Wnt Signaling in Stem Cell Maintenance and Differentiation in the Drosophila Germarium

Abstract
:1. Introduction
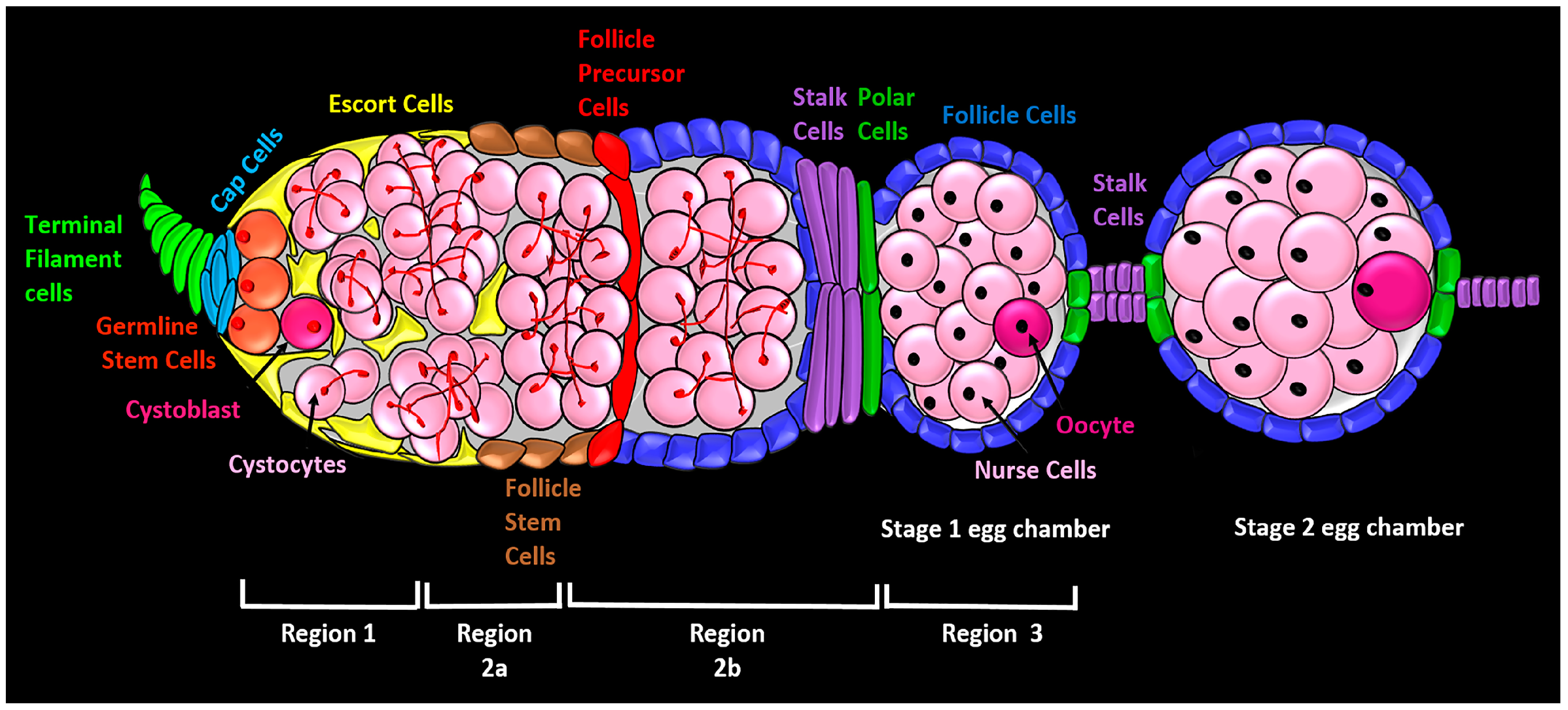
2. Anatomy of the Germarium and an Overview of Egg Chamber Development
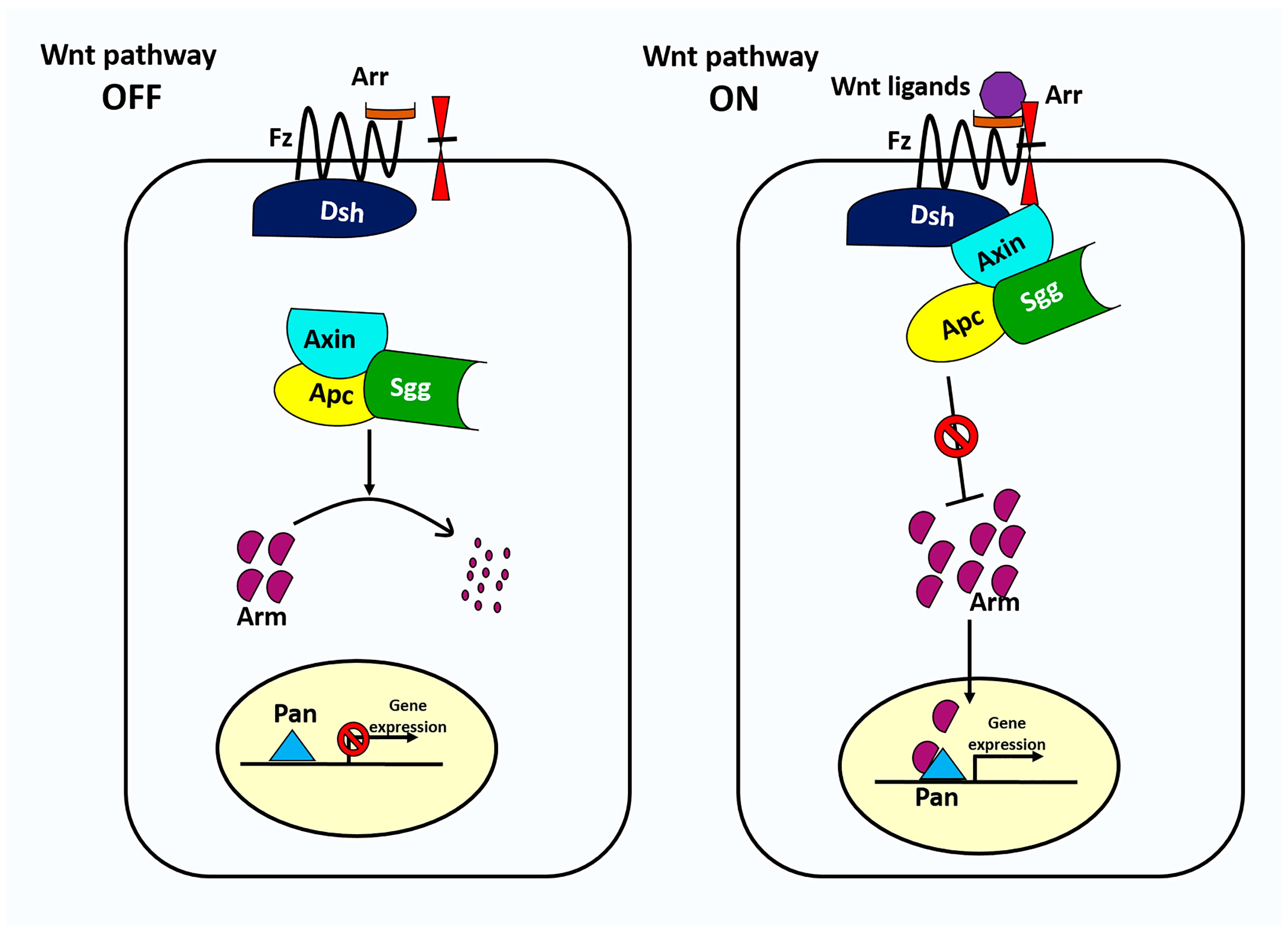
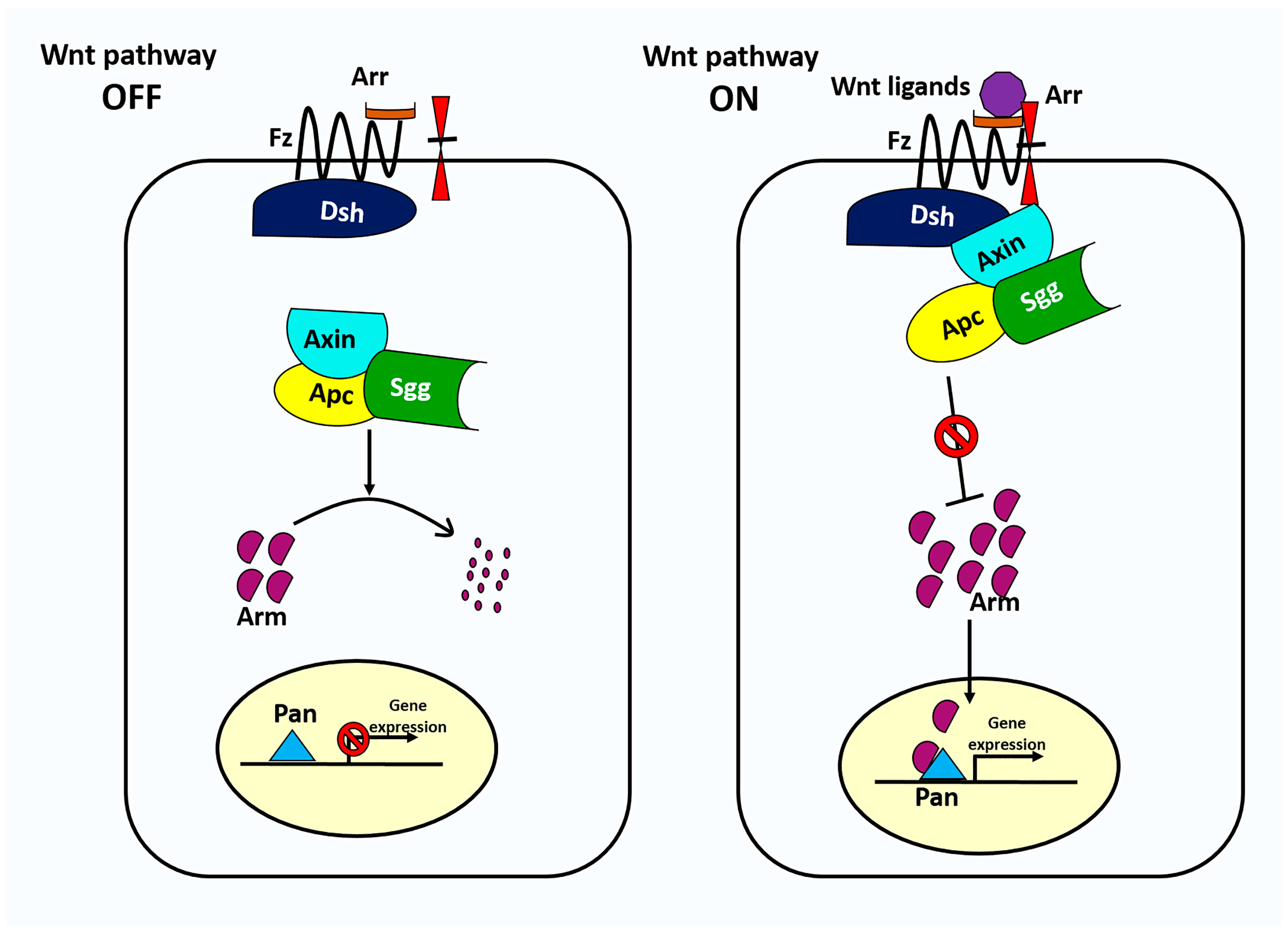
3. The Wnt Signaling Pathway
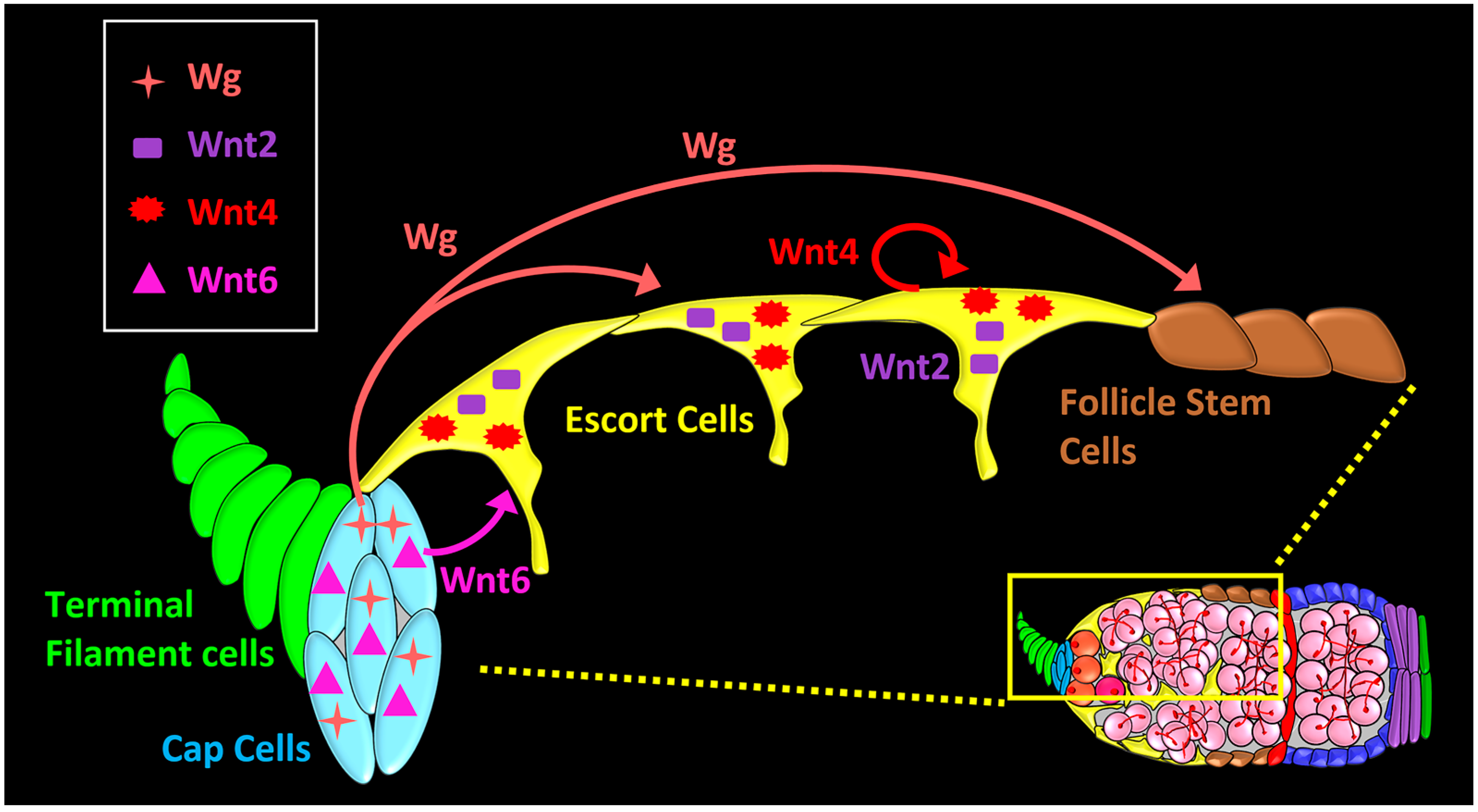
4. Wnt Pathway Functions in the Germarium
4.1. How Wnt Signaling Affects Maintenance of Germline Stem Cells
4.2. How Wnt Signaling Affects GSC Differentiation
4.3. How Wnt Signaling Affects Follicle Stem Cells Differentiation and Proliferation
4.4. How Wnt Signaling Affects Pupal Development of the Germarium
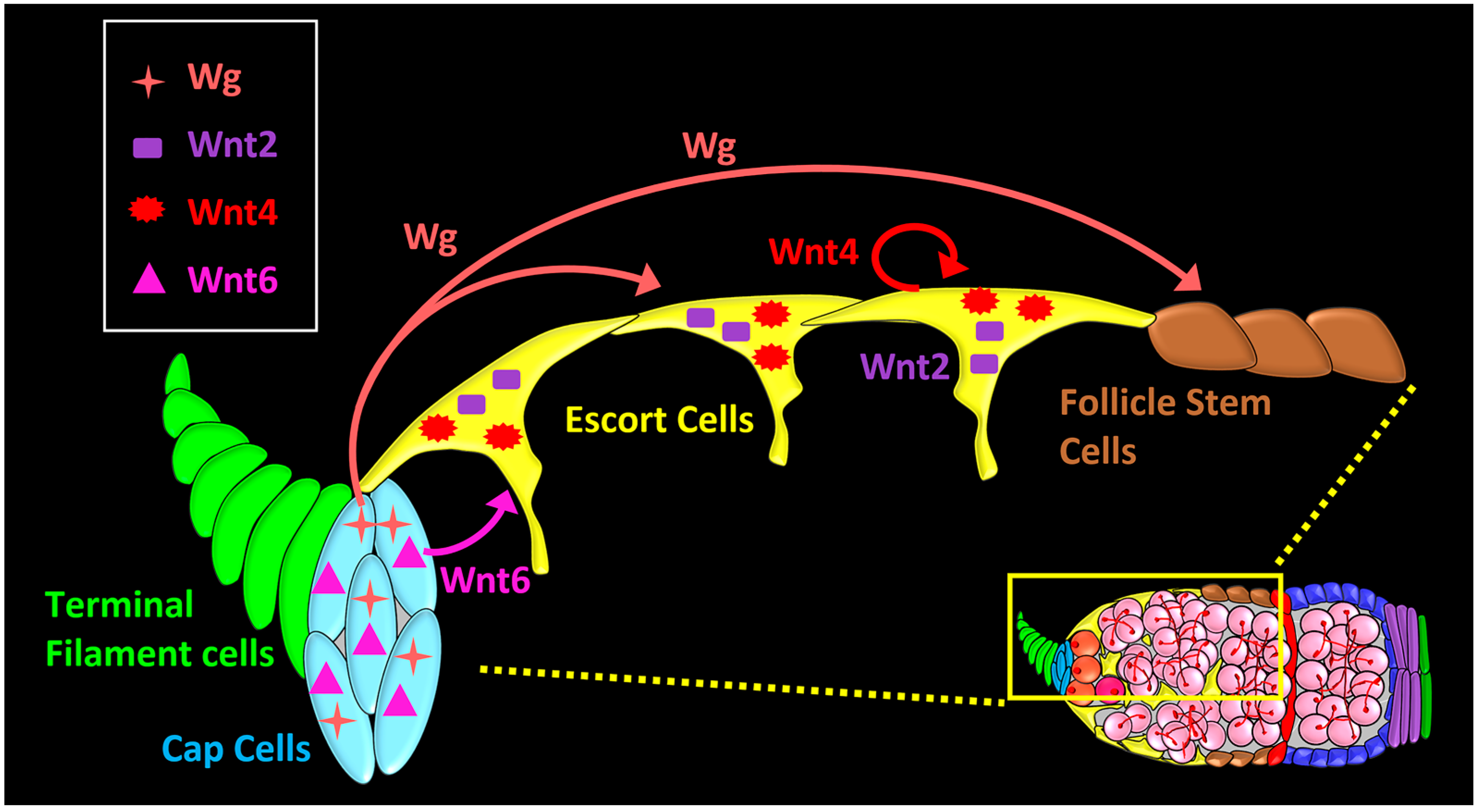
5. Long-Range and Short-Range Wnt Signaling
6. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Xie, T.; Spradling, A.C. A niche maintaining germ line stem cells in the Drosophila ovary. Science 2000, 290, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhu, C.H.; Doan, C.; Xie, T. Germline stem cells anchored by adherens junctions in the Drosophila ovary niches. Science 2002, 296, 1855–1857. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.X.; Spradling, A.C. Long-term live imaging provides new insight into stem cell regulation and germline-soma coordination in the Drosophila ovary. Development 2011, 138, 2207–2215. [Google Scholar] [CrossRef] [PubMed]
- Decotto, E.; Spradling, A.C. The Drosophila ovarian and testis stem cell niches: Similar somatic stem cells and signals. Dev. Cell 2005, 9, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Spradling, A.C. Decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell 1998, 94, 251–260. [Google Scholar] [CrossRef]
- Cox, D.N.; Chao, A.; Baker, J.; Chang, L.; Qiao, D.; Lin, H. A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal. Genes Dev. 1998, 12, 3715–3727. [Google Scholar] [CrossRef] [PubMed]
- King, F.J.; Lin, H. Somatic signaling mediated by fs(1)Yb is essential for germline stem cell maintenance during Drosophila oogenesis. Development 1999, 126, 1833. [Google Scholar] [PubMed]
- Kirilly, D.; Wang, S.; Xie, T. Self-maintained escort cells form a germline stem cell differentiation niche. Development 2011, 138, 5087–5097. [Google Scholar] [CrossRef] [PubMed]
- Ehrman, L. Ovarian Development in Drosophila melanogaster. Robert C. King. Q. Rev. Biol. 1971, 46, 79. [Google Scholar] [CrossRef]
- Horne-Badovinac, S.; Bilder, D. Mass transit: Epithelial morphogenesis in the Drosophila egg chamber. Dev. Dyn. 2005, 232, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kalderon, D. Hedgehog acts as a somatic stem cell factor in the Drosophila ovary. Nature 2001, 410, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Margolis, J.; Spradling, A. Identification and behavior of epithelial stem cells in the Drosophila ovary. Development 1995, 121, 3797–3807. [Google Scholar] [PubMed]
- Nystul, T.; Spradling, A. An epithelial niche in the Drosophila ovary undergoes long-range stem cell replacement. Cell Stem Cell 2007, 1, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Nystul, T.; Spradling, A. Regulation of epithelial stem cell replacement and follicle formation in the Drosophila ovary. Genetics 2010, 184, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Reilein, A.; Melamed, D.; Park, K.S.; Berg, A.; Cimetta, E.; Tandon, N.; Vunjak-Novakovic, G.; Finkelstein, S.; Kalderon, D. Alternative direct stem cell derivatives defined by stem cell location and graded Wnt signalling. Nat. Cell Biol. 2017, 19, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Peterson, A.; Kenney, T.; Burrous, H.; Montell, D.J. Quantitative microscopy of the Drosophila ovary shows multiple niche signals specify progenitor cell fate. Nat. Commun. 2017, 8, 1244. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Lin, H. Spectrosomes and fusomes anchor mitotic spindles during asymmetric germ cell divisions and facilitate the formation of a polarized microtubule array for oocyte specification in Drosophila. Dev. Biol. 1997, 189, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Spradling, A.C. Fusome asymmetry and oocyte determination in Drosophila. Dev. Genet. 1995, 16, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Swarup, S.; Verheyen, E.M. Wnt/Wingless signaling in Drosophila. Cold Spring Harb. Perspect. Biol. 2012, 4, a007930. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Tian, A.; Jiang, J. Intestinal stem cell response to injury: Lessons from Drosophila. Cell. Mol. Life Sci. 2016, 73, 3337–3349. [Google Scholar] [CrossRef] [PubMed]
- Saito-Diaz, K.; Chen, T.W.; Wang, X.; Thorne, C.A.; Wallace, H.A.; Page-McCaw, A.; Lee, E. The way Wnt works: Components and mechanism. Growth Factors 2013, 31, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Fanto, M.; McNeill, H. Planar polarity from flies to vertebrates. J. Cell Sci. 2004, 117, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mlodzik, M. Wnt-Frizzled/planar cell polarity signaling: Cellular orientation by facing the wind (Wnt). Annu. Rev. Cell Dev. Biol. 2015, 31, 623–646. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Loh, K.M.; Nusse, R. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 2014, 346, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/β-Catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Kim, Y.-M.; Kahn, M. Wnt/Catenin signaling in adult stem cell physiology and disease. Stem Cell Rev. Rep. 2014, 10, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Mah, A.T.; Yan, K.S.; Kuo, C.J. Wnt pathway regulation of intestinal stem cells. J. Physiol. 2016, 594, 4837–4847. [Google Scholar] [CrossRef] [PubMed]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef]
- Nusse, R.; Brown, A.; Papkoff, J.; Scambler, P.; Shackleford, G.; McMahon, A.; Moon, R.; Varmus, H. A new nomenclature for int-1 and related genes: The Wnt gene family. Cell 1991, 64, 231. [Google Scholar] [CrossRef]
- Bejsovec, A. Wingless/Wnt signaling in Drosophila: The pattern and the pathway. Mol. Reprod. Dev. 2013, 80, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K.; Clevers, H. Wnt/β-catenin signaling in adult mammalian epithelial stem cells. Dev. Biol. 2017, 428, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wang, H.; Fan, C.; Liu, S.; Cai, Y. Wnt ligands regulate Tkv expression to constrain Dpp activity in the Drosophila ovarian stem cell niche. J. Cell Biol. 2015, 209, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, Y.; Song, X.; Ma, X.; Zhu, X.; Mao, Y.; Yang, Z.; Ni, J.; Li, H.; Malanowski, K.E.; et al. Wnt signaling-mediated redox regulation maintains the germ line stem cell differentiation niche. eLife 2015, 4, e08174. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.J.; Spradling, A.C.; Ingham, P.W.; Lin, H. The role of segment polarity genes during early oogenesis in Drosophila. Development 1996, 122, 3283–3294. [Google Scholar] [PubMed]
- Wang, X.; Page-McCaw, A. A matrix metalloproteinase mediates long-distance attenuation of stem cell proliferation. J. Cell Biol. 2014, 206, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Xie, T. Wingless signaling regulates the maintenance of ovarian somatic stem cells in Drosophila. Development 2003, 130, 3259–3268. [Google Scholar] [CrossRef] [PubMed]
- Seto, E.S.; Bellen, H.J. The ins and outs of Wingless signaling. Trends Cell Biol. 2004, 14, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gaitan, M. Endocytic trafficking during Drosophila development. Mech. Dev. 2003, 120, 1265–1282. [Google Scholar] [CrossRef] [PubMed]
- Zecca, M.; Basler, K.; Struhl, G. Direct and long-range action of a Wingless morphogen gradient. Cell 1996, 87, 833–844. [Google Scholar] [CrossRef]
- Neumann, C.J.; Cohen, S.M. Long-range action of Wingless organizes the dorsal-ventral axis of the Drosophila wing. Development 1997, 124, 871–880. [Google Scholar] [PubMed]
- Port, F.; Basler, K. Wnt trafficking: New insights into Wnt maturation, secretion and spreading. Traffic 2010, 11, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Vied, C.; Reilein, A.; Field, N.S.; Kalderon, D. Regulation of stem cells by intersecting gradients of long-range niche signals. Dev. Cell 2012, 23, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Page-McCaw, A. Wnt6 maintains anterior escort cells as an integral component of the germline stem cell niche. Development 2018, 145, dev158527. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lim, T.M.; Cai, Y. The Drosophila female germline stem cell lineage acts to spatially restrict DPP function within the niche. Sci. Signal. 2010, 3, ra57. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Rios, P.; Guerrero, I.; Gonzalez-Reyes, A. Cytoneme-mediated delivery of Hedgehog regulates the expression of Bone Morphogenetic Proteins to maintain germline stem cells in Drosophila. PLoS Biol. 2012, 10, e1001298. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wong, M.D.; Kawase, E.; Xi, R.; Ding, B.C.; McCarthy, J.J.; Xie, T. BMP signals from niche cells directly repress transcription of a differentiation-promoting gene, bag of marbles, in germline stem cells in the Drosophila ovary. Development 2004, 131, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, M.; Martino Cortez, Y.; Wong-Deyrup, S.; Tavares, L.; Schowalter, S.; Flora, P.; Hill, C.; Nasrallah, M.A.; Chittur, S.; Rangan, P. Transposon dysregulation modulates dWnt4 signaling to control germline stem cell differentiation in Drosophila. PLoS Genet. 2016, 12, e1005918. [Google Scholar] [CrossRef] [PubMed]
- Mottier-Pavie, V.I.; Palacios, V.; Eliazer, S.; Scoggin, S.; Buszczak, M. The Wnt pathway limits BMP signaling outside of the germline stem cell niche in Drosophila ovaries. Dev. Biol. 2016, 417, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Hamada-Kawaguchi, N.; Nore, B.F.; Kuwada, Y.; Smith, C.I.; Yamamoto, D. Btk29A promotes Wnt4 signaling in the niche to terminate germ cell proliferation in Drosophila. Science 2014, 343, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Eliazer, S.; Palacios, V.; Wang, Z.; Kollipara, R.K.; Kittler, R.; Buszczak, M. Lsd1 restricts the number of germline stem cells by regulating multiple targets in escort cells. PLoS Genet. 2014, 10, e1004200. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Flynt, A.S.; Lai, E.C. Drosophila piwi mutants exhibit germline stem cell tumors that are sustained by elevated Dpp signaling. Curr. Biol. 2013, 23, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, S.; Do, T.; Song, X.; Inaba, M.; Nishimoto, Y.; Liu, L.P.; Gao, Y.; Mao, Y.; Li, H.; et al. Piwi is required in multiple cell types to control germline stem cell lineage development in the Drosophila ovary. PLoS ONE 2014, 9, e90267. [Google Scholar]
- Mukai, M.; Kato, H.; Hira, S.; Nakamura, K.; Kita, H.; Kobayashi, S. Innexin2 gap junctions in somatic support cells are required for cyst formation and for egg chamber formation in Drosophila. Mech. Dev. 2011, 128, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pan, L.; Wang, S.; Zhou, J.; McDowell, W.; Park, J.; Haug, J.; Staehling, K.; Tang, H.; Xie, T. Histone H3K9 trimethylase Eggless controls germline stem cell maintenance and differentiation. PLoS Genet. 2011, 7, e1002426. [Google Scholar] [CrossRef] [PubMed]
- Rangan, P.; Malone, C.D.; Navarro, C.; Newbold, S.P.; Hayes, P.S.; Sachidanandam, R.; Hannon, G.J.; Lehmann, R. piRNA production requires heterochromatin formation in Drosophila. Curr. Biol. 2011, 21, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.; Xin, T.; He, J.; Tan, J.; Gao, Y.; Feng, S.; He, L.; Zhao, G.; Li, M. dBre1/dSet1-dependent pathway for histone H3K4 trimethylation has essential roles in controlling germline stem cell maintenance and germ cell differentiation in the Drosophila ovary. Dev. Biol. 2013, 379, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Aravin, A.A.; Hannon, G.J.; Brennecke, J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science 2007, 318, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.J.; Weiner, M.M.; Lin, H. PIWI proteins and PIWI-interacting RNAs in the soma. Nature 2014, 505, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Guzzardo, P.M.; Muerdter, F.; Hannon, G.J. The piRNA pathway in flies: Highlights and future directions. Curr. Opin. Genet. Dev. 2013, 23, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Konig, A.; Shcherbata, H.R. Soma influences GSC progeny differentiation via the cell adhesion-mediated steroid-let-7-Wingless signaling cascade that regulates chromatin dynamics. Biol. Open 2015, 4, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Toung, Y.P.; Hsieh, T.S.; Tu, C.P. The Glutathione S-Transferase D genes. A divergently organized, intronless gene family in Drosophila melanogaster. J. Biol. Chem. 1993, 268, 9737–9746. [Google Scholar] [PubMed]
- Nickla, H.; Anderson, J.; Palzkill, T. Enzymes involved in oxygen detoxification during development of Drosophila melanogaster. Experientia 1983, 39, 610–612. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, A.M.; Lee, H.H.; Simon, M.A. Integrins control the positioning and proliferation of follicle stem cells in the Drosophila ovary. J. Cell Biol. 2008, 182, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.J.; Lin, H.; Ingham, P.W.; Spradling, A.C. Hedgehog is required for the proliferation and specification of ovarian somatic cells prior to egg chamber formation in Drosophila. Development 1996, 122, 1125–1135. [Google Scholar] [PubMed]
- Wang, X.; Page-McCaw, A.; (Vanderbilt University, Nashville, TN). Unpublished data. 2015.
- Alexandre, C.; Baena-Lopez, A.; Vincent, J.P. Patterning and growth control by membrane-tethered Wingless. Nature 2014, 505, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Han, Y.; Xi, R. Polycomb group genes Psc and Su(z)2 restrict follicle stem cell self-renewal and extrusion by controlling canonical and noncanonical Wnt signaling. Genes Dev. 2010, 24, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.D.; Mariol, M.C.; Wallace, R.M.; Weyers, J.; Kamberov, Y.G.; Pradel, J.; Wilder, E.L. DWnt4 regulates cell movement and focal adhesion kinase during Drosophila ovarian morphogenesis. Dev. Cell 2002, 2, 437–448. [Google Scholar] [CrossRef]
- Stanganello, E.; Scholpp, S. Role of cytonemes in Wnt transport. J. Cell Sci. 2016, 129, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Sahai-Hernandez, P.; Nystul, T.G. A dynamic population of stromal cells contributes to the follicle stem cell niche in the Drosophila ovary. Development 2013, 140, 4490–4498. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Weber, F.A.; Kornberg, T.B. Cytonemes: Cellular processes that project to the principal signaling center in Drosophila imaginal discs. Cell 1999, 97, 599–607. [Google Scholar] [CrossRef]
- Kornberg, T.B.; Roy, S. Cytonemes as specialized signaling filopodia. Development 2014, 141, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kornberg, T.B. Myoblast cytonemes mediate Wg signaling from the wing imaginal disc and Delta-Notch signaling to the air sac primordium. eLife 2015, 4, e06114. [Google Scholar] [CrossRef] [PubMed]
- Stanganello, E.; Hagemann, A.I.; Mattes, B.; Sinner, C.; Meyen, D.; Weber, S.; Schug, A.; Raz, E.; Scholpp, S. Filopodia-based Wnt transport during vertebrate tissue patterning. Nat. Commun. 2015, 6, 5846. [Google Scholar] [CrossRef] [PubMed]
- Sagar; Pröls, F.; Wiegreffe, C.; Scaal, M. Communication between distant epithelial cells by filopodia-like protrusions during embryonic development. Development 2015, 142, 665. [Google Scholar] [CrossRef] [PubMed]
- Baeg, G.H.; Lin, X.; Khare, N.; Baumgartner, S.; Perrimon, N. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 2001, 128, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yan, D.; Belenkaya, T.Y.; Lin, X. Drosophila glypicans Dally and Dally-like shape the extracellular Wingless morphogen gradient in the wing disc. Development 2005, 132, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wu, Y.; Feng, Y.; Lin, S.C.; Lin, X. The core protein of glypican Dally-like determines its biphasic activity in wingless morphogen signaling. Dev. Cell 2009, 17, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Gallet, A.; Staccini-Lavenant, L.; Therond, P.P. Cellular trafficking of the glypican Dally-like is required for full-strength Hedgehog signaling and wingless transcytosis. Dev. Cell 2008, 14, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Sexton, T.R.; Dejima, K.; Perry, D.W.; Takemura, M.; Kobayashi, S.; Nakato, H.; Harrison, D.A. Glypicans regulate JAK/STAT signaling and distribution of the Unpaired morphogen. Development 2012, 139, 4162–4171. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kornberg, T.B. Cells must express components of the planar cell polarity system and extracellular matrix to support cytonemes. eLife 2016, 5, e18979. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Fly Gene Name | Vertebrate Family | Function |
|---|---|---|
| wingless (wg)/Wnt1 Wnt2 Wnt4 Wnt5 Wnt6 Wnt8/WntD Wnt10 | Wnt | Ligand (Positive regulator) |
| frizzled (fz) frizzled 2 (fz2) frizzled3 (fz3) frizzled 4 (fz4) | Frizzled (Fz) | Receptor (Positive regulator) |
| arrow (arr) | Low-density lipoprotein receptor-related protein 5/6 (LRP5/6) | Co-receptor (Positive regulator) |
| dishevelled (dsh) | Dishevelled (Dsh or Dvl) | Signal transduction (Positive regulator) |
| APC-like (Apc) Adenomatous polyposis coli 2 (Apc2) | Adenomatous Polyposis Coli (APC) | Signal transduction (Negative regulator—component of the ‘destruction complex’) |
| Axin (Axn) | Axin (Axn) | Signal transduction (Negative regulator—component of the ‘destruction complex’) |
| shaggy (sgg) | Glycogen Synthase Kinase 3β (GSK3β) | Signal transduction (Negative regulator—component of the ‘destruction complex’) |
| armadillo (arm) | β-Catenin (β-Cat) | Transcriptional co-activator (Positive regulator)/cell-adhesion molecule |
| pangolin (pan) | T-cell factor ( TCF) | Transcriptional co-activator (Positive regulator) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waghmare, I.; Page-McCaw, A. Wnt Signaling in Stem Cell Maintenance and Differentiation in the Drosophila Germarium. Genes 2018, 9, 127. https://doi.org/10.3390/genes9030127
Waghmare I, Page-McCaw A. Wnt Signaling in Stem Cell Maintenance and Differentiation in the Drosophila Germarium. Genes. 2018; 9(3):127. https://doi.org/10.3390/genes9030127
Chicago/Turabian StyleWaghmare, Indrayani, and Andrea Page-McCaw. 2018. "Wnt Signaling in Stem Cell Maintenance and Differentiation in the Drosophila Germarium" Genes 9, no. 3: 127. https://doi.org/10.3390/genes9030127
APA StyleWaghmare, I., & Page-McCaw, A. (2018). Wnt Signaling in Stem Cell Maintenance and Differentiation in the Drosophila Germarium. Genes, 9(3), 127. https://doi.org/10.3390/genes9030127