Chitinase mRNA Levels Determined by QPCR in Crab-Eating Monkey (Macaca fascicularis) Tissues: Species-Specific Expression of Acidic Mammalian Chitinase and Chitotriosidase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA and Complementary DNA Preparation
2.2. Selection of Primer Pairs for qPCR
2.3. Construction of the Monkey Standard DNA and qPCR
2.4. Mouse, Monkey, and Human Stomach and Lung Extracts
2.5. Chitinase Enzymatic Assays
2.6. Statistical Analysis
3. Results
3.1. Characterization of the qPCR System for Detection of Chitinases and Reference Genes in Crab-Eating Monkey Tissues
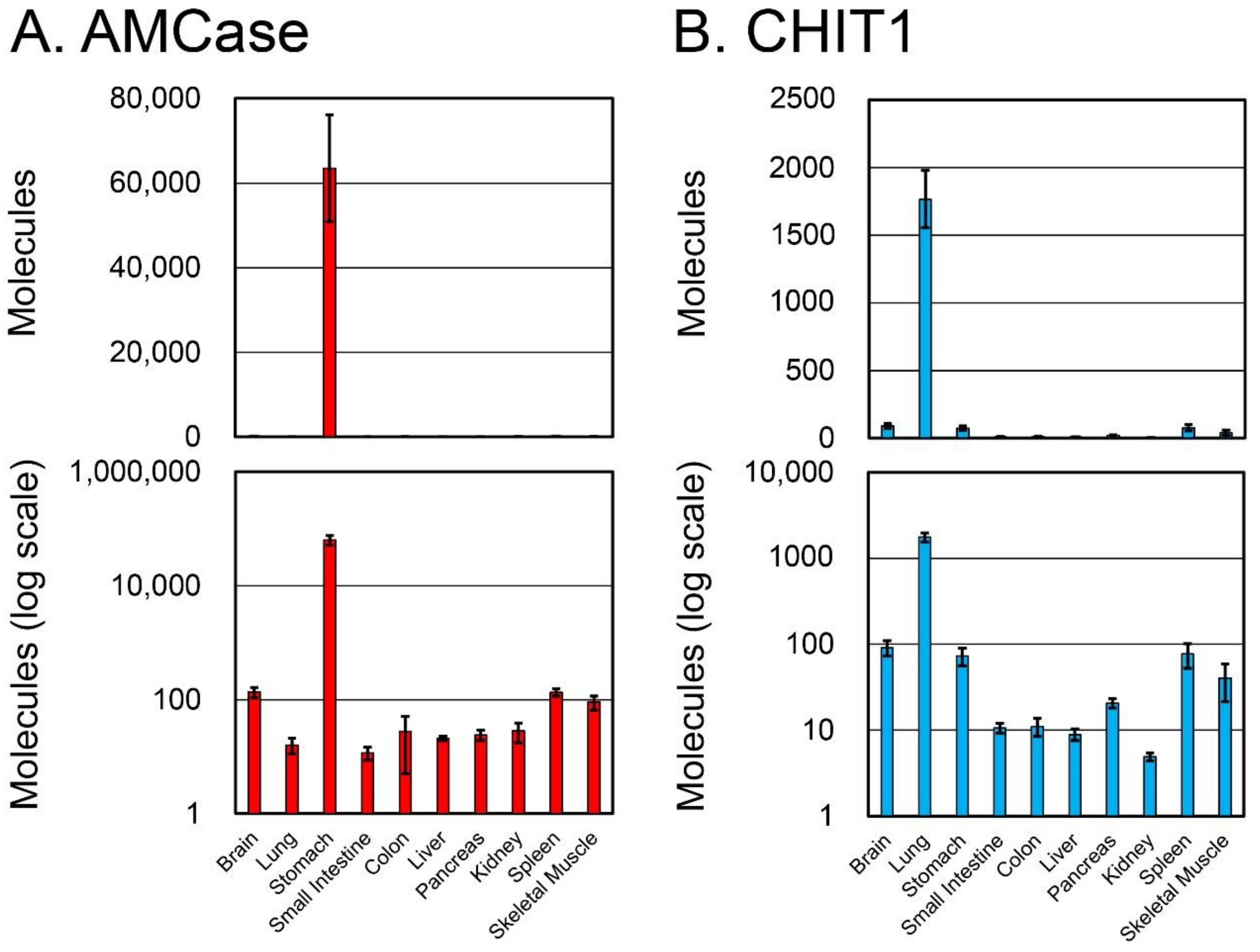
3.2. Expression of AMCase and CHIT1 in Normal Crab-Eating Monkey Tissues
3.3. Expression of AMCase, CHIT1, Pepsinogen C, and GAPDH mRNAs in Normal Crab-Eating Monkey Stomach and Lung Tissues
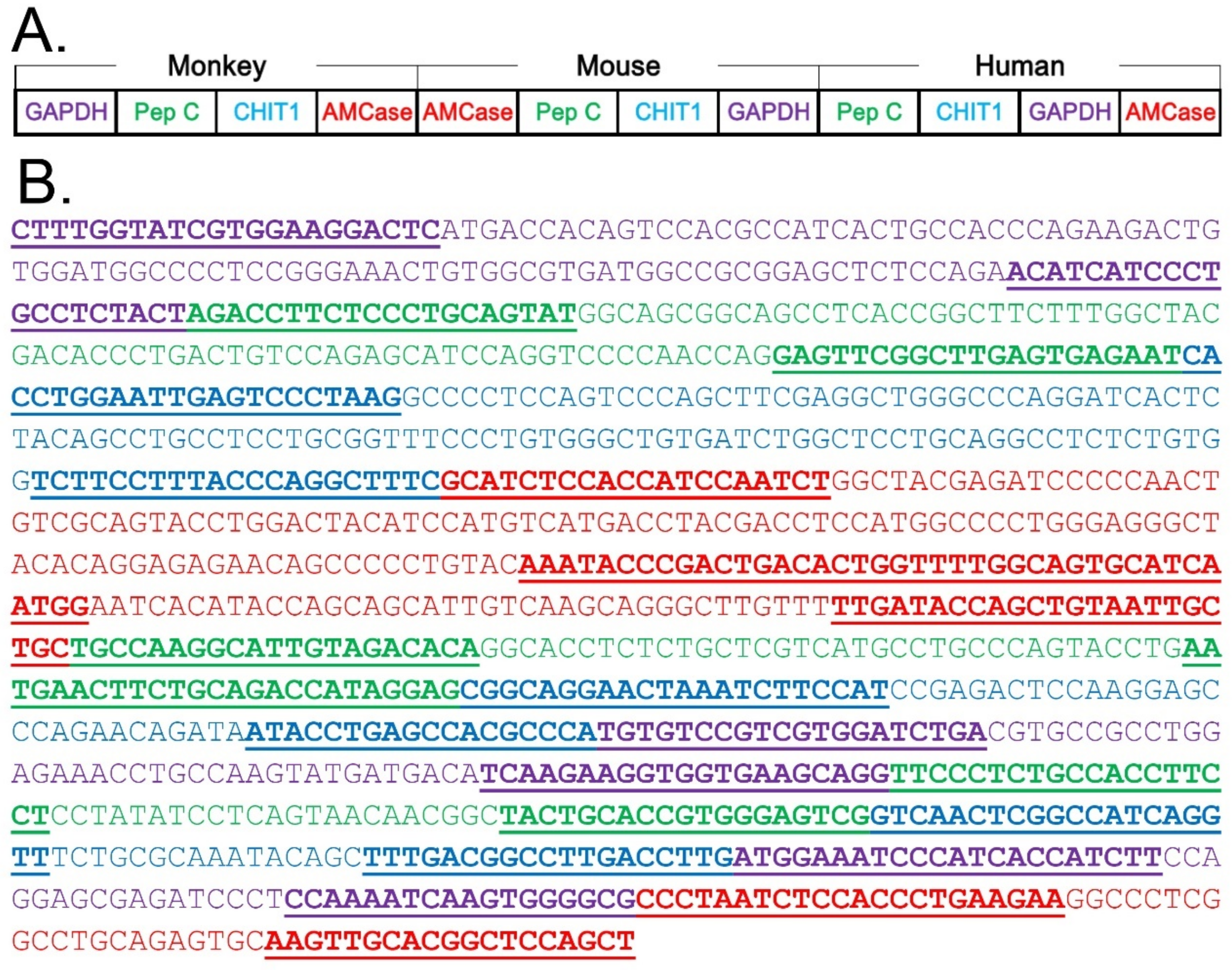
3.4. Characterization of the qPCR System for Chitinase mRNA Level Comparison in Three Species Using a Monkey–Mouse–Human Hybrid DNA
3.5. Comparison of Chitinase and Reference Gene mRNA Levels between Normal Mouse, Monkey, and Human Stomach Tissues
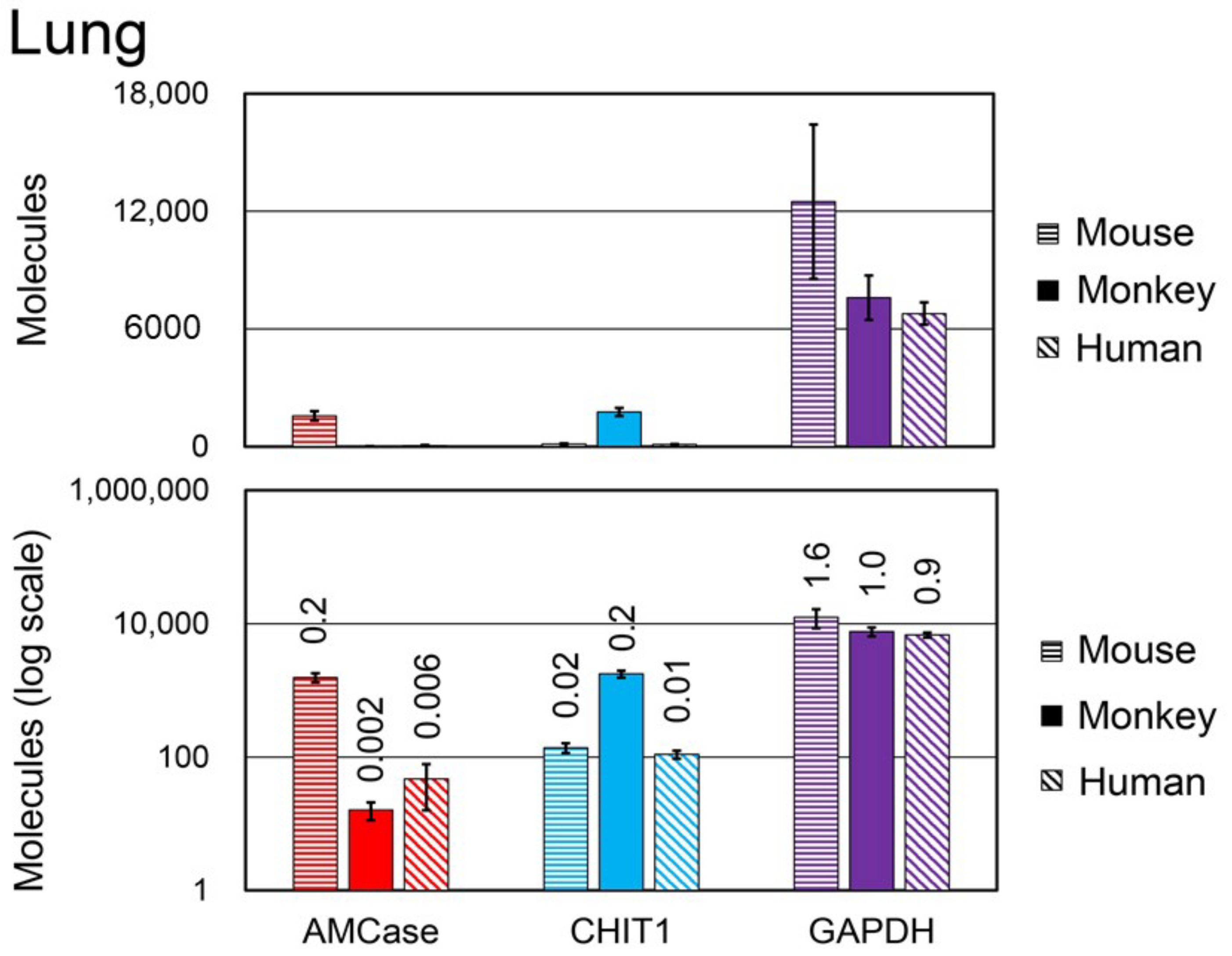
3.6. Comparison of Chitinase and Reference Gene mRNA Levels between Normal Mouse, Monkey, and Human Lung Tissues
3.7. Chitinase Levels and Activity in Mouse, Monkey, and Human Stomach and Lung
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Khoushab, F.; Yamabhai, M. Chitin research revisited. Mar. Drugs 2010, 8, 1988–2012. [Google Scholar] [CrossRef] [PubMed]
- Koch, B.E.; Stougaard, J.; Spaink, H.P. Keeping track of the growing number of biological functions of chitin and its interaction partners in biomedical research. Glycobiology 2015, 25, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Bueter, C.L.; Specht, C.A.; Levitz, S.M. Innate sensing of chitin and chitosan. PLoS Pathog. 2013, 9, e1003080. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.G.; Da Silva, C.A.; Dela Cruz, C.S.; Ahangari, F.; Ma, B.; Kang, M.J.; He, C.H.; Takyar, S.; Elias, J.A. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu. Rev. Physiol. 2011, 73, 479–501. [Google Scholar] [CrossRef] [PubMed]
- Hollak, C.E.; van Weely, S.; van Oers, M.H.; Aerts, J.M. Marked elevation of plasma chitotriosidase activity. A novel hallmark of Gaucher disease. J. Clin. Investig. 1994, 93, 1288–1292. [Google Scholar] [CrossRef] [PubMed]
- Renkema, G.H.; Boot, R.G.; Muijsers, A.O.; Donker-Koopman, W.E.; Aerts, J.M. Purification and characterization of human chitotriosidase, a novel member of the chitinase family of proteins. J. Biol. Chem. 1995, 270, 2198–2202. [Google Scholar] [CrossRef] [PubMed]
- Boot, R.G.; Renkema, G.H.; Strijland, A.; van Zonneveld, A.J.; Aerts, J.M. Cloning of a cDNA encoding chitotriosidase, a human chitinase produced by macrophages. J. Biol. Chem. 1995, 270, 26252–26256. [Google Scholar] [CrossRef] [PubMed]
- Boot, R.G.; Blommaart, E.F.; Swart, E.; Ghauharali-van der Vlugt, K.; Bijl, N.; Moe, C.; Place, A.; Aerts, J.M. Identification of a novel acidic mammalian chitinase distinct from chitotriosidase. J. Biol. Chem. 2001, 276, 6770–6778. [Google Scholar] [CrossRef] [PubMed]
- Boot, R.G.; Bussink, A.P.; Verhoek, M.; de Boer, P.A.; Moorman, A.F.; Aerts, J.M. Marked differences in tissue-specific expression of chitinases in mouse and man. J. Histochem. Cytochem. 2005, 53, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Letuve, S.; Kozhich, A.; Humbles, A.; Brewah, Y.; Dombret, M.C.; Grandsaigne, M.; Adle, H.; Kolbeck, R.; Aubier, M.; Coyle, A.J.; et al. Lung chitinolytic activity and chitotriosidase are elevated in chronic obstructive pulmonary disease and contribute to lung inflammation. Am. J. Pathol. 2010, 176, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Watabe-Rudolph, M.; Song, Z.; Lausser, L.; Schnack, C.; Begus-Nahrmann, Y.; Scheithauer, M.O.; Rettinger, G.; Otto, M.; Tumani, H.; Thal, D.R.; et al. Chitinase enzyme activity in CSF is a powerful biomarker of Alzheimer disease. Neurology 2012, 78, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Seibold, M.A.; Donnelly, S.; Solon, M.; Innes, A.; Woodruff, P.G.; Boot, R.G.; Burchard, E.G.; Fahy, J.V. Chitotriosidase is the primary active chitinase in the human lung and is modulated by genotype and smoking habit. J. Allergy Clin. Immunol. 2008, 122, 944–950.e3. [Google Scholar] [CrossRef] [PubMed]
- Artieda, M.; Cenarro, A.; Ganan, A.; Jerico, I.; Gonzalvo, C.; Casado, J.M.; Vitoria, I.; Puzo, J.; Pocovi, M.; Civeira, F. Serum chitotriosidase activity is increased in subjects with atherosclerosis disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Sonmez, A.; Haymana, C.; Tapan, S.; Safer, U.; Celebi, G.; Ozturk, O.; Genc, H.; Dogru, T.; Tasci, I.; Erdem, G.; et al. Chitotriosidase activity predicts endothelial dysfunction in type-2 diabetes mellitus. Endocrine 2010, 37, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Livnat, G.; Bar-Yoseph, R.; Mory, A.; Dagan, E.; Elias, N.; Gershoni, R.; Bentur, L. Duplication in CHIT1 gene and the risk for Aspergillus lung disease in CF patients. Pediatr. Pulmonol. 2014, 49, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.J.; Morroll, S.; Tighe, P.; Gotz, F.; Falcone, F.H. Human chitotriosidase is expressed in the eye and lacrimal gland and has an antimicrobial spectrum different from lysozyme. Microbes Infect. 2008, 10, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zheng, T.; Homer, R.J.; Kim, Y.K.; Chen, N.Y.; Cohn, L.; Hamid, Q.; Elias, J.A. Acidic mammalian chitinase in asthmatic Th2 inflammation and IL-13 pathway activation. Science 2004, 304, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Reese, T.A.; Liang, H.E.; Tager, A.M.; Luster, A.D.; Van Rooijen, N.; Voehringer, D.; Locksley, R.M. Chitin induces accumulation in tissue of innate immune cells associated with allergy. Nature 2007, 447, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Bierbaum, S.; Nickel, R.; Koch, A.; Lau, S.; Deichmann, K.A.; Wahn, U.; Superti-Furga, A.; Heinzmann, A. Polymorphisms and haplotypes of acid mammalian chitinase are associated with bronchial asthma. Am. J. Respir. Crit. Care Med. 2005, 172, 1505–1509. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.; Batra, J.; Das, S.; Sharma, S.K.; Ghosh, B. Genetic association of acidic mammalian chitinase with atopic asthma and serum total IgE levels. J. Allergy Clin. Immunol. 2008, 122, 202–208.e7. [Google Scholar] [CrossRef] [PubMed]
- Seibold, M.A.; Reese, T.A.; Choudhry, S.; Salam, M.T.; Beckman, K.; Eng, C.; Atakilit, A.; Meade, K.; Lenoir, M.; Watson, H.G.; et al. Differential enzymatic activity of common haplotypic versions of the human acidic mammalian chitinase protein. J. Biol. Chem. 2009, 284, 19650–19658. [Google Scholar] [CrossRef] [PubMed]
- Okawa, K.; Ohno, M.; Kashimura, A.; Kimura, M.; Kobayashi, Y.; Sakaguchi, M.; Sugahara, Y.; Kamaya, M.; Kino, Y.; Bauer, P.O.; et al. Loss and gain of human acidic mammalian chitinase activity by nonsynonymous SNPs. Mol. Biol. Evol. 2016, 33, 3183–3193. [Google Scholar] [CrossRef] [PubMed]
- Van Dyken, S.J.; Liang, H.E.; Naikawadi, R.P.; Woodruff, P.G.; Wolters, P.J.; Erle, D.J.; Locksley, R.M. Spontaneous chitin accumulation in airways and age-related fibrotic lung disease. Cell 2017, 169, 497–509.e13. [Google Scholar] [CrossRef] [PubMed]
- Vannella, K.M.; Ramalingam, T.R.; Hart, K.M.; de Queiroz Prado, R.; Sciurba, J.; Barron, L.; Borthwick, L.A.; Smith, A.D.; Mentink-Kane, M.; White, S.; et al. Acidic chitinase primes the protective immune response to gastrointestinal nematodes. Nat. Immunol. 2016, 17, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Tsuda, K.; Sakaguchi, M.; Sugahara, Y.; Oyama, F. Chitinase mRNA levels by quantitative PCR using the single standard DNA: Acidic mammalian chitinase is a major transcript in the mouse stomach. PLoS ONE 2012, 7, e50381. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Togashi, Y.; Tsuda, K.; Okawa, K.; Kamaya, M.; Sakaguchi, M.; Sugahara, Y.; Oyama, F. Quantification of chitinase mRNA levels in human and mouse tissues by real-time PCR: Species-specific expression of acidic mammalian chitinase in stomach tissues. PLoS ONE 2013, 8, e67399. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Kimura, M.; Miyazaki, H.; Okawa, K.; Onuki, R.; Nemoto, C.; Tabata, E.; Wakita, S.; Kashimura, A.; Sakaguchi, M.; et al. Acidic mammalian chitinase is a proteases-resistant glycosidase in mouse digestive system. Sci. Rep. 2016, 6, 37756. [Google Scholar] [CrossRef] [PubMed]
- Tabata, E.; Kashimura, A.; Wakita, S.; Ohno, M.; Sakaguchi, M.; Sugahara, Y.; Kino, Y.; Matoska, V.; Bauer, P.O.; Oyama, F. Gastric and intestinal proteases resistance of chicken acidic chitinase nominates chitin-containing organisms for alternative whole edible diets for poultry. Sci. Rep. 2017, 7, 6662. [Google Scholar] [CrossRef] [PubMed]
- Tabata, E.; Kashimura, A.; Wakita, S.; Ohno, M.; Sakaguchi, M.; Sugahara, Y.; Imamura, Y.; Seki, S.; Ueda, H.; Matoska, V.; et al. Protease resistance of porcine acidic mammalian chitinase under gastrointestinal conditions implies that chitin-containing organisms can be sustainable dietary resources. Sci. Rep. 2017, 7, 12963. [Google Scholar] [CrossRef] [PubMed]
- Tabata, E.; Kashimura, A.; Kikuchi, A.; Masuda, H.; Miyahara, R.; Hiruma, Y.; Wakita, S.; Ohno, M.; Sakaguchi, M.; Sugahara, Y.; et al. Chitin digestibility is dependent on feeding behaviors, which determine acidic chitinase mRNA levels in mammalian and poultry stomachs. Sci. Rep. 2018, 8, 1461. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.W.; Kim, Y.H.; Park, S.J.; Kim, D.S.; Lee, S.R.; Kim, K.M.; Jeong, K.J.; Kim, J.S.; Song, B.S.; Sim, B.W.; et al. Large-scale transcriptome sequencing and gene analyses in the crab-eating macaque (Macaca fascicularis) for biomedical research. BMC Genom. 2012, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Ilham, K.; Rizaldi; Nurdin, J.; Tsuji, Y. Status of urban populations of the long-tailed macaque (Macaca fascicularis) in west Sumatra, Indonesia. Primates 2017, 58, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Register, T.C.; Carlson, C.S.; Adams, M.R. Serum YKL-40 is associated with osteoarthritis and atherosclerosis in nonhuman primates. Clin. Chem. 2001, 47, 2159–2161. [Google Scholar] [PubMed]
- Krykbaev, R.; Fitz, L.J.; Reddy, P.S.; Winkler, A.; Xuan, D.; Yang, X.; Fleming, M.; Wolf, S.F. Evolutionary and biochemical differences between human and monkey acidic mammalian chitinases. Gene 2010, 452, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The miqe guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Beaulieu, J.F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.; Olsvik, P.A.; Penning, L.C.; Toegel, S. MIQE précis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time PCR experiments. BMC Mol. Biol. 2010, 11, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zainuddin, A.; Chua, K.H.; Abdul Rahim, N.; Makpol, S. Effect of experimental treatment on GAPDH mRNA expression as a housekeeping gene in human diploid fibroblasts. BMC Mol. Biol. 2010, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T. Pepsinogens, progastricsins, and prochymosins: Structure, function, evolution, and development. Cell. Mol. Life Sci. 2002, 59, 288–306. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.T.; Yao, S.; Czerwinski, R.; Fleming, M.; Krykbaev, R.; Xuan, D.; Zhou, H.; Brooks, J.; Fitz, L.; Strand, J.; et al. Kinetic characterization of recombinant human acidic mammalian chitinase. Biochemistry 2006, 45, 4444–4454. [Google Scholar] [CrossRef] [PubMed]
- Strobel, S.; Roswag, A.; Becker, N.I.; Trenczek, T.E.; Encarnacao, J.A. Insectivorous bats digest chitin in the stomach using acidic mammalian chitinase. PLoS ONE 2013, 8, e72770. [Google Scholar] [CrossRef] [PubMed]
- Janiak, M.C.; Chaney, M.E.; Tosi, A.J. Evolution of acidic mammalian chitinase genes (CHIA) is related to body mass and insectivory in primates. Mol. Biol. Evol. 2018, 35, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Shinoda, T.; Nakashima, H.; Watanabe, T.; Yokohama, S. Characteristics of the gastric pH profiles of unfed and fed cynomolgus monkeys as pharmaceutical product development subjects. Biopharm. Drug Dispos. 2003, 24, 45–51. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uehara, M.; Tabata, E.; Ishii, K.; Sawa, A.; Ohno, M.; Sakaguchi, M.; Matoska, V.; Bauer, P.O.; Oyama, F. Chitinase mRNA Levels Determined by QPCR in Crab-Eating Monkey (Macaca fascicularis) Tissues: Species-Specific Expression of Acidic Mammalian Chitinase and Chitotriosidase. Genes 2018, 9, 244. https://doi.org/10.3390/genes9050244
Uehara M, Tabata E, Ishii K, Sawa A, Ohno M, Sakaguchi M, Matoska V, Bauer PO, Oyama F. Chitinase mRNA Levels Determined by QPCR in Crab-Eating Monkey (Macaca fascicularis) Tissues: Species-Specific Expression of Acidic Mammalian Chitinase and Chitotriosidase. Genes. 2018; 9(5):244. https://doi.org/10.3390/genes9050244
Chicago/Turabian StyleUehara, Maiko, Eri Tabata, Kazuhiro Ishii, Akira Sawa, Misa Ohno, Masayoshi Sakaguchi, Vaclav Matoska, Peter O. Bauer, and Fumitaka Oyama. 2018. "Chitinase mRNA Levels Determined by QPCR in Crab-Eating Monkey (Macaca fascicularis) Tissues: Species-Specific Expression of Acidic Mammalian Chitinase and Chitotriosidase" Genes 9, no. 5: 244. https://doi.org/10.3390/genes9050244
APA StyleUehara, M., Tabata, E., Ishii, K., Sawa, A., Ohno, M., Sakaguchi, M., Matoska, V., Bauer, P. O., & Oyama, F. (2018). Chitinase mRNA Levels Determined by QPCR in Crab-Eating Monkey (Macaca fascicularis) Tissues: Species-Specific Expression of Acidic Mammalian Chitinase and Chitotriosidase. Genes, 9(5), 244. https://doi.org/10.3390/genes9050244