidate Genes with Response to Heat and Newcastle Disease Virus. Genes. 2018; 9(11):560. https://doi.org/10.3390/genes9110560
Chicago/Turabian StyleRowland, Kaylee, Perot Saelao, Ying Wang, Janet E. Fulton, Grant N. Liebe, Amy M. McCarron, Anna Wolc, Rodrigo A. Gallardo, Terra Kelly, Huaijun Zhou, and et al. 2018. "Association of Candidate Genes with Response to Heat and Newcastle Disease Virus" Genes 9, no. 11: 560. https://doi.org/10.3390/genes9110560
APA StyleRowland, K., Saelao, P., Wang, Y., Fulton, J. E., Liebe, G. N., McCarron, A. M., Wolc, A., Gallardo, R. A., Kelly, T., Zhou, H., Dekkers, J. C. M., & Lamont, S. J. (2018). Association of Candidate Genes with Response to Heat and Newcastle Disease Virus. Genes, 9(11), 560. https://doi.org/10.3390/genes9110560