Divergent Roles of RPA Homologs of the Model Archaeon Halobacterium salinarum in Survival of DNA Damage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
2.2. Transformation
2.3. Plasmids and Strain Construction
2.4. DNA Damage Treatments
2.4.1. Ionizing Radiation
2.4.2. Ultraviolet Irradiation
2.4.3. Mitomycin C Treatment
2.5. Reverse Transcription—Polymerase Chain Reaction and Transcript Analysis
2.6. Quantitative PCR
2.7. Statistical Analysis
3. Results
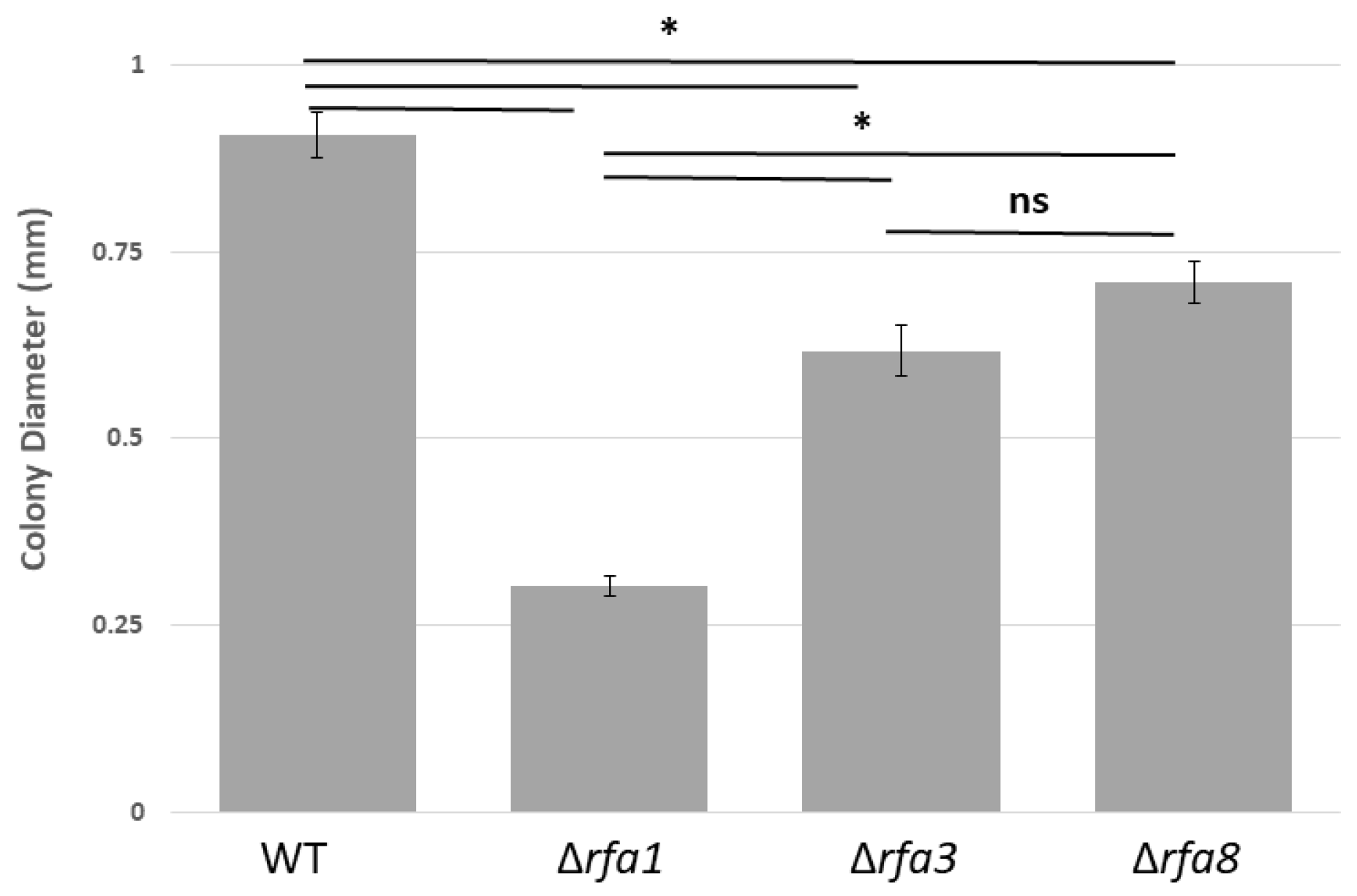
3.1. Effect of Replication Protein A Deletions on Growth
rfa1, rfa3 and rfa8 Deletions Affect Growth
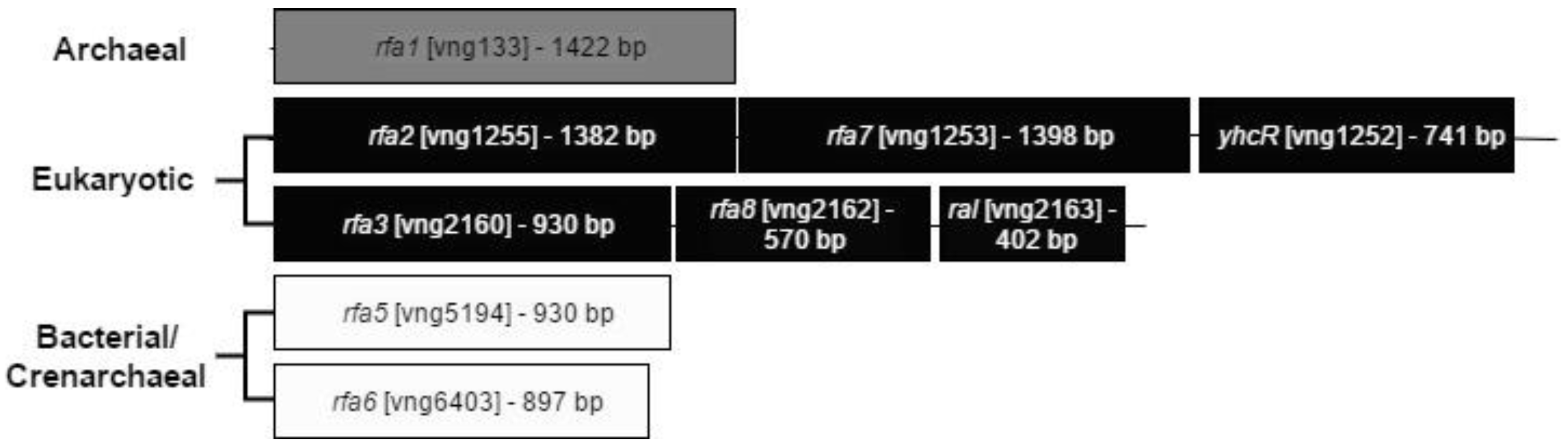
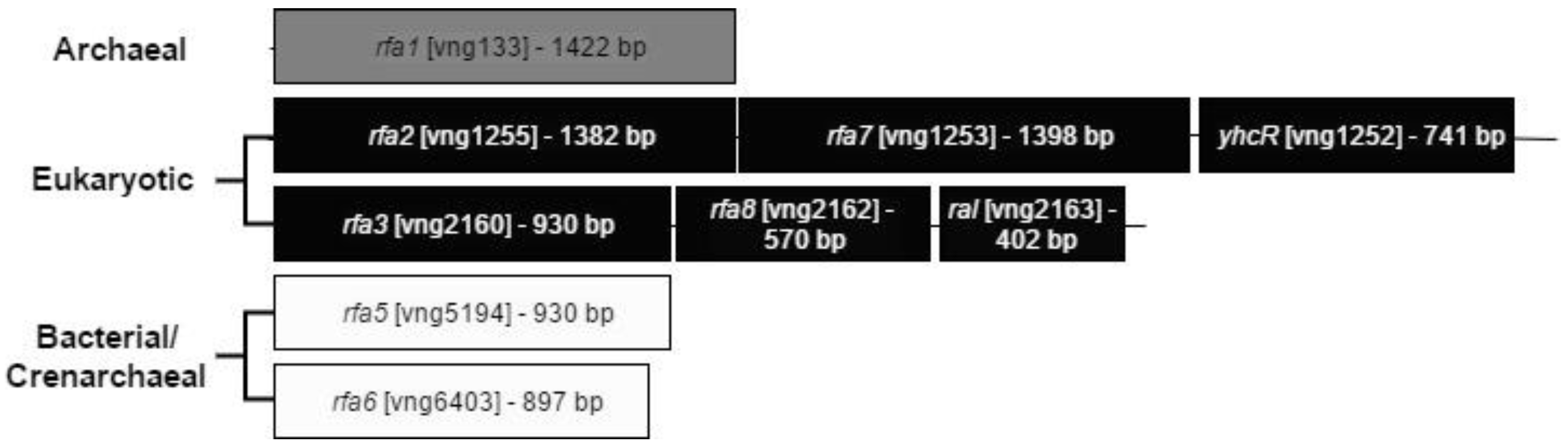
3.2. Deletion of rfa2, rfa7, and yhcR
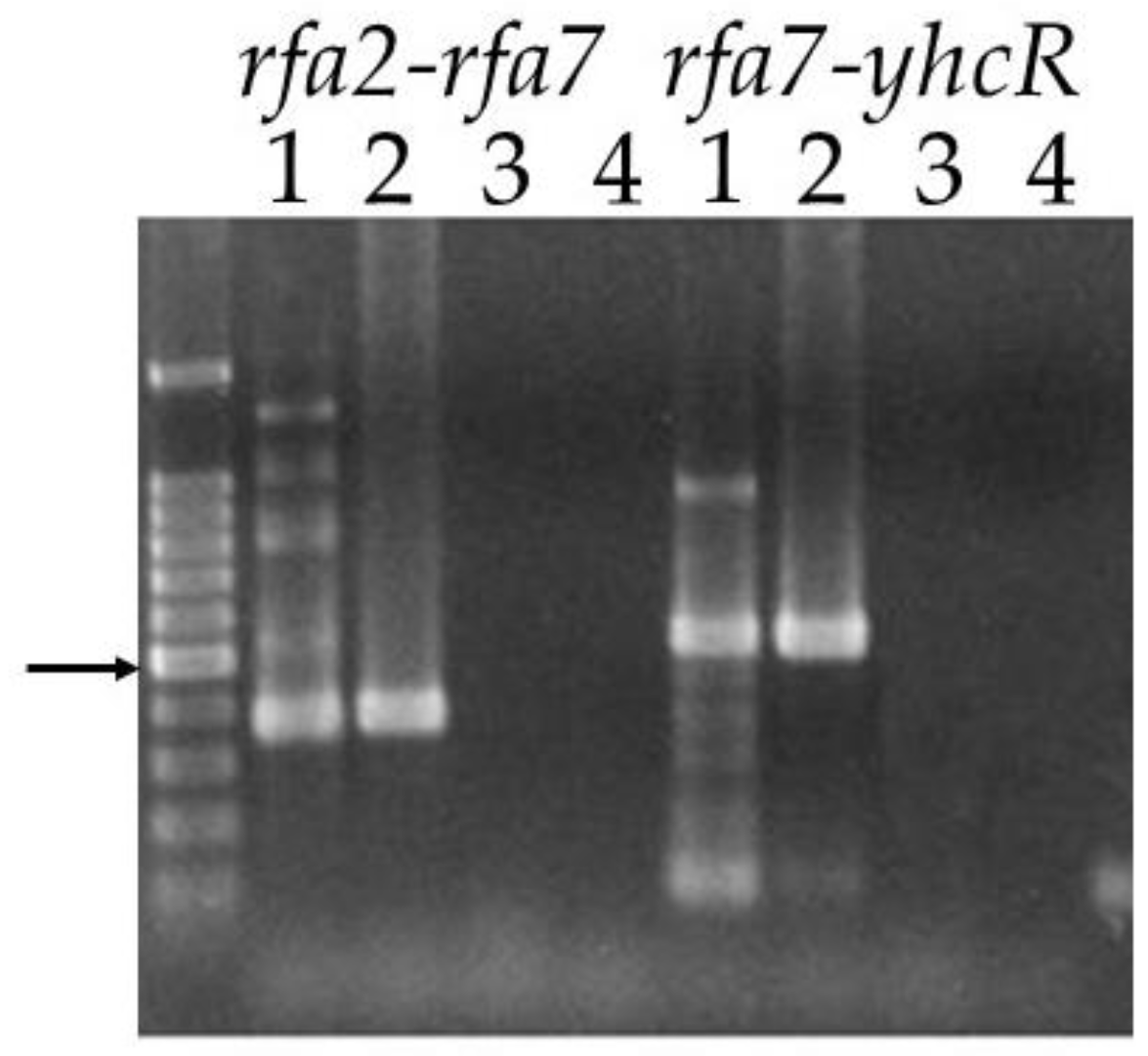
3.2.1. Co-Transcription of the rfa2, rfa7, and yhcR Genes
3.2.2. rfa2 May Be Essential
3.3. Effect of Deletions on Survival to Ionizing Radiation, Ultraviolet C, and Mitomycin C
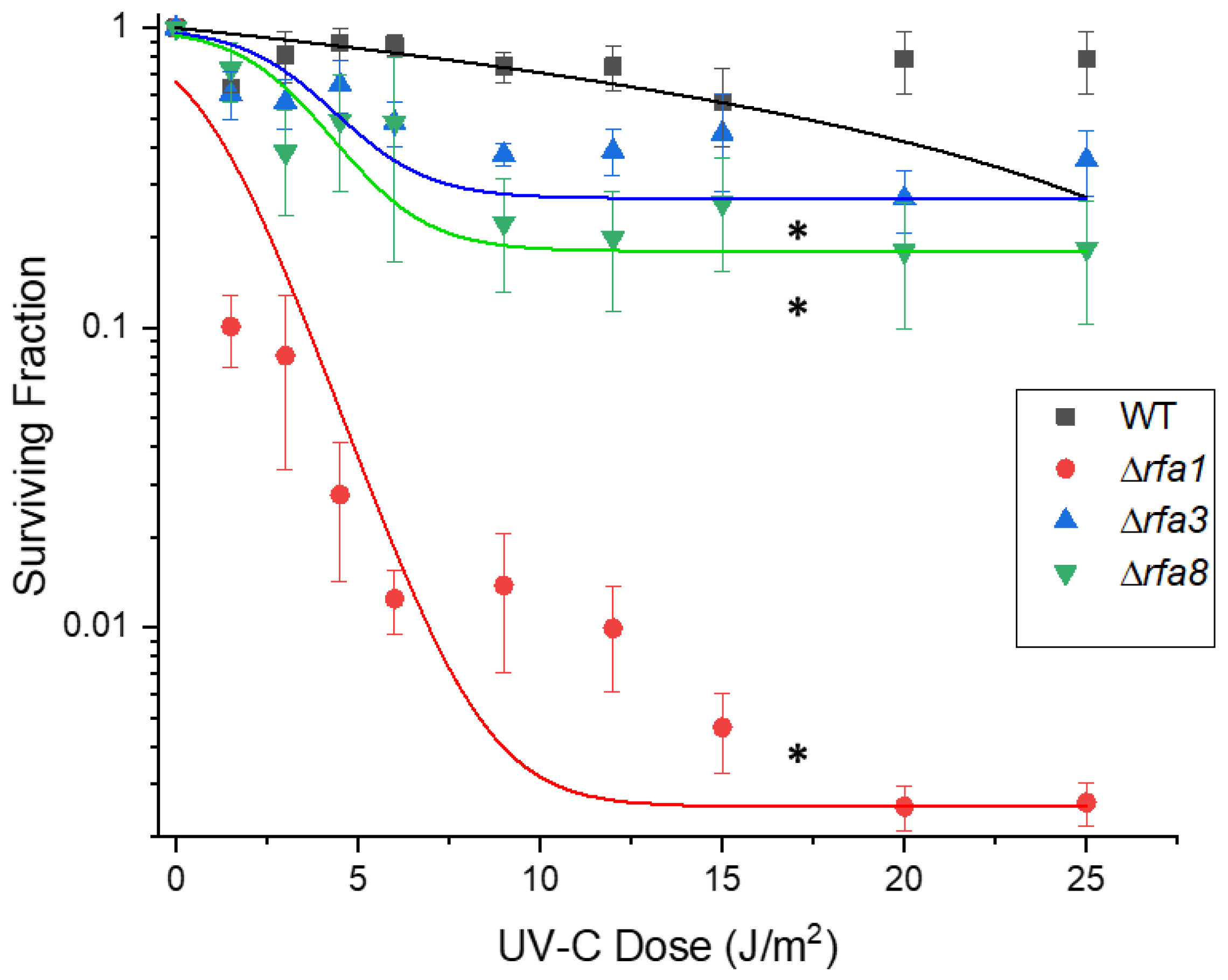
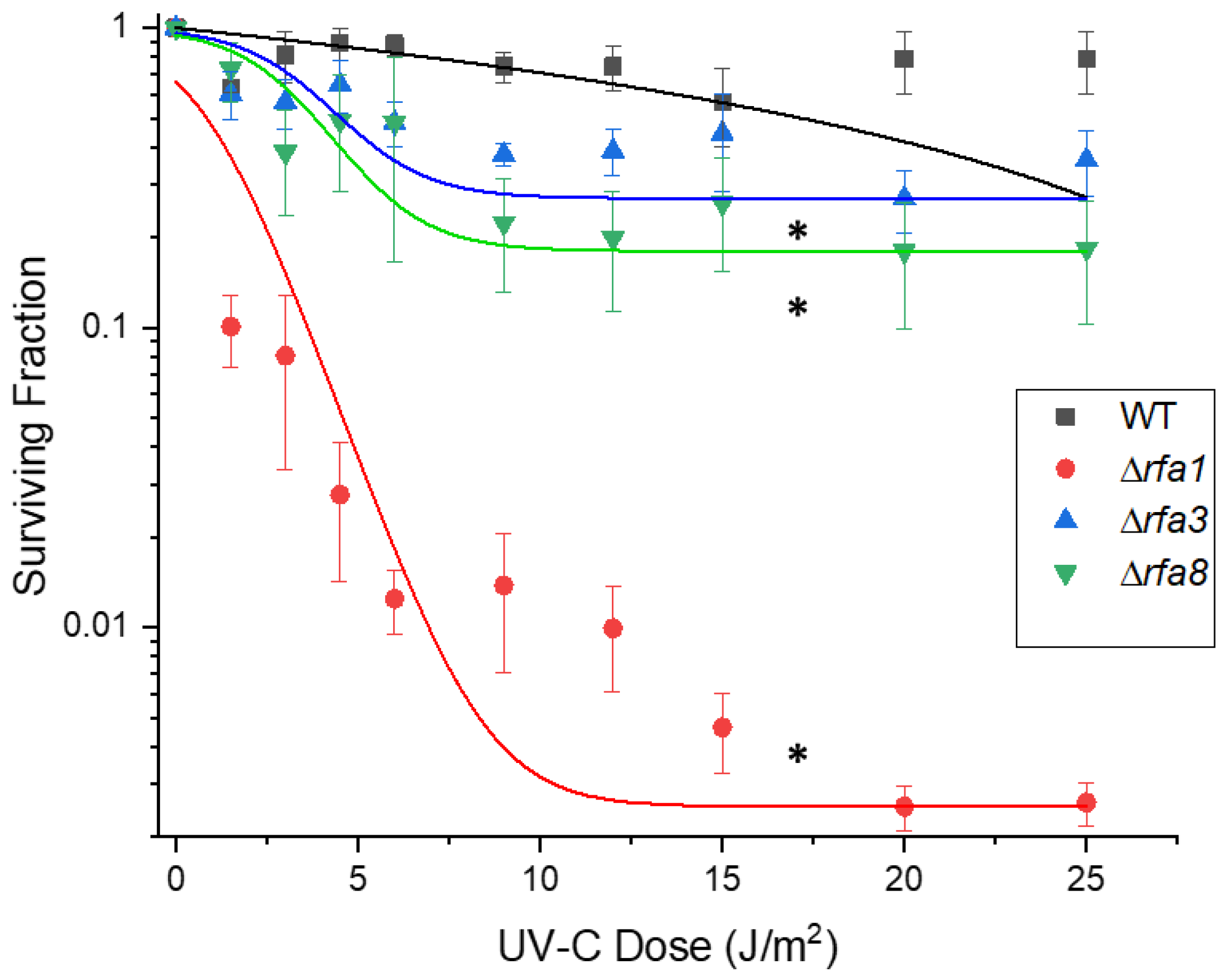
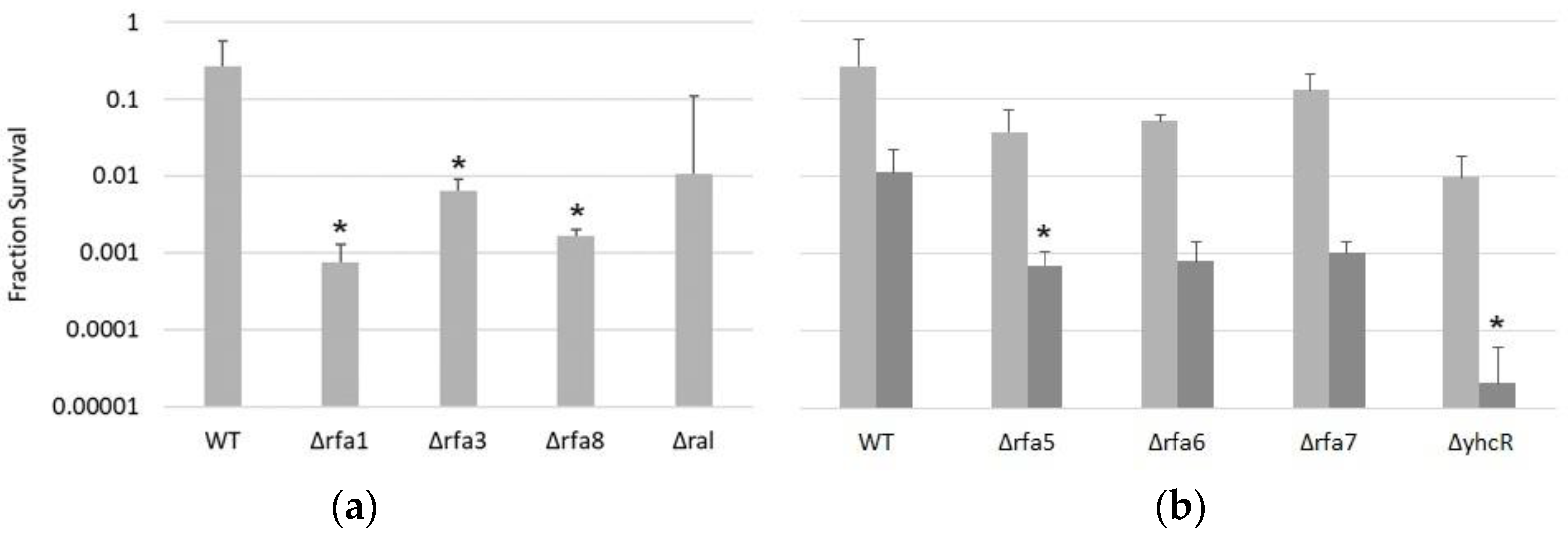
3.3.1. Δrfa3, Δrfa8, and Δrfa1 Confer Ionizing Radiation Sensitivity
3.3.2. Ultraviolet C Survival is Similar to Ionizing Radiation
3.3.3. Δrfa5, Δrfa6, Δrfa7, and ΔyhcR Strains Are Also Sensitive to Mitomycin C
3.4. Expression Levels of Replication Protein A Genes
3.4.1. Replication Protein A Genes are Expressed Under Normal Growing Conditions
3.4.2. DNA-Damaging Treatment Results in Differential Expression
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Awate, S.; Brosh, R.M., Jr. Interactive roles of DNA helicases and translocases with the single-stranded DNA binding protein RPA in nucleic acid metabolism. Int. J. Mol. Sci. 2017, 18, 1233. [Google Scholar] [CrossRef] [PubMed]
- Komori, K.; Ishino, Y. Replication Protein A in Pyrococcus furiosus is involved in homologous DNA recombination. J. Biol. Chem. 2001, 276, 25654–25660. [Google Scholar] [CrossRef] [PubMed]
- Shereda, R.D.; Bernstein, D.A.; Keck, J.L. A central role for SSB in Escherichia coli RecQ DNA helicase function. J. Biol. Chem. 2007, 282, 19247–19258. [Google Scholar] [CrossRef] [PubMed]
- Shereda, R.D.; Kozlov, A.G.; Lohman, T.M.; Cox, M.M.; Keck, J.L. SSB as an organizer/mobilizer of genome maintenance complexes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 289–318. [Google Scholar] [CrossRef] [PubMed]
- Plate, I.; Hallwyl, S.C.; Shi, I.; Krejci, L.; Muller, C.; Albertsen, L.; Sung, P.; Mortensen, U.H. Interaction with RPA is necessary for Rad52 repair center formation and for its mediator activity. J. Biol. Chem. 2008, 283, 29077–29085. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.W.; Loo, D.; Paquet, N.; O’Byrne, K.J.; Richard, D.J. Novel insight into the composition of human single-stranded DNA-binding protein 1 (hSSB1)-containing protein complexes. BMC Mol. Biol. 2016, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, L.J.; Sriratana, P.; Coleman, K.; Ha, T.; Spies, M.; Cann, I.K. Engineering of functional Replication Protein A homologs based on insights into the evolution of oligonucleotide/oligosaccharide-binding folds. J. Bacteriol. 2008, 190, 5766–5780. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.B.; McKinney, M.C.; Guzman, C.E.; Sriratana, B.; Fitz-Gibbon, S.; Ha, T.; Cann, I.K. The euryarchaeota, nature’s medium for engineering of single-stranded DNA-binding proteins. J. Biol. Chem. 2005, 280, 15325–15339. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.J.; Bolderson, E.; Cubeddu, L.; Wadsworth, R.I.; Savage, K.; Sharma, G.G.; Nicolette, M.L.; Tsvetanov, S.; McIlwraith, M.J.; Pandita, R.K.; et al. Single-stranded DNA-binding protein hSSB1 is critical for genomic stability. Nature 2008, 453, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Norais, C.A.; Chitteni-Pattu, S.; Wood, E.A.; Inman, R.B.; Cox, M.M. Ddrb protein, an alternative Deinococcus radiodurans SSB induced by ionizing radiation. J. Biol. Chem. 2009, 284, 21402–21411. [Google Scholar] [CrossRef] [PubMed]
- Wold, M.S. Replication Protein A: A heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu. Rev. Biochem. 1997, 66, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.B.; Murphy, M.C.; White, B.A.; Mackie, R.I.; Ha, T.; Cann, I.K. Functional analysis of multiple single-stranded DNA-binding proteins from Methanosarcina acetivorans and their effects on DNA synthesis by DNA polymerase bi. J. Biol. Chem. 2004, 279, 6315–6326. [Google Scholar] [CrossRef] [PubMed]
- Kamekura, M. Diversity of extremely halophilic bacteria. Extremophiles 1998, 2, 289–295. [Google Scholar] [CrossRef] [PubMed]
- DeVeaux, L.C.; Muller, J.A.; Smith, J.; Petrisko, J.; Wells, D.P.; DasSarma, S. Extremely radiation-resistant mutants of a halophilic archaeon with increased single-stranded DNA-binding protein (RPA) gene expression. Radiat. Res. 2007, 168, 507–514. [Google Scholar] [PubMed]
- Webb, K.M.; Yu, J.; Robinson, C.K.; Noboru, T.; Lee, Y.C.; DiRuggiero, J. Effects of intracellular Mn on the radiation resistance of the halophilic archaeon Halobacterium salinarum. Extremophiles 2013, 17, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Karan, R.; DasSarma, P.; Balcer-Kubiczek, E.; Weng, R.R.; Liao, C.C.; Goodlett, D.R.; Ng, W.V.; Dassarma, S. Bioengineering radioresistance by overproduction of RPA, a mammalian-type single-stranded DNA-binding protein, in a halophilic archaeon. Appl. Microbiol. Biotechnol. 2014, 98, 1737–1747. [Google Scholar] [CrossRef] [PubMed]
- Skowyra, A.; MacNeill, S.A. Identification of essential and non-essential single-stranded DNA-binding proteins in a model archaeal organism. Nucleic Acids Res. 2012, 40, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Capes, M.D.; DasSarma, P.; DasSarma, S. The core and unique proteins of haloarchaea. BMC Genom. 2012, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- Grey, V.L.; Fitt, P.S. An improved synthetic growth medium for Halobacterium cutirubrum. Can. J. Microbiol. 1976, 22, 440–442. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, S.; Fleischmann, E. Halophiles. In Archaea: A Laboratory Manual; Robb, F.T., Place, A.R., Eds.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1995; p. 280. [Google Scholar]
- Miller, J. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: New York, NY, USA, 1972. [Google Scholar]
- Gygli, P.E.; DeVeaux, L.C. Adaptation of the Halobacterium salinarum ssp. NRC-1 gene deletion system for modification of chromosomal loci. J. Microbiol. Methods 2014, 99, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Berman, M.L.; Enquist, L.W. Experiments with Gene Fusions; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1984; p. 303. [Google Scholar]
- Peck, R.F.; DasSarma, S.; Krebs, M.P. Homologous gene knockout in the archaeon Halobacterium salinarum with ura3 as a counterselectable marker. Mol. Microbiol. 2000, 35, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, J.S.; DeVeaux, L.C. The essential role of the Deinococcus radiodurans SSB gene in cell survival and radiation tolerance. PLoS ONE 2013, 8, e71651. [Google Scholar] [CrossRef] [PubMed]
- Boubriak, I.; Ng, W.L.; DasSarma, P.; DasSarma, S.; Crowley, D.J.; McCready, S.J. Transcriptional responses to biologically relevant doses of UV-B radiation in the model archaeon, Halobacterium sp. NRC-1. Saline Syst. 2008, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671. [Google Scholar] [CrossRef]
- Ng, W.V.; Kennedy, S.P.; Mahairas, G.G.; Berquist, B.; Pan, M.; Shukla, H.D.; Lasky, S.R.; Baliga, N.S.; Thorsson, V.; Sbrogna, J.; et al. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 2000, 97, 12176–12181. [Google Scholar] [CrossRef] [PubMed]
- Gygli, P.E.; Prajapati, S.; DeVeaux, L.C.; DasSarma, S.; DasSarma, P.; Mestari, M.A.; Wells, D.P. Resistance of the extreme halophile Halobacterium sp. NRC-1 to multiple stresses. AIP Conf. Proc. 2009, 1099, 993–996. [Google Scholar]
- McCready, S.; Marcello, L. Repair of UV damage in Halobacterium salinarum. Biochem. Soc. Trans. 2003, 31, 694–698. [Google Scholar] [CrossRef] [PubMed]
- McCready, S.; Muller, J.A.; Boubriak, I.; Berquist, B.R.; Ng, W.L.; DasSarma, S. UV irradiation induces homologous recombination genes in the model archaeon, Halobacterium sp. NRC-1. Saline Syst. 2005, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, A.; Van, P.T.; Busch, C.R.; Robinson, C.K.; Pan, M.; Pang, W.L.; Reiss, D.J.; DiRuggiero, J.; Baliga, N.S. Coordination of frontline defense mechanisms under severe oxidative stress. Mol. Syst. Biol. 2010, 6, 393. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K.; Kish, A.; Pan, M.; Kaur, A.; Reiss, D.J.; King, N.; Hohmann, L.; DiRuggiero, J.; Baliga, N.S. An integrated systems approach for understanding cellular responses to gamma radiation. Mol. Syst. Biol. 2006, 2, 47. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Earl, A.M.; Howell, H.A.; Park, M.J.; Eisen, J.A.; Peterson, S.N.; Battista, J.R. Analysis of Deinococcus radiodurans’s transcriptional response to ionizing radiation and desiccation reveals novel proteins that contribute to extreme radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.A.; Smith, P.; Shuman, S. Structure of bacterial Ligd 3′-phosphoesterase unveils a DNA repair superfamily. Proc. Natl. Acad. Sci. USA 2010, 107, 12822–12827. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hbt. salinarum | Hfx. volcanii | ||||||
|---|---|---|---|---|---|---|---|
| RPA/SSB Type | Protein | Size (aa) | Protein | Size (aa) | Shared Features | % Identity | E Value |
| Eukaryotic | Rfa2 | 460 | RpaA1 | 427 | two OB-folds/ one Zinc Finger | 64 | 1 × 10−179 |
| Rfa3 | 473 | RpaB1 | 311 | one OB-fold/ one Zinc Finger | 67 | 2 × 10−150 | |
| Rfa7 | 465 | RpaA2 | 623 | COG3390 domain; uncharacterized | 65 | 8 × 10−93 | |
| Rfa8 | 190 | RpaB2 | 196 | COG3390 domain; uncharacterized | 75 | 2 × 10−101 | |
| YhcR | 247 | HVO_1336 | 260 | Phosphoesterase | 57 | 1 × 10−78 | |
| Ral | 134 | HVO_0290 | 137 | Unique to Haloarchaea [18] | 66 | 7 × 10−65 | |
| Crenarchaeal/Bacterial | Rfa5 Rfa6 | 330 299 | HVO_A0019 | 302 | 1 OB fold | 88 * 78 | 0.0 * 3 × 10−173 |
| HVO_A0374 | 301 | 1 OB fold | 88 83 | 0.0 0.0 | |||
| HVO_A0409 | 287 | 1 OB fold | 73 71 | 8 × 10−155 4 × 10−150 | |||
| Euryarchaeal | Rfa1 | 474 | RpaC | 483 | 3 OB folds; Essential in Hfx. volcanii | 60 | 0.0 |
| Strain | Genotype | Source |
|---|---|---|
| NRC-1 | Wild-type | Lab stock |
| LH5 | Radiation-resistant mutant of NRC-1 | [14] |
| LH101 | As NRC-1 but Δura3 | This study |
| LH102 | As LH5 but Δura3 | This study |
| LH110 | As LH101 but Δrfa5 | This study |
| LH128 | As LH101 but ΔyhcR | This study |
| LH134 | As LH101 but Δrfa3 | This study |
| LH136 | As LH101 but Δrfa8 | This study |
| LH138 | As LH101 but Δral | This study |
| LH140 | As LH101 but Δrfa1 | This study |
| LH142 | As LH101 but Δrfa6 | This study |
| LH154 | As LH102 but Δrfa2 | This study |
| LH166 | As LH101 but Δrfa7 | This study |
| Strain | (rfa2)/(eef2) | Relative Expression |
|---|---|---|
| LH101 | 0.001 | 1.0 |
| LH154 | 0.003 | 0.32 |
| Target | Quantity (Copies/µL) | Standard Deviation |
|---|---|---|
| rfa1 (‡,¥) | 3.22 × 107 | ±1.48 × 107 |
| rfa2 (‡,¥) | 2.18 × 106 | ±2.08 × 105 |
| rfa3 (‡) | 1.8 × 108 | ±8.83 × 107 |
| rfa5.6 (‡,¥) | 4.88 × 106 | ±1.07 × 106 |
| rfa7 (‡,¥) | 1.10 × 107 | ±1.87 × 106 |
| rfa8 (‡) | 9.70 × 107 | ±1.76 × 107 |
| eef2 | 2.69 × 108 | ±5.9 × 107 |
| Gene | Treatment | Copies/µL | (Target)/(eef2) | Relative Change | p-Value |
|---|---|---|---|---|---|
| rfa1 | Untreated | 3.22 × 107 | 0.12 | 1.0 | |
| UV-C | 3.04 × 107 | 0.10 | 0.81 | 0.76 | |
| MMC | 2.67 × 107 | 0.08 | 0.65 | 0.90 | |
| rfa2 | Untreated | 2.18 × 106 | 0.01 | 1.0 | |
| UV-C | 1.97 × 107 | 0.06 | 7.74 | 0.001 | |
| MMC | 1.93 × 107 | 0.06 | 7.01 | 0.001 | |
| rfa7 | Untreated | 1.10 × 107 | 0.04 | 1.0 | |
| UV-C | 1.32 × 107 | 0.04 | 1.03 | 0.65 | |
| MMC | 9.44 × 106 | 0.03 | 0.68 | 0.92 | |
| rfa3 | Untreated | 1.79 × 108 | 0.67 | 1.0 | |
| UV-C | 3.57 × 108 | 1.15 | 1.71 | 0.19 | |
| MMC | 2.54 × 108 | 0.75 | 1.120 | 0.47 | |
| rfa8 | Untreated | 9.70 × 107 | 0.36 | 1.0 | |
| UV | 2.27 × 108 | 0.73 | 2.01 | 0.33 | |
| MMC | 2.49 × 108 | 0.73 | 2.03 | 0.03 | |
| rfa5.6 | Untreated | 4.88 × 106 | 0.02 | 1.0 | |
| UV-C | 9.92 × 106 | 0.03 | 1.74 | 0.51 | |
| MMC | 1.70 × 107 | 0.05 | 2.75 | 0.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, J.J.; Gygli, P.E.; McCaskill, J.; DeVeaux, L.C. Divergent Roles of RPA Homologs of the Model Archaeon Halobacterium salinarum in Survival of DNA Damage. Genes 2018, 9, 223. https://doi.org/10.3390/genes9040223
Evans JJ, Gygli PE, McCaskill J, DeVeaux LC. Divergent Roles of RPA Homologs of the Model Archaeon Halobacterium salinarum in Survival of DNA Damage. Genes. 2018; 9(4):223. https://doi.org/10.3390/genes9040223
Chicago/Turabian StyleEvans, Jessica J., Patrick E. Gygli, Julienne McCaskill, and Linda C. DeVeaux. 2018. "Divergent Roles of RPA Homologs of the Model Archaeon Halobacterium salinarum in Survival of DNA Damage" Genes 9, no. 4: 223. https://doi.org/10.3390/genes9040223
APA StyleEvans, J. J., Gygli, P. E., McCaskill, J., & DeVeaux, L. C. (2018). Divergent Roles of RPA Homologs of the Model Archaeon Halobacterium salinarum in Survival of DNA Damage. Genes, 9(4), 223. https://doi.org/10.3390/genes9040223