Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling



Abstract
:1. Introduction
2. Significance of Ca2+ Signaling in Trypanosomatids
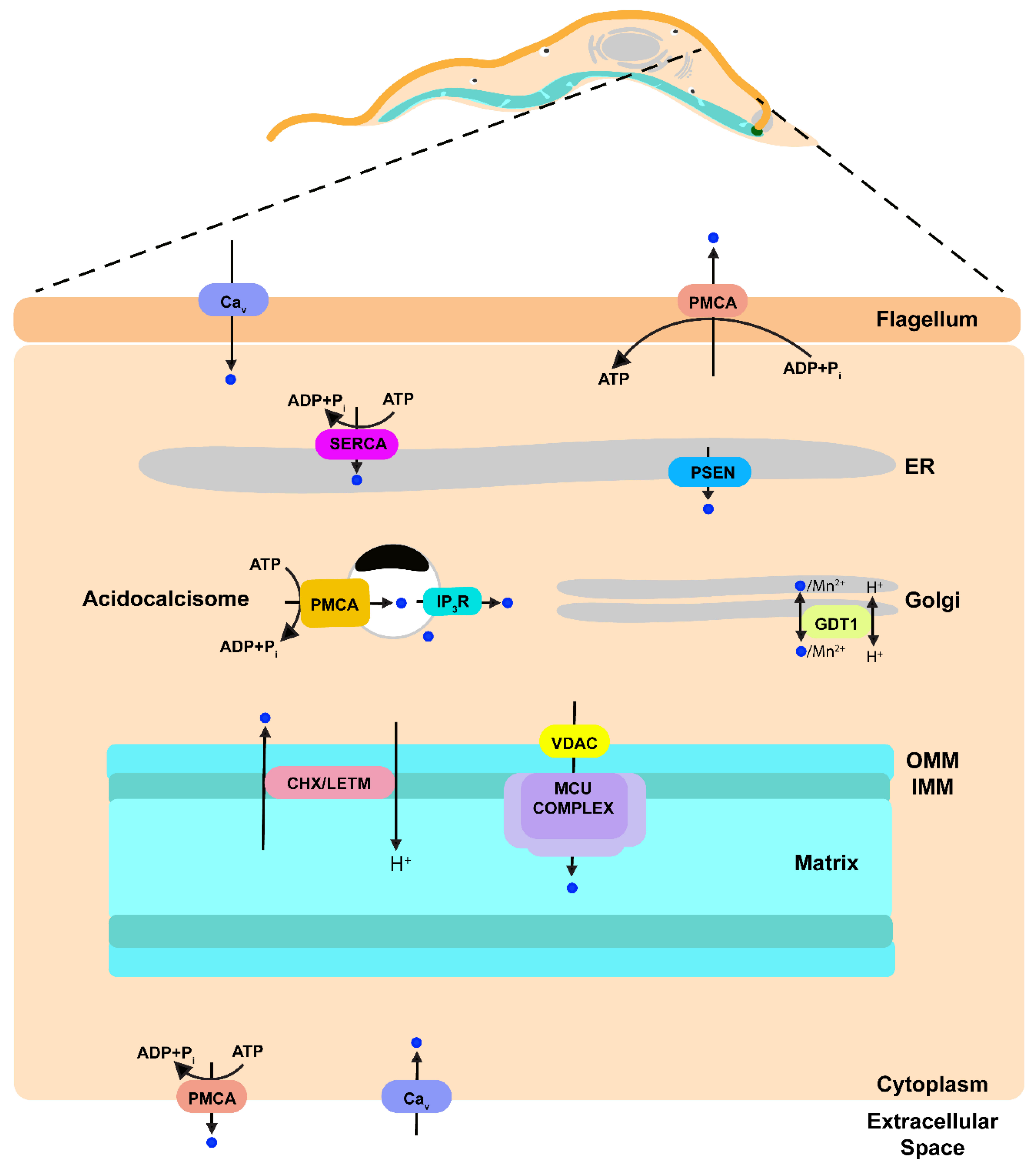
3. Calcium Transport Proteins in the Plasma Membrane
4. Calcium Transport Proteins in the Membrane of the Endoplasmic Reticulum
5. Calcium Transport Proteins in the Membranes of the Mitochondrion
6. Calcium Transport Proteins in the Membrane of Acidocalcisomes
7. An Emerging Role for Membrane Contact Sites in Ca2+ Signaling
8. A Potential Role for the Golgi Complex in Ca2+ Signaling
9. Calcium Transport Proteins in the Flagellar Membrane
10. Ca2+ Binding Proteins
11. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- Docampo, R.; Vercesi, A.E. Characteristics of Ca2+ transport by Trypanosoma cruzi mitochondria in situ. Arch. Biochem. Biophys. 1989, 272, 122–129. [Google Scholar] [CrossRef]
- Docampo, R.; Vercesi, A.E. Ca2+ transport by coupled Trypanosoma cruzi mitochondria in situ. J. Biol. Chem. 1989, 264, 108–111. [Google Scholar] [PubMed]
- Docampo, R.; Lukes, J. Trypanosomes and the solution to a 50-year mitochondrial calcium mystery. Trends Parasitol. 2012, 28, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perocchi, F.; Gohil, V.M.; Girgis, H.S.; Bao, X.R.; McCombs, J.E.; Palmer, A.E.; Mootha, V.K. MICU1 encodes a mitochondrial EF hand protein required for Ca2+ uptake. Nature 2010, 467, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Baughman, J.M.; Perocchi, F.; Girgis, H.S.; Plovanich, M.; Belcher-Timme, C.A.; Sancak, Y.; Bao, X.R.; Strittmatter, L.; Goldberger, O.; Bogorad, R.L.; et al. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 2011, 476, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Stefani, D.; Raffaello, A.; Teardo, E.; Szabo, I.; Rizzuto, R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 2011, 476, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Vercesi, A.E.; Docampo, R. Essential regulation of cell bioenergetics in Trypanosoma brucei by the mitochondrial calcium uniporter. Nat. Commun. 2013, 4, 2865. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Liu, J.; Nguyen, T.; Liu, C.; Sun, J.; Teng, Y.; Fergusson, M.M.; Rovira, I.I.; Allen, M.; Springer, D.A.; et al. The physiological role of mitochondrial calcium revealed by mice lacking the mitochondrial calcium uniporter. Nat. Cell Biol. 2013, 15, 1464–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docampo, R.; de Souza, W.; Miranda, K.; Rohloff, P.; Moreno, S.N. Acidocalcisomes—Conserved from bacteria to man. Nat. Rev. Microbiol. 2005, 3, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Bartlett, P.J.; Thomas, A.P.; Moreno, S.N.; Docampo, R. Acidocalcisomes of Trypanosoma brucei have an inositol 1,4,5-trisphosphate receptor that is required for growth and infectivity. Proc. Natl. Acad. Sci. USA 2013, 110, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Lander, N.; Chiurillo, M.A.; Storey, M.; Vercesi, A.E.; Docampo, R. CRISPR/Cas9-mediated endogenous C-terminal tagging of Trypanosoma cruzi genes reveals the acidocalcisome localization of the inositol 1,4,5-trisphosphate receptor. J. Biol. Chem. 2016, 291, 25505–25515. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Krebs, J. Why calcium? How calcium became the best communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.D. Charge density-dependent strength of hydration and biological structure. Biophys. J. 1997, 72, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Parsons, M.; Ruben, L. Pathways involved in environmental sensing in trypanosomatids. Parasitol. Today 2000, 16, 56–62. [Google Scholar] [CrossRef]
- Swulius, M.T.; Waxham, M.N. Ca2+/calmodulin-dependent protein kinases. Cell. Mol. Life Sci. 2008, 65, 2637–2657. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, N.P.; de Souza, C.F.; Saraiva, F.M.; Sultano, P.E.; Dalmau, S.R.; Bruno, R.E.; Goncalves Rde, L.; Laranja, G.A.; Leal, L.H.; Coelho, M.G.; et al. Heme-induced ros in Trypanosoma cruzi activates CaMKII-like that triggers epimastigote proliferation. One helpful effect of ROS. PLoS ONE 2011, 6, e25935. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.F.; Carneiro, A.B.; Silveira, A.B.; Laranja, G.A.; Silva-Neto, M.A.; Costa, S.C.; Paes, M.C. Heme-induced Trypanosoma cruzi proliferation is mediated by CaM kinase II. Biochem. Biophys. Res. Commun. 2009, 390, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ogueta, S.; Intosh, G.M.; Tellez-Iñon, M.T. Regulation of Ca2+/calmodulin-dependent protein kinase from Trypanosoma cruzi. Mol. Biochem. Parasitol. 1996, 78, 171–183. [Google Scholar] [CrossRef]
- Ogueta, S.B.; Macintosh, G.C.; Tellez-Iñon, M.T. Stage-specific substrate phosphorylation by a Ca2+/calmodulin-dependent protein kinase in Trypanosoma cruzi. J. Eukaryot. Microbiol. 1998, 45, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Kashuba, C.; Docampo, R.; Moreno, S.N. A novel phosphatidylinositol-phospholipase C of Trypanosoma cruzi that is lipid modified and activated during trypomastigote to amastigote differentiation. J. Biol. Chem. 2000, 275, 6428–6438. [Google Scholar] [CrossRef] [PubMed]
- Lammel, E.M.; Barbieri, M.A.; Wilkowsky, S.E.; Bertini, F.; Isola, E.L. Trypanosoma cruzi: Involvement of intracellular calcium in multiplication and differentiation. Exp. Parasitol. 1996, 83, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Stojdl, D.F.; Clarke, M.W. Trypanosoma brucei: Analysis of cytoplasmic Ca2+ during differentiation of bloodstream stages in vitro. Exp. Parasitol. 1996, 83, 134–146. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, M.A.; Montagna, A.E.; Sanguineti, S.; Torres, H.N.; Flawia, M.M. A novel calcium-stimulated adenylyl cyclase from Trypanosoma cruzi, which interacts with the structural flagellar protein paraflagellar rod. J. Biol. Chem. 2002, 277, 35025–35034. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Hu, H.; Lun, Z.R.; Li, Z. Centrin3 in trypanosomes maintains the stability of a flagellar inner-arm dynein for cell motility. Nat. Commun. 2014, 5, 4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvapandiyan, A.; Kumar, P.; Morris, J.C.; Salisbury, J.L.; Wang, C.C.; Nakhasi, H.L. Centrin1 is required for organelle segregation and cytokinesis in Trypanosoma brucei. Mol. Biol. Cell 2007, 18, 3290–3301. [Google Scholar] [CrossRef] [PubMed]
- Araya, J.E.; Cornejo, A.; Orrego, P.R.; Cordero, E.M.; Cortez, M.; Olivares, H.; Neira, I.; Sagua, H.; da Silveira, J.F.; Yoshida, N.; et al. Calcineurin B of the human protozoan parasite Trypanosoma cruzi is involved in cell invasion. Microbes Infect. 2008, 10, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Selvapandiyan, A.; Debrabant, A.; Duncan, R.; Muller, J.; Salotra, P.; Sreenivas, G.; Salisbury, J.L.; Nakhasi, H.L. Centrin gene disruption impairs stage-specific basal body duplication and cell cycle progression in Leishmania. J. Biol. Chem. 2004, 279, 25703–25710. [Google Scholar] [CrossRef] [PubMed]
- Selvapandiyan, A.; Duncan, R.; Debrabant, A.; Bertholet, S.; Sreenivas, G.; Negi, N.S.; Salotra, P.; Nakhasi, H.L. Expression of a mutant form of Leishmania donovani centrin reduces the growth of the parasite. J. Biol. Chem. 2001, 276, 43253–43261. [Google Scholar] [CrossRef] [PubMed]
- Moreno, V.R.; Aguero, F.; Tekiel, V.; Sanchez, D.O. The calcineurin a homologue from Trypanosoma cruzi lacks two important regulatory domains. Acta Trop. 2007, 101, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.N.; Silva, J.; Vercesi, A.E.; Docampo, R. Cytosolic-free calcium elevation in Trypanosoma cruzi is required for cell invasion. J. Exp. Med. 1994, 180, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.G.; Zhong, L.; Chang, K.P.; Docampo, R. Intracellular Ca2+ pool content and signaling and expression of a calcium pump are linked to virulence in Leishmania mexicana amazonesis amastigotes. J. Biol. Chem. 1997, 272, 9464–9473. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, M.A.; Majumder, S.; Kierszenbaum, F. Changes in Trypanosoma cruzi infectivity by treatments that affect calcium ion levels. Mol. Biochem. Parasitol. 1994, 66, 119–125. [Google Scholar] [CrossRef]
- Rohloff, P.; Rodrigues, C.O.; Docampo, R. Regulatory volume decrease in Trypanosoma cruzi involves amino acid efflux and changes in intracellular calcium. Mol. Biochem. Parasitol. 2003, 126, 219–230. [Google Scholar] [CrossRef]
- Irigoin, F.; Inada, N.M.; Fernandes, M.P.; Piacenza, L.; Gadelha, F.R.; Vercesi, A.E.; Radi, R. Mitochondrial calcium overload triggers complement-dependent superoxide-mediated programmed cell death in Trypanosoma cruzi. Biochem. J. 2009, 418, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Selzer, P.M.; Webster, P.; Duszenko, M. Influence of Ca2+ depletion on cytoskeleton and nucleolus morphology in Trypanosoma brucei. Eur. J. Cell. Biol. 1991, 56, 104–112. [Google Scholar] [PubMed]
- Prole, D.L.; Taylor, C.W. Identification of intracellular and plasma membrane calcium channel homologues in pathogenic parasites. PLoS ONE 2011, 6, e26218. [Google Scholar] [CrossRef] [PubMed]
- Oberholzer, M.; Langousis, G.; Nguyen, H.T.; Saada, E.A.; Shimogawa, M.M.; Jonsson, Z.O.; Nguyen, S.M.; Wohlschlegel, J.A.; Hill, K.L. Independent analysis of the flagellum surface and matrix proteomes provides insight into flagellum signaling in mammalian-infectious Trypanosoma brucei. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Benaim, G.; Garcia-Marchan, Y.; Reyes, C.; Uzcanga, G.; Figarella, K. Identification of a sphingosine-sensitive Ca2+ channel in the plasma membrane of Leishmania mexicana. Biochem. Biophys. Res. Commun. 2013, 430, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Martinez, A.K.; Rodriguez-Duran, J.; Serrano-Martin, X.; Hernandez-Rodriguez, V.; Benaim, G. Mechanism of action of miltefosine on Leishmania donovani involves the impairment of acidocalcisome function and the activation of the sphingosine-dependent plasma membrane Ca2+ channel. Antimicrob. Agents Chemother. 2018, 62, e01614–e1617. [Google Scholar] [CrossRef] [PubMed]
- Docampo, R.; Huang, G. Calcium signaling in trypanosomatid parasites. Cell Calcium 2015, 57, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.C.; McLatchie, A.P.; Kelly, J.M. Evidence that transport of iron from the lysosome to the cytosol in african trypanosomes is mediated by a mucolipin orthologue. Mol. Microbiol. 2013, 89, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Bustos, T.; Moreno, S.N.J.; Docampo, R. Detection of weakly expressed Trypanosoma cruzi membrane proteins using high-performance probes. J. Eukaryot. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Prakriya, M.; Lewis, R.S. Store-operated calcium channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, P.; Cintron, R.; Docampo, R. Calcium homeostasis and acidocalcisomes in Trypanosoma cruzi. In Structures and Organelles in Pathogenic Protists; Souza, W.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 299–318. [Google Scholar]
- Lu, H.G.; Zhong, L.; de Souza, W.; Benchimol, M.; Moreno, S.; Docampo, R. Ca2+ content and expression of an acidocalcisomal calcium pump are elevated in intracellular forms of Trypanosoma cruzi. Mol. Cell. Biol. 1998, 18, 2309–2323. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Rohloff, P.; Cox, J.; Uyemura, S.A.; Docampo, R. Trypanosoma brucei plasma membrane-type Ca2+-ATPase 1 (TbPMC1) and 2 (TbPMC2) genes encode functional Ca2+-ATPases localized to the acidocalcisomes and plasma membrane, and essential for Ca2+ homeostasis and growth. J. Biol. Chem. 2004, 279, 14427–14439. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Mukherjee, T.; Sarkar, S.; Majumdar, S.; Bhaduri, A. The plasma-membrane Ca2+-ATPase of Leishmania donovani is an extrusion pump for Ca2+. Biochem. J. 1997, 322 Pt 1, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Benaim, G.; Losada, S.; Gadelha, F.R.; Docampo, R. A calmodulin-activated (Ca2+-Mg2+)-ATPase is involved in Ca2+ transport by plasma membrane vesicles from Trypanosoma cruzi. Biochem. J. 1991, 280 Pt 3, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Benaim, G.; Cervino, V.; Hermoso, T.; Felibert, P.; Laurentin, A. Intracellular calcium homeostasis in Leishmania mexicana. Identification and characterization of a plasma membrane calmodulin-dependent Ca2+-ATPase. Biol Res 1993, 26, 141–150. [Google Scholar] [PubMed]
- Ramirez-Iglesias, J.R.; Perez-Gordones, M.C.; Del Castillo, J.R.; Mijares, A.; Benaim, G.; Mendoza, M. Identification and characterization of a calmodulin binding domain in the plasma membrane Ca2+-ATPase from Trypanosoma equiperdum. Mol. Biochem. Parasitol. 2018, 222, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gordones, M.C.; Ramirez-Iglesias, J.R.; Cervino, V.; Uzcanga, G.L.; Benaim, G.; Mendoza, M. Evidence of the presence of a calmodulin-sensitive plasma membrane Ca2+-ATPase in Trypanosoma equiperdum. Mol. Biochem. Parasitol. 2017, 213, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.H.; Hashimi, H.; Lun, Z.R.; Ayala, F.J.; Lukes, J. Adaptations of Trypanosoma brucei to gradual loss of kinetoplast DNA: Trypanosoma equiperdum and Trypanosoma evansi are petite mutants of T. brucei. Proc. Natl. Acad. Sci. USA 2008, 105, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Nolan, D.P.; Reverlard, P.; Pays, E. Overexpression and characterization of a gene for a Ca2+-ATPase of the endoplasmic reticulum in Trypanosoma brucei. J. Biol. Chem. 1994, 269, 26045–26051. [Google Scholar] [PubMed]
- Mendoza, M.; Mijares, A.; Rojas, H.; Colina, C.; Cervino, V.; DiPolo, R.; Benaim, G. Evaluation of the presence of a thapsigargin-sensitive calcium store in trypanosomatids using Trypanosoma evansi as a model. J. Parasitol. 2004, 90, 1181–1183. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Okura, M.; Ruiz, F.A.; Scott, D.A.; Docampo, R. TcSCA complements yeast mutants defective in Ca2+ pumps and encodes a Ca2+-ATPase that localizes to the endoplasmic reticulum of Trypanosoma cruzi. J. Biol. Chem. 2001, 276, 32437–32445. [Google Scholar] [CrossRef] [PubMed]
- Conte, I.; Labriola, C.; Cazzulo, J.J.; Docampo, R.; Parodi, A.J. The interplay between folding-facilitating mechanisms in Trypanosoma cruzi endoplasmic reticulum. Mol. Biol. Cell 2003, 14, 3529–3540. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, G.; Valck, C.; Aguilar, L.; Kemmerling, U.; Lopez-Munoz, R.; Cabrera, G.; Morello, A.; Ferreira, J.; Maya, J.D.; Galanti, N.; et al. Roles of Trypanosoma cruzi calreticulin in parasite-host interactions and in tumor growth. Mol. Immunol. 2012, 52, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, M.T.; Tenner, A.J.; Bobak, D.A.; Joiner, K.A. Complement component C1q enhances invasion of human mononuclear phagocytes and fibroblasts by Trypanosoma cruzi trypomastigotes. J. Clin. Investig. 1989, 84, 1982–1989. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Toloza, G.; Abello, P.; Ferreira, A. Is the antitumor property of Trypanosoma cruzi infection mediated by its calreticulin? Front. Immunol. 2016, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Pogue, G.P.; Duncan, R.C.; Lee, N.S.; Singh, N.K.; Atreya, C.D.; Dwyer, D.M.; Nakhasi, H.L. Isolation and characterization of Leishmania donovani calreticulin gene and its conservation of the RNA binding activity. Mol. Biochem. Parasitol. 1996, 81, 53–64. [Google Scholar] [CrossRef]
- Debrabant, A.; Lee, N.; Dwyer, D.M.; Nakhasi, H.L. Role of calreticulin in Leishmania parasite secretory pathway and pathogenesis. In Calreticulin; Eggleton, P.M.M., Ed.; Springer: Boston, MA, USA, 2003; pp. 220–237. [Google Scholar]
- Honarnejad, K.; Herms, J. Presenilins: Role in calcium homeostasis. Int. J. Biochem. Cell Biol. 2012, 44, 1983–1986. [Google Scholar] [CrossRef] [PubMed]
- Denton, R.M. Regulation of mitochondrial dehydrogenases by calcium ions. Biochim. Biophys. Acta 2009, 1787, 1309–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denton, R.M.; Randle, P.J.; Martin, B.R. Stimulation by calcium ions of pyruvate dehydrogenase phosphate phosphatase. Biochem. J. 1972, 128, 161–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, J.G.; Denton, R.M. The effects of calcium ions and adenine nucleotides on the activity of pig heart 2-oxoglutarate dehydrogenase complex. Biochem. J. 1979, 180, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gincel, D.; Zaid, H.; Shoshan-Barmatz, V. Calcium binding and translocation by the voltage-dependent anion channel: A possible regulatory mechanism in mitochondrial function. Biochem. J. 2001, 358, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Raffaello, A.; De Stefani, D.; Sabbadin, D.; Teardo, E.; Merli, G.; Picard, A.; Checchetto, V.; Moro, S.; Szabo, I.; Rizzuto, R. The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit. EMBO J. 2013, 32, 2362–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plovanich, M.; Bogorad, R.L.; Sancak, Y.; Kamer, K.J.; Strittmatter, L.; Li, A.A.; Girgis, H.S.; Kuchimanchi, S.; De Groot, J.; Speciner, L.; et al. MICU2, a paralog of MICU1, resides within the mitochondrial uniporter complex to regulate calcium handling. PLoS ONE 2013, 8, e55785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, Y.; Markhard, A.L.; Kitami, T.; Kovacs-Bogdan, E.; Kamer, K.J.; Udeshi, N.D.; Carr, S.A.; Chaudhuri, D.; Clapham, D.E.; Li, A.A.; et al. EMRE is an essential component of the mitochondrial calcium uniporter complex. Science 2013, 342, 1379–1382. [Google Scholar] [CrossRef] [PubMed]
- Mallilankaraman, K.; Cardenas, C.; Doonan, P.J.; Chandramoorthy, H.C.; Irrinki, K.M.; Golenar, T.; Csordas, G.; Madireddi, P.; Yang, J.; Muller, M.; et al. MCUR1 is an essential component of mitochondrial Ca2+ uptake that regulates cellular metabolism. Nat. Cell Biol. 2012, 14, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Paupe, V.; Prudent, J.; Dassa, E.P.; Rendon, O.Z.; Shoubridge, E.A. Ccdc90a (MCUR1) is a cytochrome c oxidase assembly factor and not a regulator of the mitochondrial calcium uniporter. Cell Metab. 2015, 21, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Vais, H.; Tanis, J.E.; Muller, M.; Payne, R.; Mallilankaraman, K.; Foskett, J.K. MCUR1, CCDC90A, is a regulator of the mitochondrial calcium uniporter. Cell Metab. 2015, 22, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Pusnik, M.; Charriere, F.; Maser, P.; Waller, R.F.; Dagley, M.J.; Lithgow, T.; Schneider, A. The single mitochondrial porin of Trypanosoma brucei is the main metabolite transporter in the outer mitochondrial membrane. Mol. Biol. Evol. 2009, 26, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Flinner, N.; Schleiff, E.; Mirus, O. Identification of two voltage-dependent anion channel-like protein sequences conserved in kinetoplastida. Biol. Lett. 2012, 8, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Chiurillo, M.A.; Lander, N.; Bertolini, M.S.; Storey, M.; Vercesi, A.E.; Docampo, R. Different roles of mitochondrial calcium uniporter complex subunits in growth and infectivity of Trypanosoma cruzi. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Docampo, R.; Vercesi, A.E.; Huang, G. Mitochondrial calcium transport in trypanosomes. Mol. Biochem. Parasitol. 2014, 196, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palty, R.; Silverman, W.F.; Hershfinkel, M.; Caporale, T.; Sensi, S.L.; Parnis, J.; Nolte, C.; Fishman, D.; Shoshan-Barmatz, V.; Herrmann, S.; et al. NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc. Natl. Acad. Sci. USA 2010, 107, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhao, L.; Clapham, D.E. Genome-wide RNAi screen identifies LETM1 as a mitochondrial Ca2+/H+ antiporter. Science 2009, 326, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Hashimi, H.; McDonald, L.; Stribrna, E.; Lukes, J. Trypanosome Letm1 protein is essential for mitochondrial potassium homeostasis. J. Biol. Chem. 2013, 288, 26914–26925. [Google Scholar] [CrossRef] [PubMed]
- Froschauer, E.; Nowikovsky, K.; Schweyen, R.J. Electroneutral K+/H+ exchange in mitochondrial membrane vesicles involves YOL027/Letm1 proteins. Biochim. Biophys. Acta 2005, 1711, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.; Tavakoli, M.; Pfeiffer, C.; Seifert, J.; Mattarei, A.; De Stefani, D.; Zoratti, M.; Nowikovsky, K. Letm1-mediated K+ and Na+ homeostasis regulates mitochondrial Ca2+ efflux. Front. Physiol. 2017, 8, 839. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, J.A.; Engel, J.C.; Leapman, R.D.; Swyt, C.R.; Pella, P.A. Trypanosoma cruzi: Elemental composition heterogeneity of cloned stocks. Mol. Biochem. Parasitol. 1988, 31, 19–26. [Google Scholar] [CrossRef]
- Scott, D.A.; Docampo, R. Two types of H+-ATPase are involved in the acidification of internal compartments in Trypanosoma cruzi. Biochem. J. 1998, 331 Pt 2, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Docampo, R.; Scott, D.A.; Vercesi, A.E.; Moreno, S.N. Intracellular Ca2+ storage in acidocalcisomes of Trypanosoma cruzi. Biochem. J. 1995, 310 Pt 3, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Docampo, R. Proteomic analysis of acidocalcisomes of Trypanosoma brucei uncovers their role in phosphate metabolism, cation homeostasis, and calcium signaling. Commun. Integr. Biol. 2015, 8, e1017174. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Enomoto, M.; Morales, J.; Kurebayashi, N.; Sakurai, T.; Hashimoto, T.; Nara, T.; Mikoshiba, K. Inositol 1,4,5-trisphosphate receptor regulates replication, differentiation, infectivity and virulence of the parasitic protist Trypanosoma cruzi. Mol. Microbiol. 2013, 87, 1133–1150. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Morales, J.; Uemura, H.; Mikoshiba, K.; Nara, T. A novel method for inducing amastigote-to-trypomastigote transformation in vitro in Trypanosoma cruzi reveals the importance of inositol 1,4,5-trisphosphate receptor. PLoS ONE 2015, 10, e0135726. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.L. The endoplasmic reticulum and calcium storage. Bioessays 1990, 12, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.J.; Voeltz, G.K. Structure and function of ER membrane contact sites with other organelles. Nat. Rev. Mol. Cell. Biol. 2016, 17, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; John Peter, A.T.; Kornmann, B. ER-mitochondria contact sites in yeast: Beyond the myths of ERMES. Curr. Opin. Cell. Biol. 2015, 35, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Hawes, C.; Hussey, P.J. Plant endoplasmic reticulum-plasma membrane contact sites. Trends Plant Sci. 2017, 22, 289–297. [Google Scholar] [CrossRef] [PubMed]
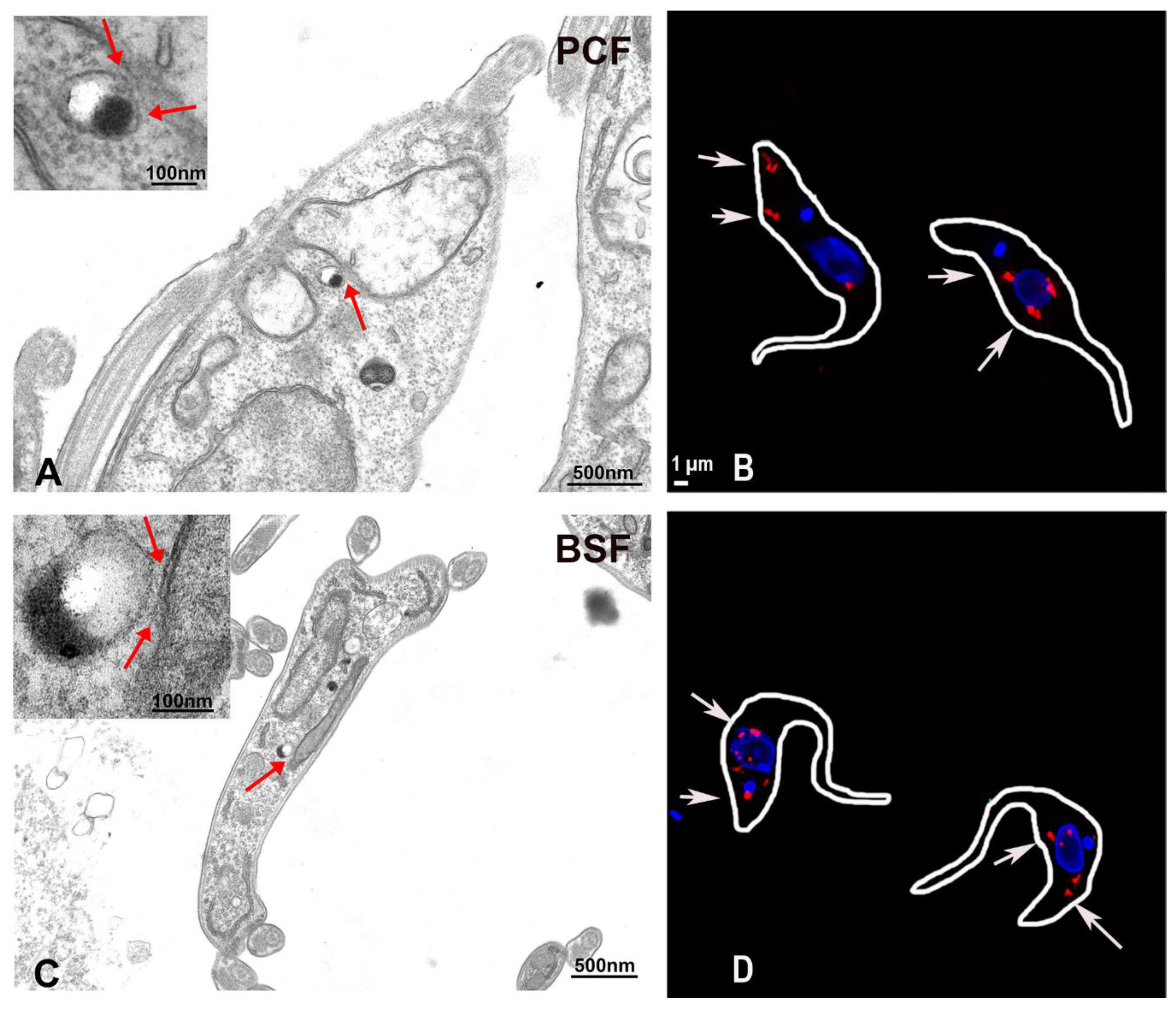
- Ramakrishnan, S.; Asady, B.; Docampo, R. Acidocalcisome-mitochondrion membrane contact sites in Trypanosoma brucei. Pathogens 2018, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.H.; Ridgley, E.L.; Enis, D.; Olness, F.; Ruben, L. Selective transfer of calcium from an acidic compartment to the mitochondrion of Trypanosoma brucei. Measurements with targeted aequorins. J. Biol. Chem. 1997, 272, 31022–31028. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Mohamalawari, D.; Rao, R. A novel isoform of the secretory pathway Ca2+,Mn2+-ATPase, HsPCA2, has unusual properties and is expressed in the brain. J. Biol. Chem. 2005, 280, 11608–11614. [Google Scholar] [CrossRef] [PubMed]
- Vanoevelen, J.; Dode, L.; Van Baelen, K.; Fairclough, R.J.; Missiaen, L.; Raeymaekers, L.; Wuytack, F. The secretory pathway Ca2+/Mn2+-ATPase 2 is a Golgi-localized pump with high affinity for Ca2+ ions. J. Biol. Chem. 2005, 280, 22800–22808. [Google Scholar] [CrossRef] [PubMed]
- Sorin, A.; Rosas, G.; Rao, R. PMR1, a Ca2+-ATPase in yeast Golgi, has properties distinct from sarco/endoplasmic reticulum and plasma membrane calcium pumps. J. Biol. Chem. 1997, 272, 9895–9901. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, P.; Lissandron, V.; Capitanio, P.; Pozzan, T. Ca2+ signalling in the Golgi apparatus. Cell Calcium 2011, 50, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Colinet, A.S.; Thines, L.; Deschamps, A.; Flemal, G.; Demaegd, D.; Morsomme, P. Acidic and uncharged polar residues in the consensus motifs of the yeast Ca2+ transporter GDT1p are required for calcium transport. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Colinet, A.S.; Sengottaiyan, P.; Deschamps, A.; Colsoul, M.L.; Thines, L.; Demaegd, D.; Duchene, M.C.; Foulquier, F.; Hols, P.; Morsomme, P. Yeast GDT1 is a Golgi-localized calcium transporter required for stress-induced calcium signaling and protein glycosylation. Sci. Rep. 2016, 6, 24282. [Google Scholar] [CrossRef] [PubMed]
- Demaegd, D.; Foulquier, F.; Colinet, A.S.; Gremillon, L.; Legrand, D.; Mariot, P.; Peiter, E.; Van Schaftingen, E.; Matthijs, G.; Morsomme, P. Newly characterized Golgi-localized family of proteins is involved in calcium and pH homeostasis in yeast and human cells. Proc. Natl. Acad. Sci. USA 2013, 110, 6859–6864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulary, E.; Potelle, S.; Legrand, D.; Foulquier, F. TMEM165 deficiencies in congenital disorders of glycosylation type II (CDG-II): Clues and evidences for roles of the protein in Golgi functions and ion homeostasis. Tissue Cell 2017, 49, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Micaroni, M. Calcium around the Golgi apparatus: Implications for intracellular membrane trafficking. Adv. Exp. Med. Biol. 2012, 740, 439–460. [Google Scholar] [PubMed]
- Cifuentes, F.; Gonzalez, C.E.; Fiordelisio, T.; Guerrero, G.; Lai, F.A.; Hernandez-Cruz, A. A ryanodine fluorescent derivative reveals the presence of high-affinity ryanodine binding sites in the golgi complex of rat sympathetic neurons, with possible functional roles in intracellular Ca2+ signaling. Cell. Signal. 2001, 13, 353–362. [Google Scholar] [CrossRef]
- Vickerman, K. On the surface coat and flagellar adhesion in trypanosomes. J. Cell Sci. 1969, 5, 163–193. [Google Scholar] [PubMed]
- Sugrue, P.; Hirons, M.R.; Adam, J.U.; Holwill, M.E. Flagellar wave reversal in the kinetoplastid flagellate Crithidia oncopelti. Biol. Cell 1988, 63, 127–131. [Google Scholar] [PubMed]
- Selvapandiyan, A.; Kumar, P.; Salisbury, J.L.; Wang, C.C.; Nakhasi, H.L. Role of centrins 2 and 3 in organelle segregation and cytokinesis in Trypanosoma brucei. PLoS ONE 2012, 7, e45288. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Deford, J.; Benjamin, R.; Lee, M.G.; Ruben, L. The gene family of EF-hand calcium-binding proteins from the flagellum of Trypanosoma brucei. Biochem. J. 1994, 304 Pt 3, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Emmer, B.T.; Daniels, M.D.; Taylor, J.M.; Epting, C.L.; Engman, D.M. Calflagin inhibition prolongs host survival and suppresses parasitemia in Trypanosoma brucei infection. Eukaryot. Cell 2010, 9, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Engman, D.M.; Krause, K.H.; Blumin, J.H.; Kim, K.S.; Kirchhoff, L.V.; Donelson, J.E. A novel flagellar Ca2+-binding protein in trypanosomes. J. Biol. Chem. 1989, 264, 18627–18631. [Google Scholar] [PubMed]
- Wingard, J.N.; Ladner, J.; Vanarotti, M.; Fisher, A.J.; Robinson, H.; Buchanan, K.T.; Engman, D.M.; Ames, J.B. Structural insights into membrane targeting by the flagellar calcium-binding protein (FCaBP), a myristoylated and palmitoylated calcium sensor in Trypanosoma cruzi. J. Biol. Chem. 2008, 283, 23388–23396. [Google Scholar] [CrossRef] [PubMed]
- Ridgley, E.; Webster, P.; Patton, C.; Ruben, L. Calmodulin-binding properties of the paraflagellar rod complex from Trypanosoma brucei. Mol. Biochem. Parasitol. 2000, 109, 195–201. [Google Scholar] [CrossRef]
- Ginger, M.L.; Collingridge, P.W.; Brown, R.W.; Sproat, R.; Shaw, M.K.; Gull, K. Calmodulin is required for paraflagellar rod assembly and flagellum-cell body attachment in trypanosomes. Protist 2013, 164, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Portman, N.; Lacomble, S.; Thomas, B.; McKean, P.G.; Gull, K. Combining RNA interference mutants and comparative proteomics to identify protein components and dependences in a eukaryotic flagellum. J. Biol. Chem. 2009, 284, 5610–5619. [Google Scholar] [CrossRef] [PubMed]
- Tellez-Inon, M.T.; Ulloa, R.M.; Torruella, M.; Torres, H.N. Calmodulin and Ca2+-dependent cyclic AMP phosphodiesterase activity in Trypanosoma cruzi. Mol. Biochem. Parasitol. 1985, 17, 143–153. [Google Scholar] [CrossRef]
- Wayne A Snedden, H.F. Calmodulin as a versatile calcium signal transducer in plants. New Phytol. 2001, 151, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Villalobo, A.; Ishida, H.; Vogel, H.J.; Berchtold, M.W. Calmodulin as a protein linker and a regulator of adaptor/scaffold proteins. Biochim. Biophys. Acta 2018, 1865, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Rohloff, P.; Montalvetti, A.; Docampo, R. Acidocalcisomes and the contractile vacuole complex are involved in osmoregulation in Trypanosoma cruzi. J. Biol. Chem. 2004, 279, 52270–52281. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, P.N.; Jimenez, V.; Park, M.; Martins, V.P.; Atwood, J., 3rd; Moles, K.; Collins, D.; Rohloff, P.; Tarleton, R.; Moreno, S.N.; et al. Identification of contractile vacuole proteins in Trypanosoma cruzi. PLoS ONE 2011, 6, e18013. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Docampo, R. Acidic calcium stores open for business: Expanding the potential for intracellular Ca2+ signaling. Trends Cell Biol. 2010, 20, 277–286. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Organelle | Protein | Trypanosoma brucei | Trypanosoma cruzi | Leishmania major |
|---|---|---|---|---|
| Endoplasmic Reticulum (ER) | SERCA | Tb927.5.3400 | TcCLB.509777.70 | LmjF.04.0010 |
| Tb927.9.15460 | TcCLB.506241.70 | LmjF.35.2080 | ||
| PSEN | Tb927.9.4940 | TcCLB.508277.50 | LmjF.15.1530 | |
| Mitochondrion | VDAC | Tb927.2.2510 | TcCLB.504225.20 | LmjF.02.0460 |
| Tb927.2.2520 | TcCLB.508741.229 | |||
| TcCLB.509141.40 | ||||
| TcCLB.511687.10 | ||||
| TcCLB.511687.10 | ||||
| VDAC like 2 | Tb927.4.1610 | TcCLB504057 | LmjF.34.3100 | |
| MCU | Tb927.11.1350 | TcCLB.503893.120 | LmjF.27.0780 | |
| MICU1 | Tb927.8.1850 | TcCLB.511391.210 | LmjF.07.0110 | |
| MICU2 | Tb927.7.2960 | TcCLB.510525.130 | ||
| MCUb | Tb927.10.300 | TcCLB.504069 | LmjF.21.1690 | |
| LETM1 | Tb927.3.4920 | TcCLB.507951.270 | LmjF.29.0920 | |
| Acidocalcisome | Ca2+ ATPase | Tb927.8.1180 | TcCLB.508543.90 | LmjF.07.0630 |
| LmjF.07.0650 | ||||
| IP3R | Tb927.8.2770 | TcCLB.509461.90 | LmjF.16.0280 | |
| Golgi | Gdt1 | - | TcCLB.508895.70 | LmjF.19.0310 |
| Plasma membrane | PMCA | Tb927.8.1180 | TcCLB.506401.70 | LmjF.07.0630 |
| Tb927.8.1160 | TcCLB.508543.90 | LmjF.07.0650 | ||
| Tb927.8.1200 | TcCLB.510769.120 | LmjF.17.0600 | ||
| Tb927.10.11620 | TcCLB.509647.150 | LmjF.33.1010 | ||
| Cav channel (flagellum) | Tb927.10.2880 | TcCLB.504105.130 | LmjF.34.0480 | |
| LmjF.17.1440 | ||||
| Acidic stores | TRP | Tb927.8.850 | TcCLB.510861.94 | LmjF.07.0910. |
| TRPML | Tb927.7.950 | TcCLB.503463.20 | LmjF.26.0990 | |
| TcCLB.503735.30 |
| Protein | T. brucei | T. cruzi | L. major |
|---|---|---|---|
| Calreticulin | Tb927.8.7410 | TcCLB.509011.40 | LmjF.31.2600 |
| Flagellar Ca2+-binding protein | Tb927.8.5440 | TcCLB.509391.10 | LmjF.16.0910 |
| Tb927.8.5460 | TcCLB.509391.20 | LmjF.16.0920 | |
| Tb927.8.5465 | TcCLB.509391.30 | ||
| Tb927.8.5470 | TcCLB.506749.20 | ||
| Ca2+-binding protein | Tb927.6.2720 | TcCLB.507925.60 | LmjF.30.1240 |
| Tb927.4.1740 | TcCLB.510879.190 | LmjF.34.2950 | |
| Calmodulin (CaM) | Tb927.11.13020 | TcCLB.507483.30 | LmjF.09.0910 |
| Tb927.11.13030 | TcCLB.507483.39 | LmjF.09.0920 | |
| Tb927.11.13040 | LmjF.09.0930 | ||
| Tb927.11.13050 | |||
| CaM-like protein | Tb927.11.9790 | TcCLB.506963.90 | LmjF.36.3675 |
| Tb927.9.11230 | TcCLB.504075.3 | LmjF.35.3890 | |
| Tb927.11.3680 | TcCLB.508731.30 | LmjF.13.1160 | |
| Tb927.11.7940 | TcCLB.506933.89 | LmjF.28.0800 | |
| Tb927.9.6130 | TcCLB.508951.50 | LmjF.21.0220 | |
| Tb927.6.4710 | TcCLB.511729.9 | LmjF.15.0930 | |
| TcCLB.507483.50 | LmjF.30.3360 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramakrishnan, S.; Docampo, R. Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling. Genes 2018, 9, 304. https://doi.org/10.3390/genes9060304
Ramakrishnan S, Docampo R. Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling. Genes. 2018; 9(6):304. https://doi.org/10.3390/genes9060304
Chicago/Turabian StyleRamakrishnan, Srinivasan, and Roberto Docampo. 2018. "Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling" Genes 9, no. 6: 304. https://doi.org/10.3390/genes9060304
APA StyleRamakrishnan, S., & Docampo, R. (2018). Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling. Genes, 9(6), 304. https://doi.org/10.3390/genes9060304