Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae


{kind=link}
Abstract
:1. Introduction
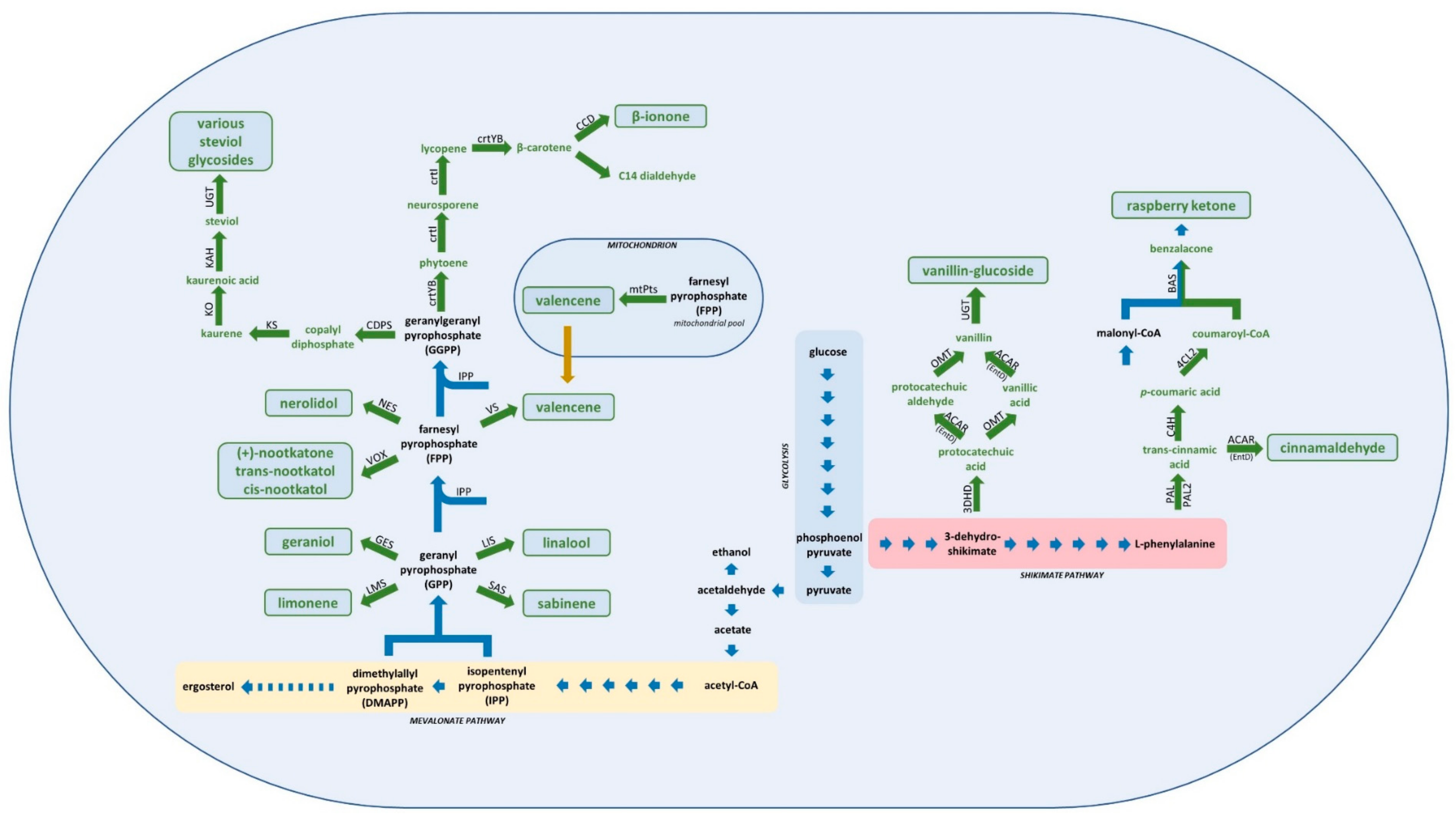
2. Heterologous Compounds Derived from Intermediates of the Shikimate Pathway
3. Heterologous Compounds Derived from Intermediates of the Mevalonate Pathway
3.1. Isoprenoids
3.1.1. Monoterpenes
3.1.2. Sesquiterpenes
3.1.3. Norisoprenoids
4. Next Generation Sweeteners
Steviol Glycosides
5. Conclusions
Funding
Conflicts of Interest
References
- Galanie, S.; Thodey, K.; Trenchard, I.J.; Filsinger Interrante, M.; Smolke, C.D. Complete biosynthesis of opioids in yeast. Science 2015, 349, 1095–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, T.L.K.H.; Kong, I.I.; Liu, J.J.; Zhang, G.C.; Jin, Y.S. Engineering and Evolution of Saccharomyces Cerevisiae to Produce Biofuels and Chemicals; Springer International Publishing: Cham, Switzerland, 2018; pp. 175–215. [Google Scholar]
- Teo, W.S.; Ling, H.; Yu, A.-Q.; Chang, M.W. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid short- and branched-chain alkyl esters biodiesel. Biotechnol. Biofuels 2015, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Amaro, R.; Nicaud, J.-M. Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol. 2016, 34, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.R.; Shin, J.H.; Cho, J.S.; Yang, D.; Lee, S.Y. Systems metabolic engineering of Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Priefert, H.; Rabenhorst, J.; Steinbuchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Brochado, A.R.; Matos, C.; Moller, B.L.; Hansen, J.; Mortensen, U.H.; Patil, K.R. Improved vanillin production in baker’s yeast through in silico design. Microb. Cell Factories 2010, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, N.J.; Narbad, A.; Faulds, C.; Williamson, G. Novel approaches to the biosynthesis of vanillin. Curr. Opin. Biotechnol. 2000, 11, 490–496. [Google Scholar] [CrossRef]
- Hansen, E.H.; Moller, B.L.; Kock, G.R.; Bunner, C.M.; Kristensen, C.; Jensen, O.R.; Okkels, F.T.; Olsen, C.E.; Motawia, M.S.; Hansen, J. De novo biosynthesis of vanillin in fission yeast (Schizosaccharomyces pombe) and baker’s yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Brochado, A.R.; Patil, K.R. Overexpression of O-methyltransferase leads to improved vanillin production in baker’s yeast only when complemented with model-guided network engineering. Biotechnol. Bioeng. 2013, 110, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Strucko, T.; Magdenoska, O.; Mortensen, U.H. Benchmarking two commonly used Saccharomyces cerevisiae strains for heterologous vanillin-β-glucoside production. Metab. Eng. Commun. 2015, 2, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Poll, L. Odour thresholds of some important aroma compounds in raspberries. Z. Lebensm.-Unters. Forsch. 1990, 191, 129–131. [Google Scholar] [CrossRef]
- Larsen, M.; Poll, L.; Callesen, O.; Lewis, M. Relations between the content of aroma compounds and the sensory evaluation of 10 raspberry varieties (Rubus idaeus L.). Acta Agric. Scand. 1991, 41, 447–454. [Google Scholar] [CrossRef]
- Lee, D.; Lloyd, N.D.; Pretorius, I.S.; Borneman, A.R. Heterologous production of raspberry ketone in the wine yeast Saccharomyces cerevisiae via pathway engineering and synthetic enzyme fusion. Microb. Cell Factories 2016, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Stabnikova, O.; Wang, J.-Y.; Ivanov, V. Value-added biotechnological products from organic wastes. In Environment Biotechnology; Humana Press: Totowa, NJ, USA, 2010; pp. 343–394. [Google Scholar]
- Abe, I.; Takahashi, Y.; Morita, H.; Noguchi, H. Benzalacetone synthase. A novel polyketide synthase that plays a crucial role in the biosynthesis of phenylbutanones in Rheum palmatum. Eur. J. Biochem. 2001, 268, 3354–3359. [Google Scholar] [CrossRef] [PubMed]
- Borejsza-Wysocki, W.; Hrazdina, G. Biosynthesis of p-hydroxyphenylbutan-2-one in raspberry fruits and tissue cultures. Phytochemistry 1994, 35, 623–628. [Google Scholar] [CrossRef]
- Koeduka, T.; Watanabe, B.; Suzuki, S.; Hiratake, J.; Mano, J.; Yazaki, K. Characterization of raspberry ketone/zingerone synthase, catalyzing the alpha, beta-hydrogenation of phenylbutenones in raspberry fruits. Biochem. Biophys. Res. Commun. 2011, 412, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van der Meer, I.M.; Sibbesen, O.; Broekgaarden, M.; Qvist, I.; Mikkelsen, J.D.; Hall, R.D. Microbial production of natural raspberry ketone. Biotechnol. J. 2007, 2, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Vannelli, T.; Qi, W.W.; Sweigard, J.; Gatenby, A.A.; Sariaslani, F.S. Production of p-hydroxycinnamic acid from glucose in Saccharomyces cerevisiae and Escherichia coli by expression of heterologous genes from plants and fungi. Metab. Eng. 2007, 9, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Jung, S.M.; Kim, M.D.; Han, N.S.; Seo, J.H. Production of resveratrol from tyrosine in metabolically engineered Saccharomyces cerevisiae. Enzyme. Microb. Technol. 2012, 51, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Raina, V.K.; Srivastava, S.K.; Aggarwal, K.K.; Ramesh, S.; Kumar, S. Essential oil composition of Cinnamomum zeylanicum Blume leaves from Little Andaman, India. Flavour Fragr. J. 2001, 16, 374–376. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Rao, L.J.; Sakariah, K.K. Chemical composition of the volatile oil from the fruits of Cinnamomum zeylanicum blume. Flavour Fragr. J. 1997, 12, 331–333. [Google Scholar] [CrossRef]
- Friedman, M.; Kozukue, N.; Harden, L.A. Cinnamaldehyde content in foods determined by gas chromatography-mass spectrometry. J. Agric. Food. Chem. 2000, 48, 5702–5709. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union: Commission Implementing Regulation (EU) No 872/2012, 1st October 2012. Available online: https://publications.europa.eu/en/publication-detail/-/publication/9d00bfce-0bc9-11e2-8e28-01aa75ed71a1/language-en (accessed on 11 May 2018).
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Gottardi, M.; Knudsen, J.D.; Prado, L.; Oreb, M.; Branduardi, P.; Boles, E. De novo biosynthesis of trans-cinnamic acid derivatives in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 4883–4893. [Google Scholar] [CrossRef] [PubMed]
- Larroy, C.; Fernandez, M.R.; Gonzalez, E.; Pares, X.; Biosca, J.A. Characterization of the Saccharomyces cerevisiae YMR318C (ADH6) gene product as a broad specificity NADPH-dependent alcohol dehydrogenase: Relevance in aldehyde reduction. Biochem. J. 2002, 361, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Croteau, R.B. Resin-based defenses in conifers. Trends Plant Sci. 1999, 4, 184–190. [Google Scholar] [CrossRef]
- Loza-Tavera, H. Monoterpenes in essential oils. Biosynthesis and properties. Adv. Exp. Med. Biol. 1999, 464, 49–62. [Google Scholar] [PubMed]
- Winter, R. A Consumer’s Dictionary of Food Additives, 7th ed.; Random House USA Inc.: New York, NY, USA, 2009; p. 592. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Oswald, M.; Fischer, M.; Dirninger, N.; Karst, F. Monoterpenoid biosynthesis in Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Withers, S.T.; Keasling, J.D. Biosynthesis and engineering of isoprenoid small molecules. Appl. Microbiol. Biotechnol. 2007, 73, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Chambon, C.; Ladeveze, V.; Oulmouden, A.; Servouse, M.; Karst, F. Isolation and properties of yeast mutants affected in farnesyl diphosphate synthetase. Curr. Genet. 1990, 18, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Gang, D.R.; Fridman, E.; Lewinsohn, E.; Pichersky, E. Characterization of geraniol synthase from the peltate glands of sweet basil. Plant Physiol. 2004, 134, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.J.; Meyer, S.; Claudel, P.; Bergdoll, M.; Karst, F. Metabolic engineering of monoterpene synthesis in yeast. Biotechnol. Bioeng. 2011, 108, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, J.; Wang, H.X.; Zeng, Y. A geraniol-synthase gene from cinnamomum tenuipilum. Phytochemistry 2005, 66, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Miettinen, K.; Goedbloed, M.; Verstappen, F.W.; Voster, A.; Jongsma, M.A.; Memelink, J.; van der Krol, S.; Bouwmeester, H.J. Characterization of two geraniol synthases from Valeriana officinalis and Lippia dulcis: Similar activity but difference in subcellular localization. Metab. Eng. 2013, 20, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Polakowski, T.; Stahl, U.; Lang, C. Overexpression of a cytosolic hydroxymethylglutaryl-coa reductase leads to squalene accumulation in yeast. Appl. Microbiol. Biotechnol. 1998, 49, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Du, G.; Chen, J.; Zhou, J. Overproduction of geraniol by enhanced precursor supply in Saccharomyces cerevisiae. J. Biotechnol. 2013, 168, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bao, X.; Li, C.; Shen, Y.; Hou, J. Improving monoterpene geraniol production through geranyl diphosphate synthesis regulation in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 100, 4561–4571. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.Z.; Yao, M.D.; Wang, Y.; Zhou, L.; Song, T.Q.; Liu, H.; Xiao, W.H.; Yuan, Y.J. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metab. Eng. 2017, 41, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Pardo, E.; Rico, J.; Gil, J.V.; Orejas, M. De novo production of six key grape aroma monoterpenes by a geraniol synthase-engineered S. cerevisiae wine strain. Microb. Cell Factories 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Cseke, L.; Blanc, V.M.; Pichersky, E. Evolution of floral scent in clarkia: Novel patterns of s-linalool synthase gene expression in the C. breweri flower. Plant Cell 1996, 8, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Herrero, O.; Ramon, D.; Orejas, M. Engineering the Saccharomyces cerevisiae isoprenoid pathway for de novo production of aromatic monoterpenes in wine. Metab. Eng. 2008, 10, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Rico, J.; Pardo, E.; Orejas, M. Enhanced production of a plant monoterpene by overexpression of the 3-hydroxy-3-methylglutaryl coenzyme a reductase catalytic domain in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2010, 76, 6449–6454. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Shahpiri, A.; Asadollahi, M.A.; Momenbeik, F.; Partow, S. Metabolic engineering of Saccharomyces cerevisiae for linalool production. Biotechnol. Lett. 2016, 38, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duetz, W.A.; Bouwmeester, H.; van Beilen, J.B.; Witholt, B. Biotransformation of limonene by bacteria, fungi, yeasts, and plants. Appl. Microbiol. Biotechnol. 2003, 61, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, B.M. Biosynthesis and biotechnology of high-value p-menthane monoterpenes, including menthol, carvone, and limonene. Adv. Biochem. Eng. Biotechnol. 2015, 148, 319–353. [Google Scholar] [CrossRef] [PubMed]
- Tranchida, P.Q.; Zoccali, M.; Bonaccorsi, I.; Dugo, P.; Mondello, L.; Dugo, G. The off-line combination of high performance liquid chromatography and comprehensive two-dimensional gas chromatography-mass spectrometry: A powerful approach for highly detailed essential oil analysis. J. Chromatogr. A 2013, 1305, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; van Griensven, L.J. Antibacterial effects of the essential oils of commonly consumed medicinal herbs using an in vitro model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [PubMed]
- Behrendorff, J.B.; Vickers, C.E.; Chrysanthopoulos, P.; Nielsen, L.K. 2,2-diphenyl-1-picrylhydrazyl as a screening tool for recombinant monoterpene biosynthesis. Microb. Cell Factories 2013, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Jongedijk, E.; Cankar, K.; Ranzijn, J.; van der Krol, S.; Bouwmeester, H.; Beekwilder, J. Capturing of the monoterpene olefin limonene produced in Saccharomyces cerevisiae. Yeast 2015, 32, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Jongedijk, E.; Cankar, K.; Buchhaupt, M.; Schrader, J.; Bouwmeester, H.; Beekwilder, J. Biotechnological production of limonene in microorganisms. Appl. Microbiol. Biotechnol. 2016, 100, 2927–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietschmann, M.; Vostrowsky, O.; Bestmann, H.J.; Pant, A.K.; Mathela, C.S. Volatile constituents of Hyptis pectinata Poit. (lamiaceae). J. Essent. Oil Res. 1998, 10, 550–552. [Google Scholar] [CrossRef]
- Arunkumar, R.; Nair, S.A.; Rameshkumar, K.B.; Subramoniam, A. The essential oil constituents of Zornia diphylla (L.) Pers, and anti-inflammatory and antimicrobial activities of the oil. Rec. Nat. Prod. 2014, 8, 385–393. [Google Scholar]
- Kampranis, S.C.; Ioannidis, D.; Purvis, A.; Mahrez, W.; Ninga, E.; Katerelos, N.A.; Anssour, S.; Dunwell, J.M.; Degenhardt, J.; Makris, A.M.; et al. Rational conversion of substrate and product specificity in a salvia monoterpene synthase: Structural insights into the evolution of terpene synthase function. Plant Cell 2007, 19, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Sharon-Asa, L.; Shalit, M.; Frydman, A.; Bar, E.; Holland, D.; Or, E.; Lavi, U.; Lewinsohn, E.; Eyal, Y. Citrus fruit flavor and aroma biosynthesis: Isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J. 2003, 36, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, M.; Hashimoto, T.; Noma, Y.; Asakawa, Y. Highly efficient production of nootkatone, the grapefruit aroma from valencene, by biotransformation. Chem. Pharm. Bull. 2005, 53, 1513–1514. [Google Scholar] [CrossRef] [PubMed]
- Farhi, M.; Marhevka, E.; Masci, T.; Marcos, E.; Eyal, Y.; Ovadis, M.; Abeliovich, H.; Vainstein, A. Harnessing yeast subcellular compartments for the production of plant terpenoids. Metab. Eng. 2011, 13, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van Houwelingen, A.; Cankar, K.; van Dijk, A.D.; de Jong, R.M.; Stoopen, G.; Bouwmeester, H.; Achkar, J.; Sonke, T.; Bosch, D. Valencene synthase from the heartwood of nootka cypress (Callitropsis nootkatensis) for biotechnological production of valencene. Plant Biotechnol. J. 2014, 12, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; MacNevin, G.; Ro, D.K. De novo synthesis of high-value plant sesquiterpenoids in yeast. Methods Enzymol. 2012, 517, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Hurt, E.C.; Pesold-Hurt, B.; Suda, K.; Oppliger, W.; Schatz, G. The first twelve amino acids (less than half of the pre-sequence) of an imported mitochondrial protein can direct mouse cytosolic dihydrofolate reductase into the yeast mitochondrial matrix. EMBO J. 1985, 4, 2061–2068. [Google Scholar] [PubMed]
- Haring, H.G.; Rijkens, F.; Boelens, H.; Van der Gen, A. Olfactory studies on enantiomeric eremophilane sesquiterpenoids. J. Agric. Food Chem. 1972, 20, 1018–1021. [Google Scholar] [CrossRef]
- Leonhardt, R.-H.; Berger, R.G. Nootkatone. Adv. Biochem. Eng. Biotechnol. 2015, 148, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, J.A.; Ortuno, A.; Garcia-Puig, D.; Porras, I.; Garcia-Lidon, A.; Sabater, F. Variations of nootkatone and valencene levels during the development of grapefruit. J. Agric. Food Chem. 1992, 40, 1488–1490. [Google Scholar] [CrossRef]
- Fraatz, M.A.; Berger, R.G.; Zorn, H. Nootkatone—A biotechnological challenge. Appl. Microbiol. Biotechnol. 2009, 83, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Hunter, G.L.K.; Brogden, W.B., Jr. Conversion of valencene to nootkatone. J. Food Sci. 1965, 30, 876–878. [Google Scholar] [CrossRef]
- Wilson, C.W.; Shaw, P.E. Synthesis of nootkatone from valencene. J. Agric. Food Chem. 1978, 26, 1430–1432. [Google Scholar] [CrossRef]
- Cankar, K.; van Houwelingen, A.; Bosch, D.; Sonke, T.; Bouwmeester, H.; Beekwilder, J. A chicory cytochrome P450 mono-oxygenase CYP71AV8 for the oxidation of (+)-valencene. FEBS Lett. 2011, 585, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Gavira, C.; Hofer, R.; Lesot, A.; Lambert, F.; Zucca, J.; Werck-Reichhart, D. Challenges and pitfalls of P450-dependent (+)-valencene bioconversion by Saccharomyces cerevisiae. Metab. Eng. 2013, 18, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Urban, P.; Mignotte, C.; Kazmaier, M.; Delorme, F.; Pompon, D. Cloning, yeast expression, and characterization of the coupling of two distantly related Arabidopsis thaliana NADPH-cytochrome P450 reductases with P450 CYP73A5. J. Biol. Chem. 1997, 272, 19176–19186. [Google Scholar] [CrossRef] [PubMed]
- Emmerstorfer, A.; Wimmer-Teubenbacher, M.; Wriessnegger, T.; Leitner, E.; Muller, M.; Kaluzna, I.; Schurmann, M.; Mink, D.; Zellnig, G.; Schwab, H.; et al. Over-expression of ice2 stabilizes cytochrome P450 reductase in Saccharomyces cerevisiae and Pichia pastoris. Biotechnol. J. 2015, 10, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Plan, M.R.; Chrysanthopoulos, P.; Hodson, M.P.; Nielsen, L.K.; Vickers, C.E. A squalene synthase protein degradation method for improved sesquiterpene production in Saccharomyces cerevisiae. Metab. Eng. 2017, 39, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Mateus, C.; Avery, S.V. Destabilized green fluorescent protein for monitoring dynamic changes in yeast gene expression with flow cytometry. Yeast 2000, 16, 1313–1323. [Google Scholar] [CrossRef] [Green Version]
- Salama, S.R.; Hendricks, K.B.; Thorner, J. G1 cyclin degradation: The PEST motif of yeast Cln2 is necessary, but not sufficient, for rapid protein turnover. Mol. Cell. Biol. 1994, 14, 7953–7966. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bustamante, E.; Sanchez, S. Microbial production of C13-norisoprenoids and other aroma compounds via carotenoid cleavage. Crit. Rev. Microbiol. 2007, 33, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Enzell, C.R. Biodegradation of carotenoids—An important route to aroma compounds. Pure Appl. Chem. 1985, 57, 693–700. [Google Scholar] [CrossRef]
- Burdock, G.A. Fenaroli’s Handbook of Flavor Ingredients, 6th ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van Rossum, H.M.; Koopman, F.; Sonntag, F.; Buchhaupt, M.; Schrader, J.; Hall, R.D.; Bosch, D.; Pronk, J.T.; van Maris, A.J.A.; et al. Polycistronic expression of a β-carotene biosynthetic pathway in Saccharomyces cerevisiae coupled to β-ionone production. J. Biotechnol. 2014, 192, 383–392. [Google Scholar] [CrossRef] [PubMed]
- López, J.; Essus, K.; Kim, I.-K.; Pereira, R.; Herzog, J.; Siewers, V.; Nielsen, J.; Agosin, E. Production of β-ionone by combined expression of carotenogenic and plant Ccd1 genes in Saccharomyces cerevisiae. Microb. Cell Factories 2015, 14, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalcinati, G.; Partow, S.; Siewers, V.; Schalk, M.; Daviet, L.; Nielsen, J. Combined metabolic engineering of precursor and co-factor supply to increase α-santalene production by Saccharomyces cerevisiae. Microb. Cell Factories 2012, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, I.A. A review of recent evidence relating to sugars, insulin resistance and diabetes. Eur. J. Nutr. 2016, 55, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, W.H. Early uses of Stevia rebaudiana (Asteraceae) leaves as a sweetener in paraguay. AGRIS 1992, 46, 336–337. [Google Scholar]
- Prakash, I.; Markosyan, A.; Bunders, C. Development of next generation stevia sweetener: Rebaudioside M. Foods 2014, 3, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Prakash, I.; Dubois, G.E.; Clos, J.F.; Wilkens, K.L.; Fosdick, L.E. Development of rebiana, a natural, non-caloric sweetener. Food. Chem. Toxicol. 2008, 46 (Suppl. 7), S75–S82. [Google Scholar] [CrossRef] [PubMed]
- Wanke, M.; Skorupinska-Tudek, K.; Swiezewska, E. Isoprenoid biosynthesis via 1-deoxy-d-xylulose 5-phosphate/2-C-methyl-d-erythritol 4-phosphate (DOXP/MEP) pathway. Acta Biochim. Pol. 2001, 48, 663–672. [Google Scholar] [PubMed]
- Oka, T.; Jigami, Y. Reconstruction of de novo pathway for synthesis of UDP-glucuronic acid and UDP-xylose from intrinsic UDP-glucose in Saccharomyces cerevisiae. FEBS J. 2006, 273, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Wang, Y.; Chen, L.; Yan, M.; Chen, K.; Xu, L.; Ouyang, P. Production of rebaudioside a from stevioside catalyzed by the engineered Saccharomyces cerevisiae. Appl. Biochem. Biotechnol. 2016, 178, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Olsson, K.; Carlsen, S.; Semmler, A.; Simon, E.; Mikkelsen, M.D.; Moller, B.L. Microbial production of next-generation stevia sweeteners. Microb. Cell Factories 2016, 15, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, M.D.; Hansen, J.; Simon, E.; Brianza, F.; Semmler, A.; Olsson, K.; Carlsen, S.; Düring, L.; Ouspensku, A.; Hicks, P. Methods for improved production of rebaudioside D and rebaudioside M. U.S. Patent No. 9,957,540, 1 May 2018. [Google Scholar]
- Global Markets for Flavors and Fragrances. Available online: www.bccresearch.com/market-research/chemicals/flavors-fragrances-global-markets-chm034c.html (accessed on 23 May 2018).
- Evolva, A.O. Available online: https://www.evolva.com/ (accessed on 21 May 2018).
- Lian, J.; Mishra, S.; Zhao, H. Recent advances in metabolic engineering of Saccharomyces cerevisiae: New tools and their applications. Metab. Eng. 2018. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutyna, D.R.; Borneman, A.R. Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae. Genes 2018, 9, 326. https://doi.org/10.3390/genes9070326
Kutyna DR, Borneman AR. Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae. Genes. 2018; 9(7):326. https://doi.org/10.3390/genes9070326
Chicago/Turabian StyleKutyna, Dariusz R., and Anthony R. Borneman. 2018. "Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae" Genes 9, no. 7: 326. https://doi.org/10.3390/genes9070326
APA StyleKutyna, D. R., & Borneman, A. R. (2018). Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae. Genes, 9(7), 326. https://doi.org/10.3390/genes9070326