Development of an Effective 6-Methylpurine Counterselection Marker for Genetic Manipulation in Thermococcus barophilus

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Media
2.2. Microbial Growth
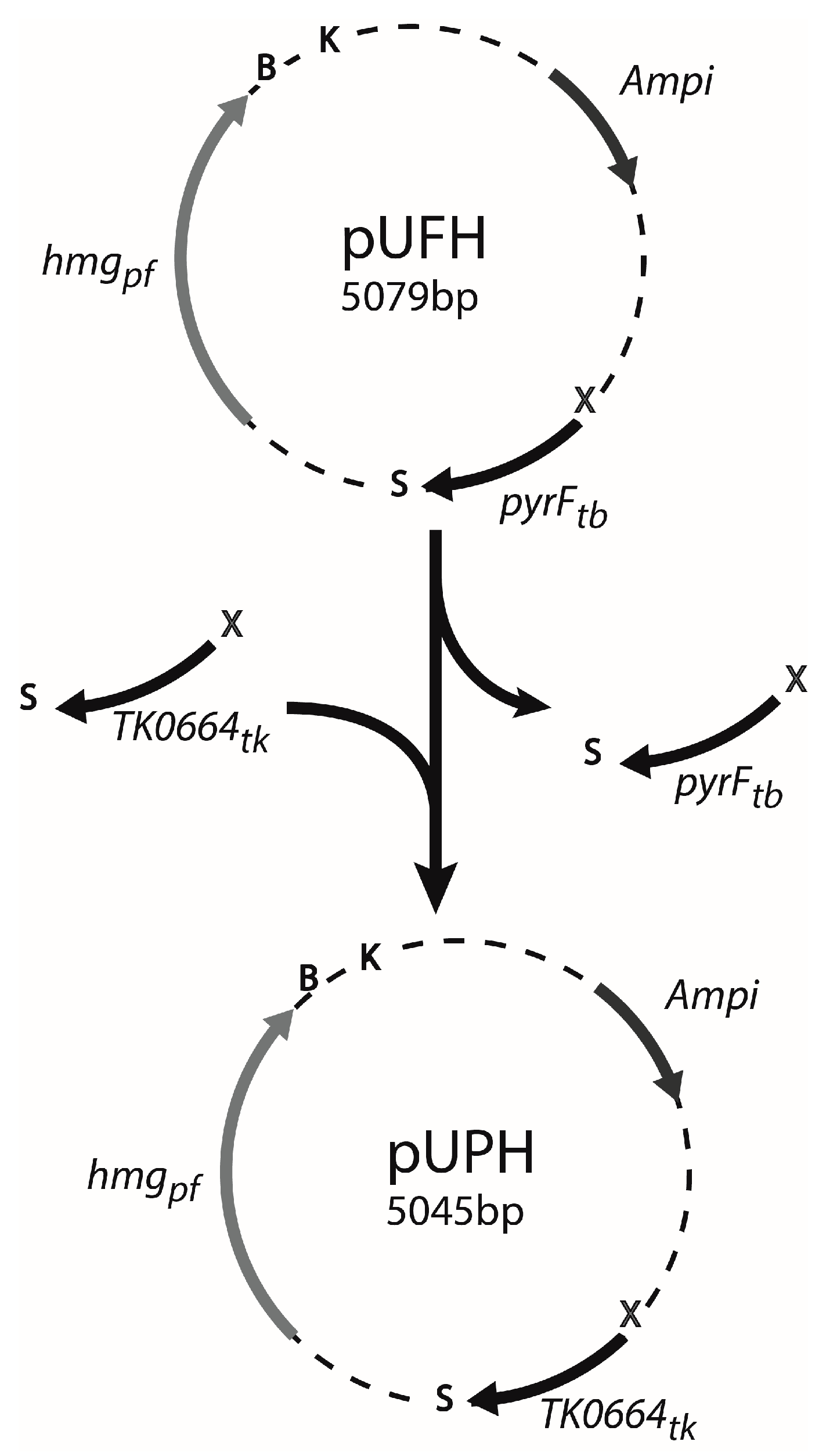
2.3. Construction of Gene Deletion Vectors
2.4. Transformation of Thermococcus barophilus
2.5. DNA Extraction and Purification
3. Results
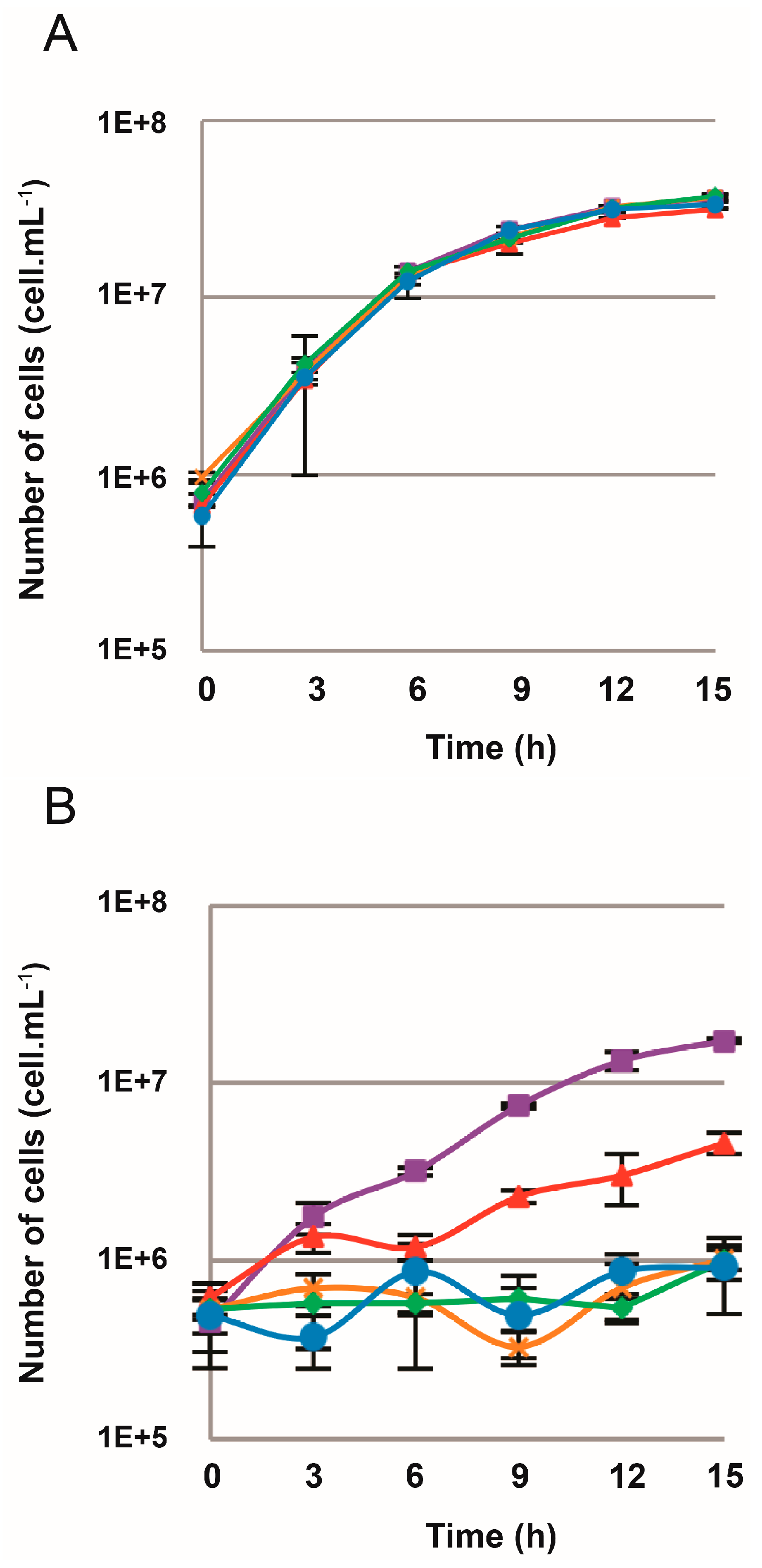
3.1. Sensitivity of T. barophilus MP to the Purine Analog 6-MP
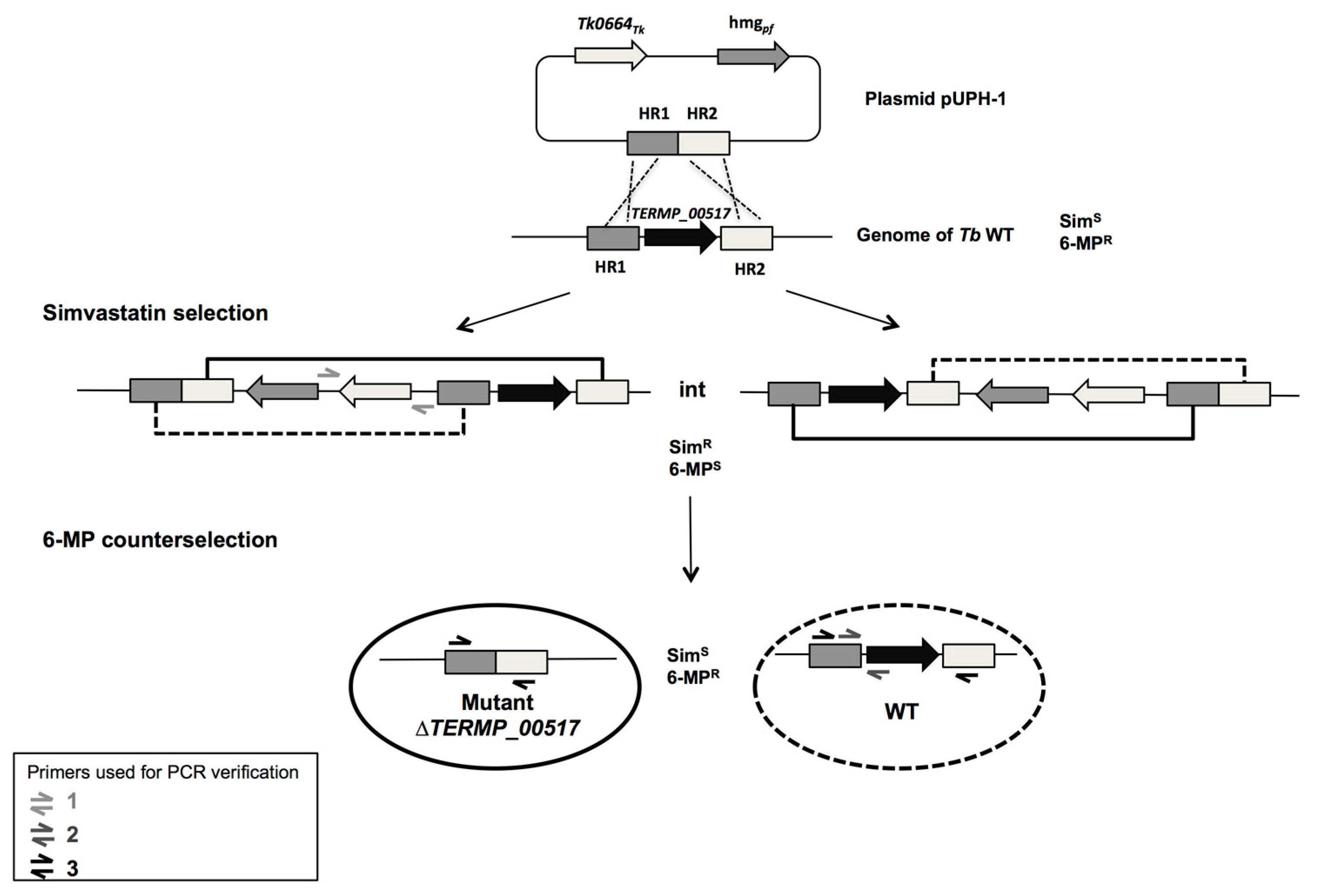
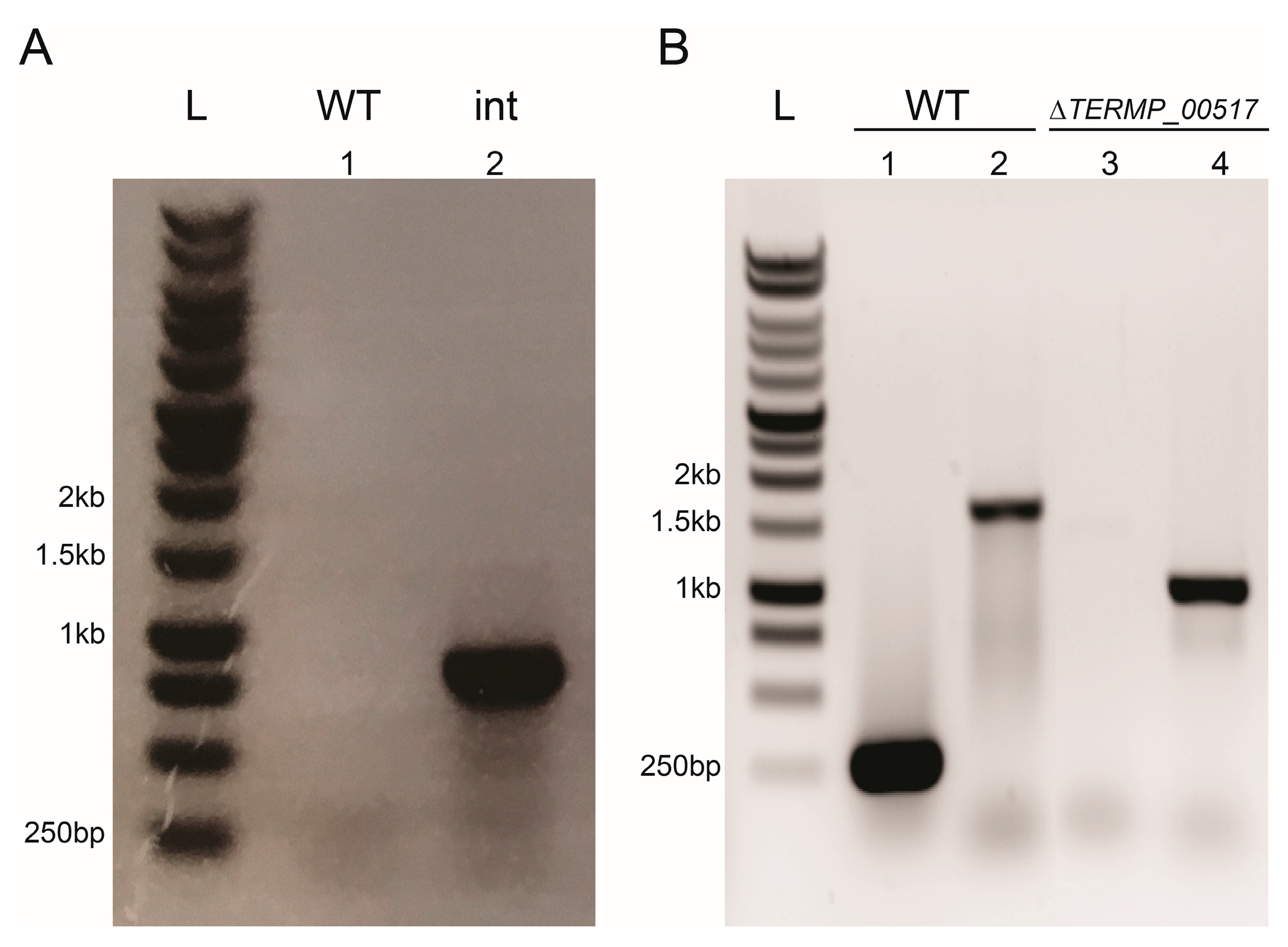
3.2. Construction of the ∆TERMP_00517
3.3. TERMP_00517 Is Responsible for 6-MP Sensitivity in Thermococcus barophilus
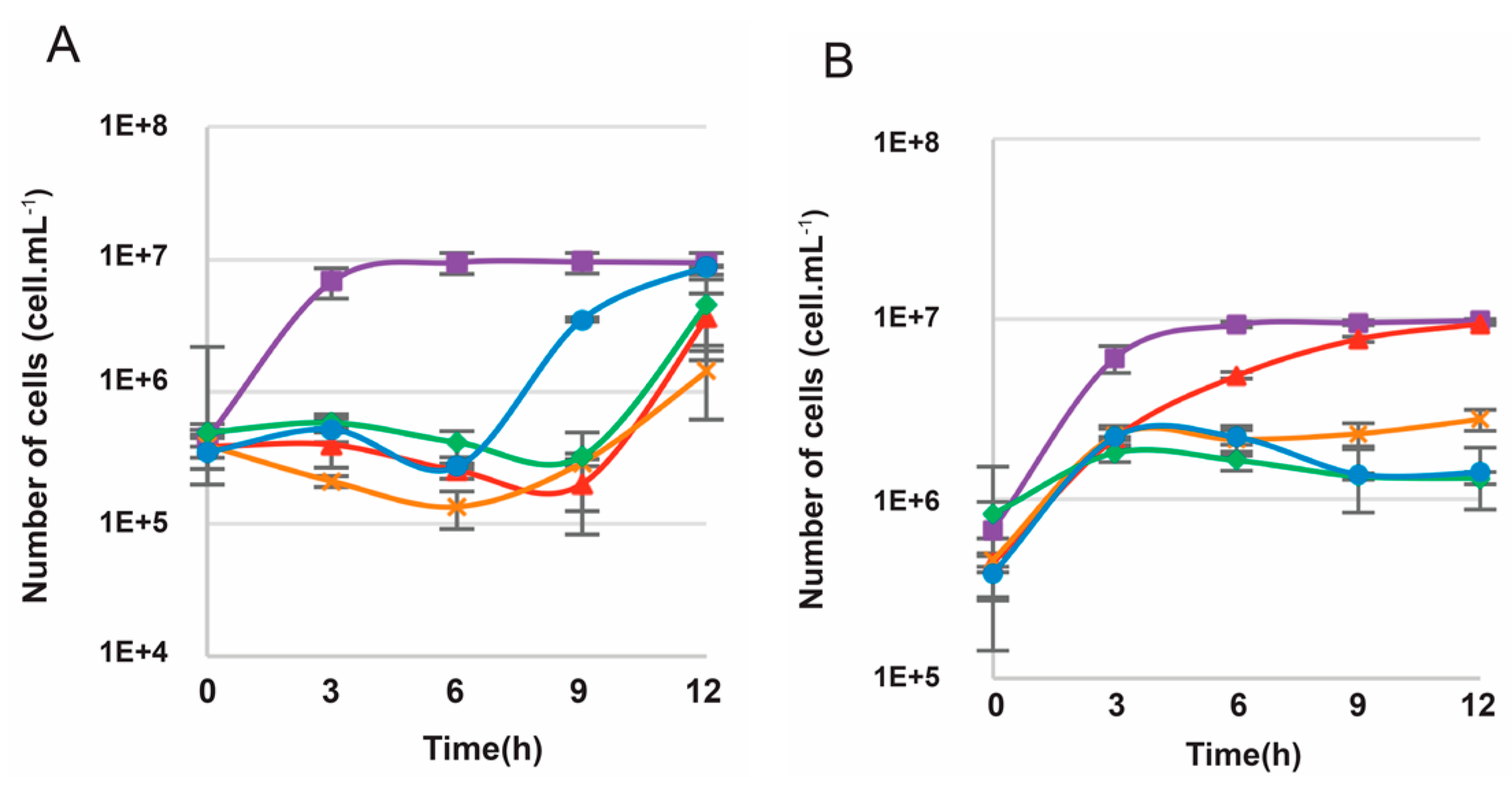
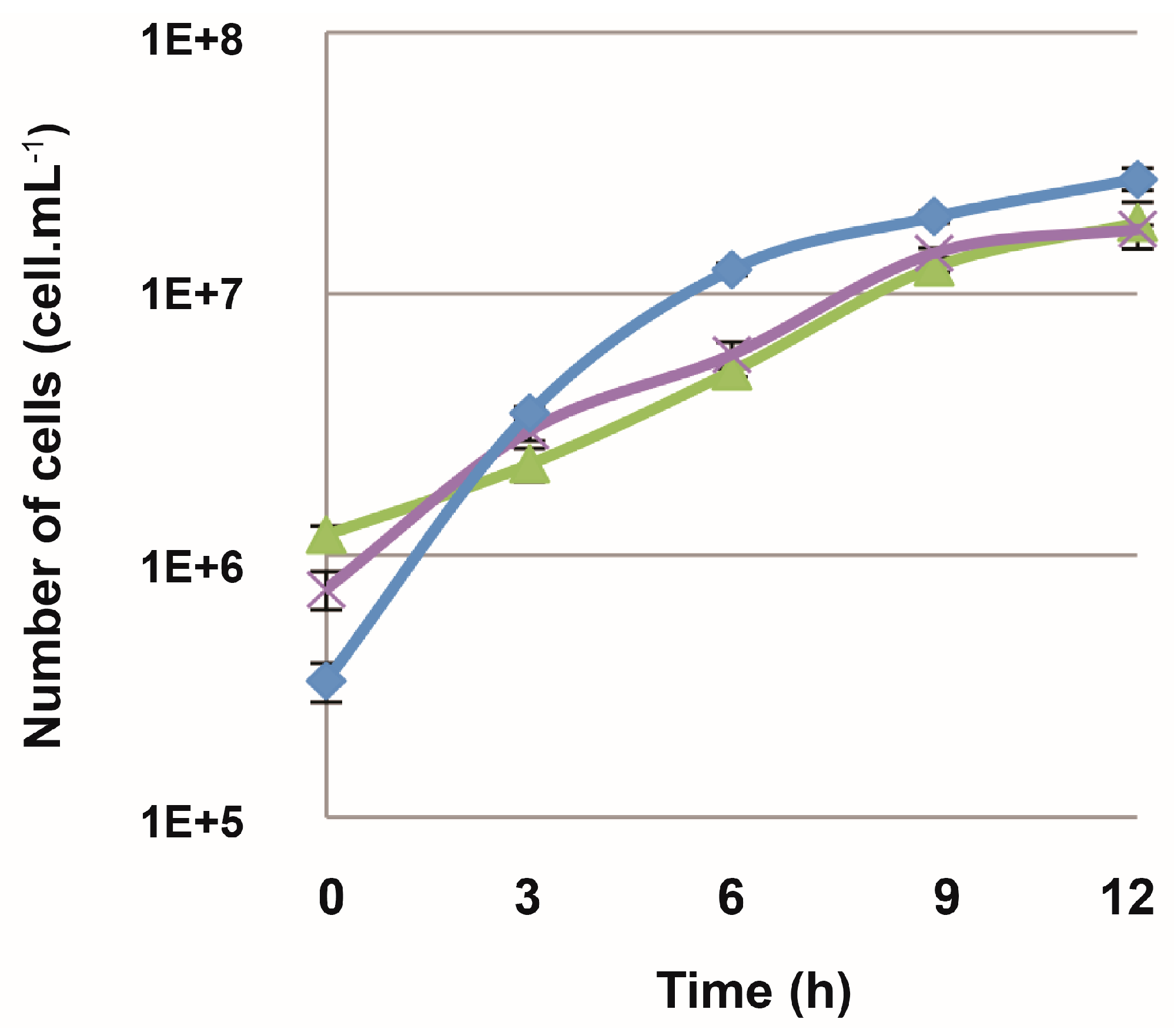
3.4. Efficiency of the New Counterselection Marker
3.5. Deletion of polB or rnhB Is Not Essential for Viability in Thermococcus barophilus
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Huber, R.; Stetter, K.O. Discovery of hyperthermophilic microorganisms. Methods Enzymol. 2001, 330, 11–24. [Google Scholar] [PubMed]
- Itoh, T. Taxonomy of nonmethanogenic hyperthermophilic and related thermophilic archaea. J. Biosci. Bioeng. 2003, 96, 203–212. [Google Scholar] [CrossRef]
- Fiala, G.; Stetter, K.O. Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100 °C. Arch. Microbiol. 1986, 145, 56–61. [Google Scholar] [CrossRef]
- Achenbach-Richter, L.; Gupta, R.; Zillig, W.; Woese, C.R. Rooting the archaebacterial tree: The pivotal role of Thermococcus celer in archaebacterial evolution. Syst. Appl. Microbiol. 1988, 10, 231–240. [Google Scholar] [CrossRef]
- Takai, K.; Sugai, A.; Itoh, T.; Horikoshi, K. Palaeococcus ferrophilus gen. nov., sp. nov., a barophilic hyperthermophilic archaeon from a deep-sea hydrothermal vent chimney. Int. J. Syst. Evol. Microbiol. 2000, 50, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, X.; Jiang, L.; Alain, K.; Jebbar, M.; Shao, Z. Palaeococcus pacificus sp. nov., an archaeon from deep-sea hydrothermal sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 2155–2159. [Google Scholar] [CrossRef] [PubMed]
- Jebbar, M.; Franzetti, B.; Girard, E.; Oger, P. Microbial diversity and adaptation to high hydrostatic pressure in deep-sea hydrothermal vents prokaryotes. Extremophiles 2015, 19, 721–740. [Google Scholar] [CrossRef] [PubMed]
- Leis, B.; Heinze, S.; Angelov, A.; Pham, V.T.; Thürmer, A.; Jebbar, M.; Golyshin, P.N.; Streit, W.R.; Daniel, R.; Liebl, W. Functional Screening of Hydrolytic Activities Reveals an Extremely Thermostable Cellulase from a Deep-Sea Archaeon. Front. Bioeng. Biotechnol. 2015, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Counts, J.A.; Zeldes, B.M.; Lee, L.L.; Straub, C.T.; Adams, M.W.W.; Kelly, R.M. Physiological, metabolic and biotechnological features of extremely thermophilic microorganisms. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Brasen, C.; Esser, D.; Rauch, B.; Siebers, B. Carbohydrate metabolism in Archaea: Current insights into unusual enzymes and pathways and their regulation. Microbiol. Mol. Biol. Rev. 2014, 78, 89–175. [Google Scholar] [CrossRef] [PubMed]
- Frock, A.D.; Kelly, R.M. Extreme Thermophiles: Moving beyond single-enzyme biocatalysis. Curr. Opin. Chem. Eng. 2012, 1, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Hedderich, R.; Klimmek, O.; Kröger, A.; Dirmeier, R.; Keller, M.; Stetter, K.O. Anaerobic respiration with elemental sulfur and with disulfides. FEMS Microbial. Rev. 1998, 22, 353–381. [Google Scholar] [CrossRef]
- Chou, C.J.; Shockley, K.R.; Conners, S.B.; Lewis, D.L.; Comfort, D.A.; Adams, M.W.; Kelly, R.M. Impact of substrate glycoside linkage and elemental sulfur on bioenergetics of and hydrogen production by the hyperthermophilic archaeon Pyrococcus furiosus. Appl. Environ. Microbiol. 2007, 73, 6842–6853. [Google Scholar] [CrossRef] [PubMed]
- Hensley, S.A.; Moreira, E.; Holden, J.F. Hydrogen Production and Enzyme Activities in the Hyperthermophile Thermococcus paralvinellae Grown on Maltose, Tryptone, and Agricultural Waste. Front. Microbiol. 2016, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Kwon, J.; Yun, S.H.; Lim, H.L.; Kim, J.; Kim, S.J.; Kang, S.G.; Lee, J.H.; Kim, S.I.; Chung, Y.H. Proteomic Insights into Sulfur Metabolism in the Hydrogen-Producing Hyperthermophilic Archaeon Thermococcus onnurineus NA1. Int. J. Mol. Sci. 2015, 16, 9167–9195. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, T.J.; Cubonova, L.; Reeve, J.N. Deletion of alternative pathways for reductant recycling in Thermococcus kodakarensis increases hydrogen production. Mol. Microbiol. 2011, 81, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Marteinsson, V.T.; Birrien, J.L.; Reysenbach, A.L.; Vernet, M.; Marie, D.; Gambacorta, A.; Messner, P.; Sleytr, U.B.; Prieur, D. Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 1999, 49, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Cario, A.; Mizgier, A.; Thiel, A.; Jebbar, M.; Oger, P.M. Restoration of the di-myo-inositol-phosphate pathway in the piezo-hyperthermophilic archaeon Thermococcus barophilus. Biochimie 2015, 118, 268–293. [Google Scholar] [CrossRef] [PubMed]
- Cario, A.; Jebbar, M.; Thiel, A.; Kervarec, N.; Oger, P.M. Molecular chaperone accumulation as a function of stress evidences adaptation to high hydrostatic pressure in the piezophilic archaeon Thermococcus barophilus. Sci. Rep. 2016, 6, 29483. [Google Scholar] [CrossRef] [PubMed]
- Vannier, P.; Michoud, G.; Oger, P.; Marteinsson, V.; Jebbar, M. Genome expression of Thermococcus barophilus and Thermococcus kodakarensis in response to different hydrostatic pressure conditions. Res. Microbiol. 2015, 166, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Allers, T.; Mevarech, M. Archaeal genetics—The third way. Nat. Rev. Genet. 2005, 6, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Atomi, H.; Imanaka, T.; Fukui, T. Overview of the genetic tools in the archaea. Front. Microbiol. 2012, 3, 337. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.A.; Picking, J.W.; Santangelo, T.J. Genetic techniques for the archaea. Annu. Rev. Genet. 2013, 47, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Hileman, T.H.; Santangelo, T.J. Genetics Techniques for Thermococcus kodakarensis. Front. Microbiol. 2012, 3, 195. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.A.; Albers, S.V.; Atomi, H.; Allers, T. Model organisms for genetics in the domain Archaea: Methanogens, halophiles, Thermococcales and Sulfolobales. FEMS Microbiol. Rev. 2011, 35, 577–608. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Fukui, T.; Atomi, H.; Imanaka, T. Improved and versatile transformation system allowing multiple genetic manipulations of the hyperthermophilic archaeon Thermococcus kodakaraensis. Appl. Environ. Microbiol. 2005, 71, 3889–3899. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, H.S.; Kim, E.S.; Bae, S.S.; Lim, J.K.; Matsumi, R.; Lebedinsky, A.V.; Sokolova, T.G.; Kozhevnikova, D.A.; Cha, S.S.; et al. Formate-driven growth coupled with H2 production. Nature 2010, 467, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, L.; Li, Z.; Ma, X.; Xiao, X.; Xu, J. Genetic tools for the piezophilic hyperthermophilic archaeon Pyrococcus yayanosii. Extremophiles 2015, 19, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lipscomb, G.L.; Stirrett, K.; Schut, G.J.; Yang, F.; Jenney, F.E., Jr.; Scott, R.A.; Adams, M.W.; Westpheling, J. Natural competence in the hyperthermophilic archaeon Pyrococcus furiosus facilitates genetic manipulation: Construction of markerless deletions of genes encoding the two cytoplasmic hydrogenases. Appl. Environ. Microbiol. 2011, 77, 2232–2238. [Google Scholar] [CrossRef] [PubMed]
- Matsumi, R.; Manabe, K.; Fukui, T.; Atomi, H.; Imanaka, T. Disruption of a sugar transporter gene cluster in a hyperthermophilic archaeon using a host-marker system based on antibiotic resistance. J. Bacteriol. 2007, 189, 2683–2691. [Google Scholar] [CrossRef] [PubMed]
- Thiel, A.; Michoud, G.; Moalic, Y.; Flament, D.; Jebbar, M. Genetic Manipulations of the Hyperthermophilic Piezophilic Archaeon Thermococcus barophilus. Appl. Environ. Microbiol. 2014, 80, 2299–2306. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Murakami, A.; Yoshida, K. Counterselection system for Geobacillus kaustophilus HTA426 through disruption of pyrF and pyrR. Appl. Environ. Microbiol. 2012, 78, 7376–7383. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.A.; Elion, G.B.; Hitchings, G.H.; Stock, C.C. Structure-activity relationships among purines related to 6-mercaptopurine. Cancer Res. 1958, 18, 445–456. [Google Scholar] [PubMed]
- Dewey, V.C.; Heinrich, M.R.; Markees, D.G.; KidderI, G.W. Multiple inhibition by 6-methylpurine. Biochem. Pharm. 1960, 3, 173–180. [Google Scholar] [CrossRef]
- Freese, E. The specific mutagenic effect of bases analogues on phage T 4. J. Mol. Biol. 1959, 1, 87–105. [Google Scholar] [CrossRef]
- Miller, J.H.; Kempner, E.S. Effets of an adenine analog on yeast metabolism. Biochim. Biophys. Acta 1963, 76, 333–340. [Google Scholar] [CrossRef]
- Santangelo, T.J.; Cubonova, L.; Reeve, J.N. Thermococcus kodakarensis genetics: TK1827-encoded beta-glycosidase, new positive-selection protocol, and targeted and repetitive deletion technology. Appl. Environ. Microbiol. 2010, 76, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, B.W.; Cubonova, L.; Heider, M.R.; Kelman, Z.; Reeve, J.N.; Santangelo, T.J. The GAN exonuclease or the flap endonuclease Fen1 and RNase HII are necessary for viability of Thermococcus kodakarensis. J. Bacteriol. 2017, 199, e00141-117. [Google Scholar] [CrossRef] [PubMed]
- Cubonova, L.; Richardson, T.; Burkhart, B.W.; Kelman, Z.; Connolly, B.A.; Reeve, J.N.; Santangelo, T.J. Archaeal DNA polymerase D but not DNA polymerase B is required for genome replication in Thermococcus kodakarensis. J. Bacteriol. 2013, 195, 2322–2328. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Birrien, J.L.; Fouquet, Y.; Cherkashov, G.; Jebbar, M.; Querellou, J.; Oger, P.; Cambon-Bonavita, M.A.; Xiao, X.; Prieur, D. Pyrococcus CH1, an obligate piezophilic hyperthermophile: Extending the upper pressure-temperature limits for life. ISME J. 2009, 3, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Waege, I.; Schmid, G.; Thumann, S.; Thomm, M.; Hausner, W. Shuttle vector-based transformation system for Pyrococcus furiosus. Appl. Environ. Microbiol. 2010, 76, 3308–3313. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Fukui, T.; Atomi, H.; Imanaka, T. Targeted gene disruption by homologous recombination in the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1. J. Bacteriol. 2003, 185, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Vannier, P.; Marteinsson, V.T.; Fridjonsson, O.H.; Oger, P.; Jebbar, M. Complete genome sequence of the hyperthermophilic, piezophilic, heterotrophic, and carboxydotrophic archaeon Thermococcus barophilus MP. J. Bacteriol. 2011, 193, 1481–1482. [Google Scholar] [CrossRef] [PubMed]
- Le Laz, S.; Le Goaziou, A.; Henneke, G. Structure-specific nuclease activities of Pyrococcus abyssi RNase HII. J. Bacteriol. 2010, 192, 3689–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; She, Q.; Bi, H.; Whitaker, R.J. The apt/6-methylpurine counterselection system and its applications in genetic studies in the hyperthermophilic archaeon Sulfolobus islandicus. Appl. Environ. Microbiol. 2016, 82, 3070–3081. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Parent Strain | References |
|---|---|---|---|
| UBOCC-M-3203 | Wild Type | T. kodakarensis KOD1 | [22] |
| UBOCC-M-3107 | Wild Type | T. barophilus MP | [17] |
| UBOCC-M-3300 | TB∆TERMP_00517 | T. barophilus MP | This study |
| UBOCC-M-3301 | TB∆TERMP_00517::TK0664 | T. barophilus MP | This study |
| UBOCC-M-3302 | TB∆TERMP_00517, ∆TERMP_01623 | T. barophilus MP | This study |
| UBOCC-M-3303 | TB∆TERMP_00517, ∆TERMP_00671 | T. barophilus MP | This study |
| Primers Used for Amplification of Flanking Regions of Targeted Gene | Sequence (5′-3′) | Tm (°C) |
|---|---|---|
| PolB_1Up | AAAAAAGGTACCGCTTAACATTCCTGACTCCCAGAATCTT | 59.4 |
| PolB_1Do | TCTATTTCATTAAATCACCTAATTTCACCCTTTTAAAAATACATGCCCAT | 57.9 |
| PolB_2Up | ATTTTTAAAAGGGTGAAATTAGGTGATTTAATGAAATAGAATGAGCAGGA | 57.9 |
| PolB_2Do | AAAAAAGGATCCCGGCTTCTGGGGAAACCTCG | 60.6 |
| 6MPb_1Up | AAAAAACGTACCAAGAAAACCGGAGTTTTAGTGAATACACC | 58.4 |
| 6MPb_1Do | TCTCATGGAAACATTTAAATGGTTGTGGTATCTTGGACAAGAAGAAAA | 59.5 |
| 6MPb_2Up | TTGTCCAAGATACCACAACCATTTAAATGTTTCCATGAGAAAAATGAAATGCAAAAA | 60.3 |
| 6MPb_2Do | AAAAAAAGATCTCTCGCTCTAAAGGAGCTTTCAACA | 56.0 |
| RHII_1Up | AAAAAAGGTACCCGGTACCCTGATAAAGAAGGCATC | 59.4 |
| RHII_1Do | TTAGTATTCAGGAAATGAGGACTCTTGAGGTTCTTTCTCTTCGGT | 61.3 |
| RHII_2_Up | AGAGAAAGAACCTCAAGAGTCCTCATTTCCTGAATACTAACGTTCCG | 63.0 |
| RHII_2_Do | AAAAAAAGATCTACGACTCATGATGTTAATCCTCTTACAGAG | 57.4 |
| Primers Used to Check Mutants | Sequence (5′-3′) | Tm (°C) |
| PolB_Up | GTCAGCTATGAGCTCGTGAAGAGTTAT | 59 |
| PolB_Do | AGAAGAGCGTAAATGTAAGGCTGGA | 59 |
| 6MPk_Up | AAAAAACTCGAGCCCGTCCAAGCTACCACTCCC | 69 |
| 6MPk_Do | AAAAAACCCGGCTTCAAAACCCAGCCAAACAACACCC | 71 |
| 6MPb_Up | ACCTCAGACATCTCTGCTGCTTG | 60 |
| 6MPb_Do | GCCCCGATAAGTGCTGAAAGATACA | 60 |
| RHII_Up | GAGGAATCGCTGAAGTTTTACTGGG | 59 |
| RHII_Do | CTACAAAGAGCATGGCGAATTTCCG | 60 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birien, T.; Thiel, A.; Henneke, G.; Flament, D.; Moalic, Y.; Jebbar, M. Development of an Effective 6-Methylpurine Counterselection Marker for Genetic Manipulation in Thermococcus barophilus. Genes 2018, 9, 77. https://doi.org/10.3390/genes9020077
Birien T, Thiel A, Henneke G, Flament D, Moalic Y, Jebbar M. Development of an Effective 6-Methylpurine Counterselection Marker for Genetic Manipulation in Thermococcus barophilus. Genes. 2018; 9(2):77. https://doi.org/10.3390/genes9020077
Chicago/Turabian StyleBirien, Tiphaine, Axel Thiel, Ghislaine Henneke, Didier Flament, Yann Moalic, and Mohamed Jebbar. 2018. "Development of an Effective 6-Methylpurine Counterselection Marker for Genetic Manipulation in Thermococcus barophilus" Genes 9, no. 2: 77. https://doi.org/10.3390/genes9020077