

Dynamical Properties of Discrete-Time HTLV-I and HIV-1 within-Host Coinfection Model

Abstract
:1. Introduction
2. Discrete-Time HTLV-I and HIV-1 Co-Infection Model
3. Preliminaries
4. Equilibria
5. Global Stability
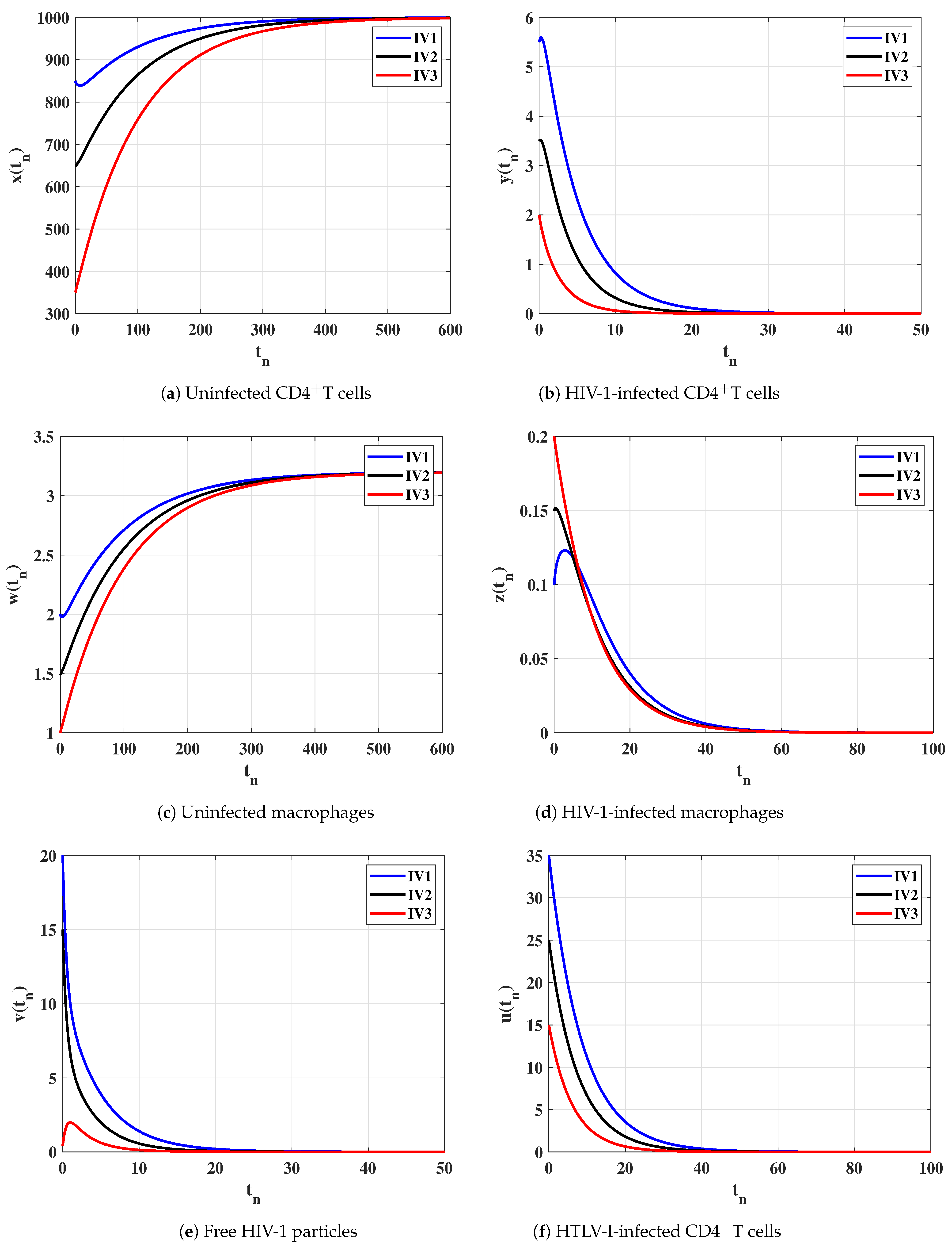
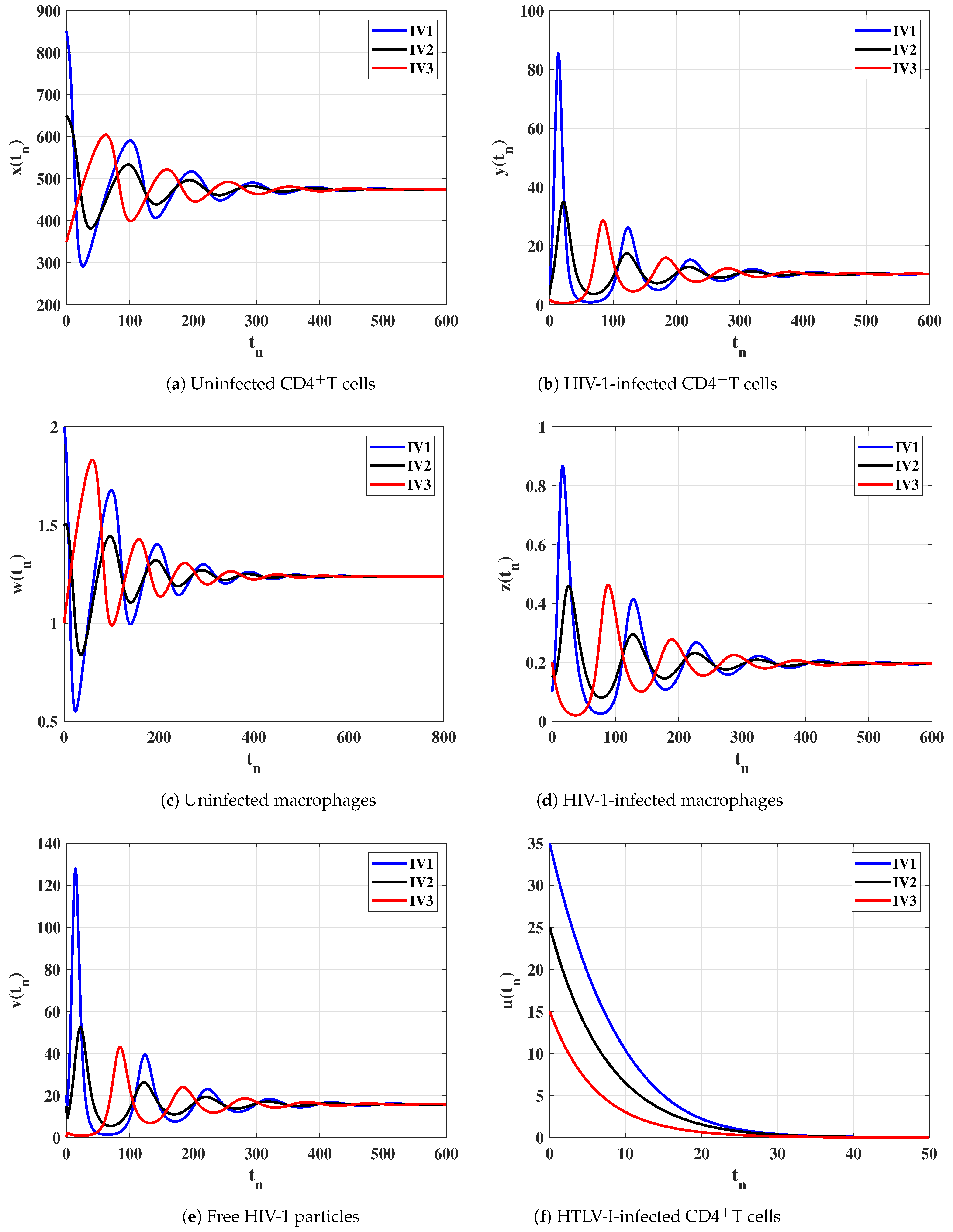
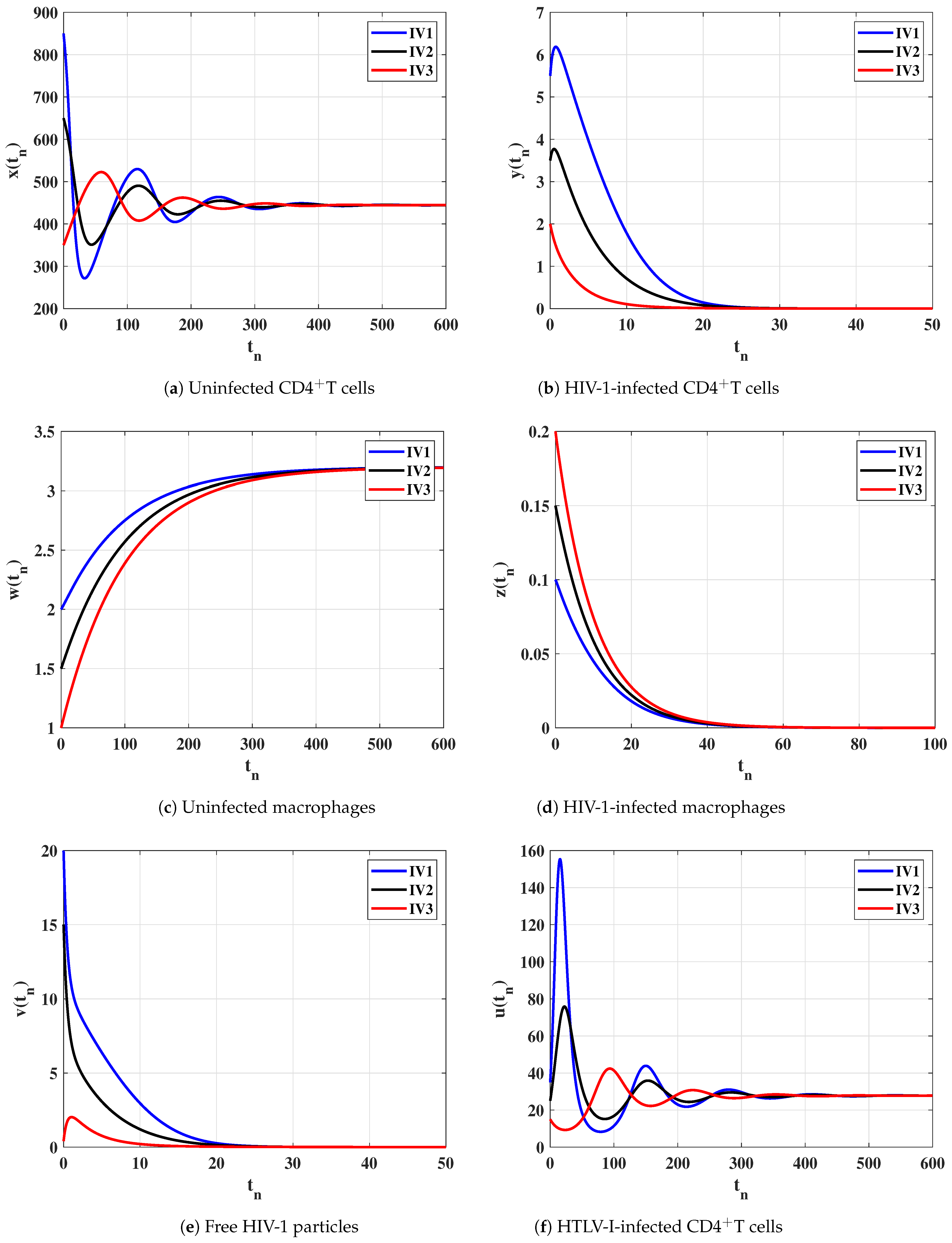
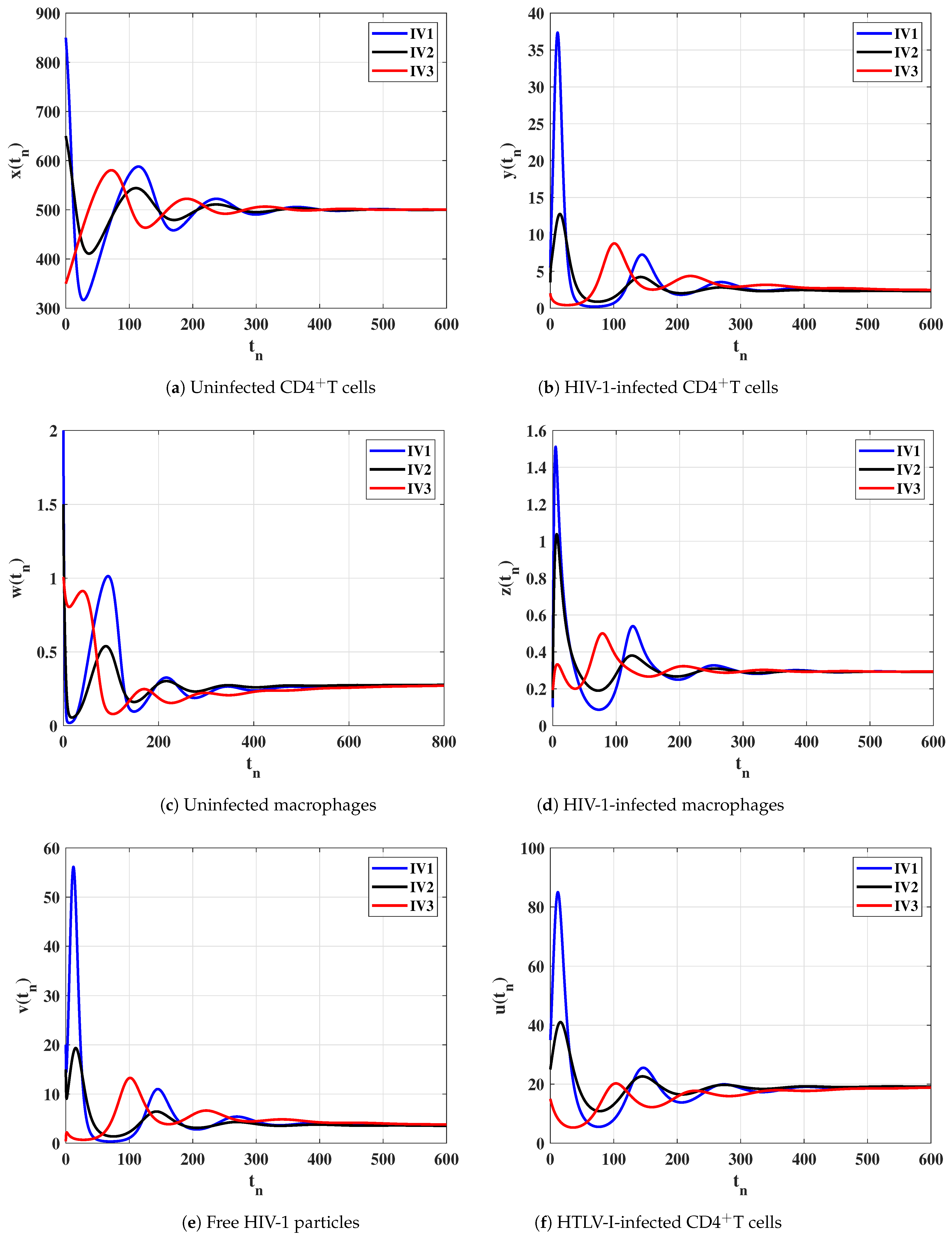
6. Numerical Simulations
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciupe, S.M.; Heffernan, J.M. In-host modeling. Infect. Dis. Model. 2017, 2, 188–202. [Google Scholar] [CrossRef]
- Nowak, M.A.; Bangham, C.R.M. Population dynamics of immune responses to persistent viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, X.; Tang, S.; Rong, L. Dynamics of an HIV model with multiple infection stages and treatment with different drug classes. Bull. Math. Biol. 2016, 78, 322–349. [Google Scholar] [CrossRef] [PubMed]
- Perelson, A.S.; Essunger, P.; Cao, Y.; Vesanen, M.; Hurley, A.; Saksela, K.; Markowitz, M.; Ho, D.D. Decay characteristics of HIV-1-infected compartments during combination therapy. Nature 1997, 387, 188–191. [Google Scholar] [CrossRef]
- Nelson, P.W.; Perelson, A.S. Mathematical analysis of delay differential equation models of HIV-1 infection. Math. Biosci. 2002, 179, 73–94. [Google Scholar] [CrossRef]
- Nelson, P.W.; Murray, J.D.; Perelson, A.S. A model of HIV-1 pathogenesis that includes an intracellular delay. Math. Biosci. 2000, 163, 201–215. [Google Scholar] [CrossRef]
- Perelson, A.S.; Nelson, P.W. Mathematical analysis of HIV-1 dynamics in vivo. SIAM Rev. 1999, 41, 3–44. [Google Scholar] [CrossRef]
- Lin, J.; Xu, R.; Tian, X. Threshold dynamics of an HIV-1 model with both viral and cellular infections, cell-mediated and humoral immune responses. Math. Biosci. Eng. 2018, 16, 292–319. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J. Threshold dynamics of a delayed nonlocal reaction-diffusion HIV infection model with both cell-free and cell-to-cell transmissions. J. Math. Anal. Appl. 2020, 488, 124047. [Google Scholar] [CrossRef]
- Feng, T.; Qiu, Z.; Meng, X.; Rong, L. Analysis of a stochastic HIV-1 infection model with degenerate diffusion. Appl. Math. Comput. 2019, 348, 437–455. [Google Scholar] [CrossRef]
- Callaway, D.S.; Perelson, A.S. HIV-1 infection and low steady state viral loads. Bull. Math. Biol. 2002, 64, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.M.; Banks, H.T.; Kwon, H.D.; Tran, H.T. Dynamic multidrug therapies for HIV: Optimal and STI control approaches. Math. Biosci. Eng. 2004, 1, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.M.; Banks, H.T.; Davidian, M.; Kwon, H.D.; Tran, H.T.; Wynne, S.N.; Rosenberg, E.S. HIV dynamics: Modeling, data analysis, and optimal treatment protocols. J. Comput. Appl. Math. 2005, 184, 10–49. [Google Scholar] [CrossRef]
- Qi, K.; Jiang, D.; Hayat, T.; Alsaedi, A. Virus dynamic behavior of a stochastic HIV/AIDS infection model including two kinds of target cell infections and CTL immune. Math. Comput. Simul. 2021, 188, 548–570. [Google Scholar] [CrossRef]
- Lim, A.G.; Maini, P.K. HTLV-I infection: A dynamic struggle between viral persistence and host immunity. J. Theor. Biol. 2014, 352, 92–108. [Google Scholar] [CrossRef]
- Li, M.Y.; Shu, H. Multiple stable periodic oscillations in a mathematical model of CTL response to HTLV-I infection. Bull. Math. Biol. 2011, 73, 1774–1793. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, W. Global dynamics of a reaction and diffusion model for an HTLV-I infection with mitotic division of actively infected cells. J. Appl. Anal. Comput. 2017, 7, 899–930. [Google Scholar] [CrossRef]
- Li, F.; Ma, W. Dynamics analysis of an HTLV-1 infection model with mitotic division of actively infected cells and delayed CTL immune response. Math. Methods Appl. Sci. 2018, 41, 3000–3017. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Heffernan, J.M. Viral dynamics of an HTLV-I infection model with intracellular delay and CTL immune response delay. J. Math. Anal. Appl. 2018, 459, 506–527. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Li, Y.; Xu, D. Complete dynamical analysis for a nonlinear HTLV-I infection model with distributed delay, CTL response and immune impairment. Discret. Contin. Dyn. Ser. B 2020, 25, 917–933. [Google Scholar] [CrossRef] [Green Version]
- Chenar, F.F.; Kyrychko, Y.N.; Blyuss, K.B. Mathematical model of immune response to hepatitis B. J. Theor. Biol. 2018, 447, 98–110. [Google Scholar] [CrossRef]
- Kitagawa, K.; Kuniya, T.; Nakaoka, S.; Asai, Y.; Watashi, K.; Iwami, S. Mathematical Analysis of a Transformed ODE from a PDE Multiscale Model of Hepatitis C Virus Infection. Bull. Math. 2019, 81, 1427–1441. [Google Scholar] [CrossRef]
- Baccam, P.; Beauchemin, C.; Macken, C.A.; Hayden, F.G.; Perelson, A.S. Kinetics of influenza A virus infection in humans. J. Virol. 2006, 80, 7590–7599. [Google Scholar] [CrossRef]
- Nuraini, N.; Tasman, H.; Soewono, E.; Sidarto, K.A. A with-in host dengue infection model with immune response. Math. Comput. Model. 2009, 49, 1148–1155. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X. Stability and Hopf bifurcation of a within-host chikungunya virus infection model with two delays. Math. Comput. Simul. 2017, 138, 31–48. [Google Scholar] [CrossRef]
- Nguyen, V.K.; Binder, S.C.; Boianelli, A.; Meyer-Hermann, M.; Hernandez-Vargas, E.A. Ebola virus infection modelling and identifiability problems. Front. Microbiol. 2015, 6, 257. [Google Scholar] [CrossRef]
- Hernandez-Vargas, E.A.; Velasco-Hernandez, J.X. In-host mathematical modelling of COVID-19 in humans. Annu. Rev. Control 2020, 50, 448–456. [Google Scholar] [CrossRef]
- Chatterjee, A.N.; Basir, F.A.; Almuqrin, M.A.; Mondal, J.; Khan, I. SARS-CoV-2 infection with lytic and nonlytic immune responses: A fractional order optimal control theoretical study. Results Phys. 2021, 26, 104260. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alsulami, R.S.; Hobiny, A.D. Modeling and stability analysis of within-host IAV/SARS-CoV-2 coinfection with antibody immunity. Mathematics 2022, 10, 4382. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Elnahary, E.K. Analysis of general humoral immunity HIV dynamics model with HAART and distributed delays. Mathematics 2019, 7, 157. [Google Scholar] [CrossRef] [Green Version]
- Elaiw, A.M.; AlShamrani, N.H. Analysis of a within-host HTLV-I/HIV-1 co-infection model with immunity. Virus Res. 2021, 295, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M.; AlShamrani, N.H. HTLV/HIV dual Infection: Modeling and analysis. Mathematics 2021, 9, 51. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H.; Dahy, E.; Abdellatif, A. Stability of within host HTLV-I/HIV-1 co-infection in the presence of macrophages. Int. J. Biomath. 2022, 16, 2250066. [Google Scholar] [CrossRef]
- Pasha, S.A.; Nawaz, Y.; Arif, M.S. On the nonstandard finite difference method for reaction–diffusion models. Chaos Solitons Fractals 2023, 166, 112929. [Google Scholar] [CrossRef]
- Maamar, M.H.; Ehrhardt, M.; Tabharit, L. A Nonstandard Finite Difference Scheme for a Time-Fractional Model of Zika Virus Transmission. 2022. Available online: https://www.imacm.uni-wuppertal.de/fileadmin/imacm/preprints/2022/imacm_22_21.pdf (accessed on 20 December 2022).
- Farooqi, A.; Ahmad, R.; Alotaibi, H.; Nofal, T.A.; Farooqi, R.; Khan, I. A comparative epidemiological stability analysis of predictor corrector type non-standard finite difference scheme for the transmissibility of measles. Results Phys. 2021, 21, 103756. [Google Scholar] [CrossRef]
- Mickens, R.E. Nonstandard Finite Difference Models of Differential Equations; World Scientific: Singapore, 1994. [Google Scholar]
- Mickens, R.E. Applications of Nonstandard Finite Difference Schemes; World Scientific: Singapore, 2000. [Google Scholar]
- Treibert, S.; Brunner, H.; Ehrhardt, M. A nonstandard finite difference scheme for the SVICDR model to predict COVID-19 dynamics. Math. Biosci. Eng. 2022, 19, 1213–1238. [Google Scholar]
- Korpusik, A. A nonstandard finite difference scheme for a basic model of cellular immune response to viral infection. Commun. Nonlinear Sci. Numer. Simul. 2017, 43, 369–384. [Google Scholar] [CrossRef]
- Yang, Y.; Xinsheng, M.; Yahui, L. Global stability of a discrete virus dynamics model with Holling type-II infection function. Math. Methods Appl. Sci. 2016, 39, 2078–2082. [Google Scholar] [CrossRef]
- Geng, Y.; Xu, J.; Hou, J. Discretization and dynamic consistency of a delayed and diffusive viral infection model. Appl. Math. Comput. 2018, 316, 282–295. [Google Scholar] [CrossRef]
- Vaz, S.; Torres, D.F.M. Discrete-time system of an intracellular delayed HIV model with CTL immune response. arXiv 2022, arXiv:2205.02199. [Google Scholar]
- Salman, S.M. A nonstandard finite difference scheme and optimal control for an HIV model with Beddington-DeAngelis incidence and cure rate. Eur. Phys. J. Plus 2020, 135, 808. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhu, C.C. A non-standard finite difference scheme for a diffusive HIV-1 infection model with immune response and intracellular delay. Axioms 2022, 11, 129. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshaikh, M.A. Stability preserving NSFD scheme for a general virus dynamics model with antibody and cell-mediated responses. Chaos Solitons Fractals 2020, 138, 109862. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshaikh, M.A. Stability of discrete-time HIV dynamics models with three categories of infected CD4+ T-cells. Adv. Differ. Equ. 2019, 2019, 407. [Google Scholar] [CrossRef]
- Mickens, R.E. Calculation of denominator functions for nonstandard finite difference schemes for differential equations satisfying apositivity condition. Numer. Methods Partial. Differ. Equ. 2007, 23, 672–691. [Google Scholar] [CrossRef]
- Shi, P.; Dong, L. Dynamical behaviors of a discrete HIV-1 virus model with bilinear infective rate. Math. Methods Appl. Sci. 2014, 37, 2271–2280. [Google Scholar] [CrossRef]
- Perelson, A.S.; Kirschner, D.E.; de Boer, R. Dynamics of HIV Infection of CD4+ T cells. Math. Biosci. 1993, 114, 81–125. [Google Scholar] [CrossRef]
- Mohri, H.; Bonhoeffer, S.; Monard, S.; Perelson, A.S.; Ho, D. Rapid turnover of T lymphocytes in SIV-infected rhesus macaques. Science 1998, 279, 1223–1227. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.A.; Azoz, S.A. Stability of delayed HIV dynamics models with two latent reservoirs and immune impairment. Adv. Differ. Equ. 2018, 50, 1–25. [Google Scholar] [CrossRef]
- Bellomo, N.; Outada, N.; Soler, J.; Tao, Y.; Winkler, M. Chemotaxis and cross diffusion models in complex environments: Modeling towards a multiscale vision. Math. Model. Methods Appl. Sci. 2022, 32, 713–792. [Google Scholar] [CrossRef]
- Gibelli, L.; Elaiw, A.M.; Alghamdi, M.A.; Althiabi, A.M. Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics. Math. Model. Methods Appl. Sci. 2017, 27, 617–640. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Source | Parameter | Value | Source | Parameter | Value | Source |
|---|---|---|---|---|---|---|---|---|
| 10 | [18,50] | [11,51] | 2 | [32] | ||||
| [12,13] | [12,13] | [17,32] | ||||||
| [6,7] | 6 | [52] | h | [46] | ||||
| [32,52] | 6 | [52] | , , | Varied | Assumed |
| Case | Steady State | Stability | |
|---|---|---|---|
| Case (I) | |||
| Case (II) | |||
| Case (III) | |||
| Case (IV) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elaiw, A.M.; Aljahdali, A.K.; Hobiny, A.D. Dynamical Properties of Discrete-Time HTLV-I and HIV-1 within-Host Coinfection Model. Axioms 2023, 12, 201. https://doi.org/10.3390/axioms12020201
Elaiw AM, Aljahdali AK, Hobiny AD. Dynamical Properties of Discrete-Time HTLV-I and HIV-1 within-Host Coinfection Model. Axioms. 2023; 12(2):201. https://doi.org/10.3390/axioms12020201
Chicago/Turabian StyleElaiw, Ahmed M., Abdulaziz K. Aljahdali, and Aatef D. Hobiny. 2023. "Dynamical Properties of Discrete-Time HTLV-I and HIV-1 within-Host Coinfection Model" Axioms 12, no. 2: 201. https://doi.org/10.3390/axioms12020201
APA StyleElaiw, A. M., Aljahdali, A. K., & Hobiny, A. D. (2023). Dynamical Properties of Discrete-Time HTLV-I and HIV-1 within-Host Coinfection Model. Axioms, 12(2), 201. https://doi.org/10.3390/axioms12020201