

Stability and Hopf Bifurcation Analysis for a Phage Therapy Model with and without Time Delay

Abstract
:1. Introduction
2. Dynamics of the Non-Delayed Model
2.1. Positivity and Boundedness
2.2. Existence of Equilibrium Points
2.3. Stability Analysis
2.3.1. Stability Analysis of
- (i)
- The equilibrium is locally asymptotically stable if .
- (ii)
- If the parameter r reaches the transcritical threshold , a transcritical bifurcation arises around for System (2).
2.3.2. Stability Analysis of
- (i)
- The phage-free equilibrium is locally asymptotically stable if
- (ii)
- The equilibrium is globally asymptotically stable in the interior of the first quadrant of the plane.
2.3.3. Stability and Hopf Bifurcation of
2.3.4. Non-Existence of Non-Trivial Periodic Solution of System (2)
2.3.5. Global Stability of
3. Dynamics of the Delayed Model
3.1. Positivity and Boundedness
3.2. Stability Analysis
3.3. Hopf Bifurcation Analysis
3.4. Direction and Stability of Hopf-Bifurcating Periodic Solution
- (a)
- The Hopf bifurcation is supercritical (subcritical) if .
- (b)
- The bifurcating periodic solutions are stable (unstable) if .
- (c)
- The period of the bifurcated periodic solution increases (decreases) if .
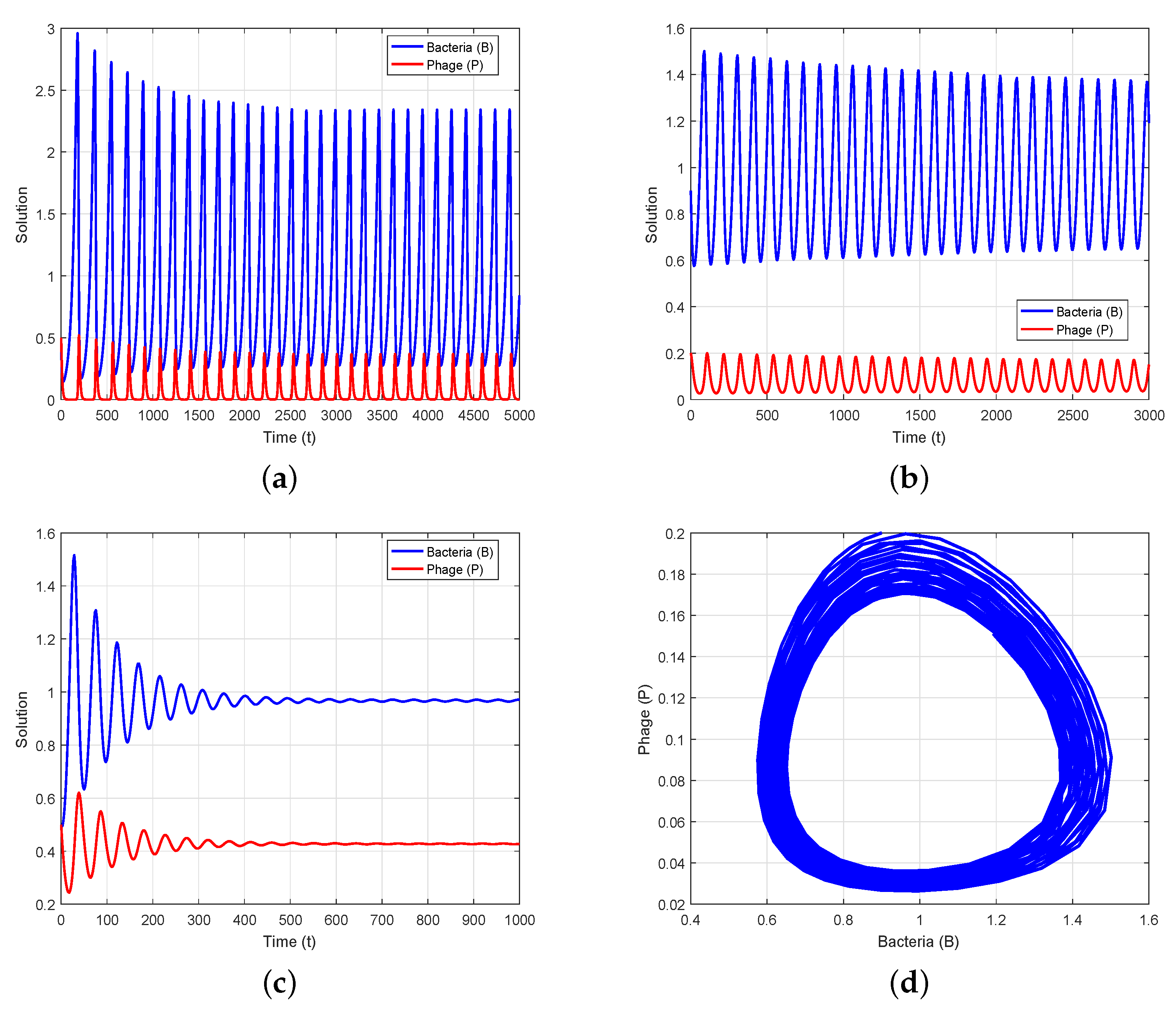
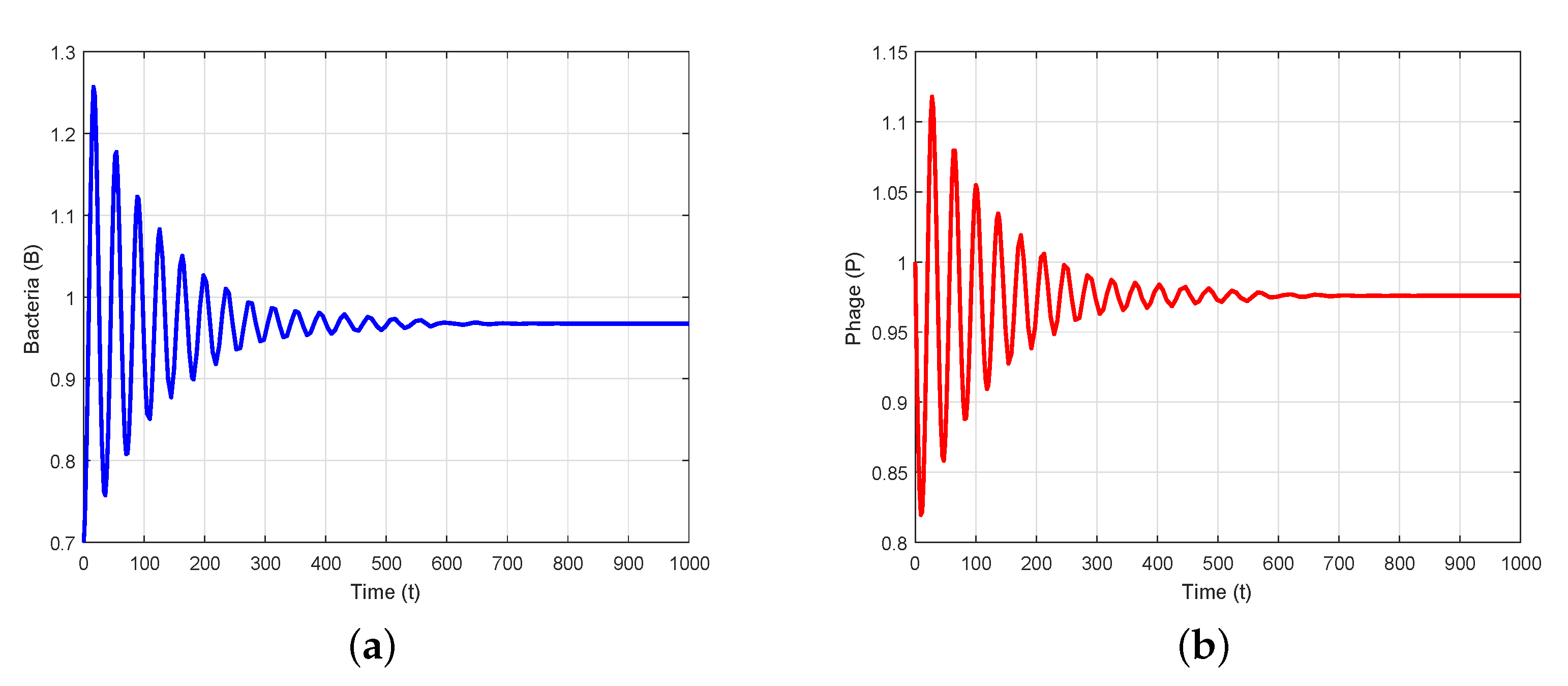
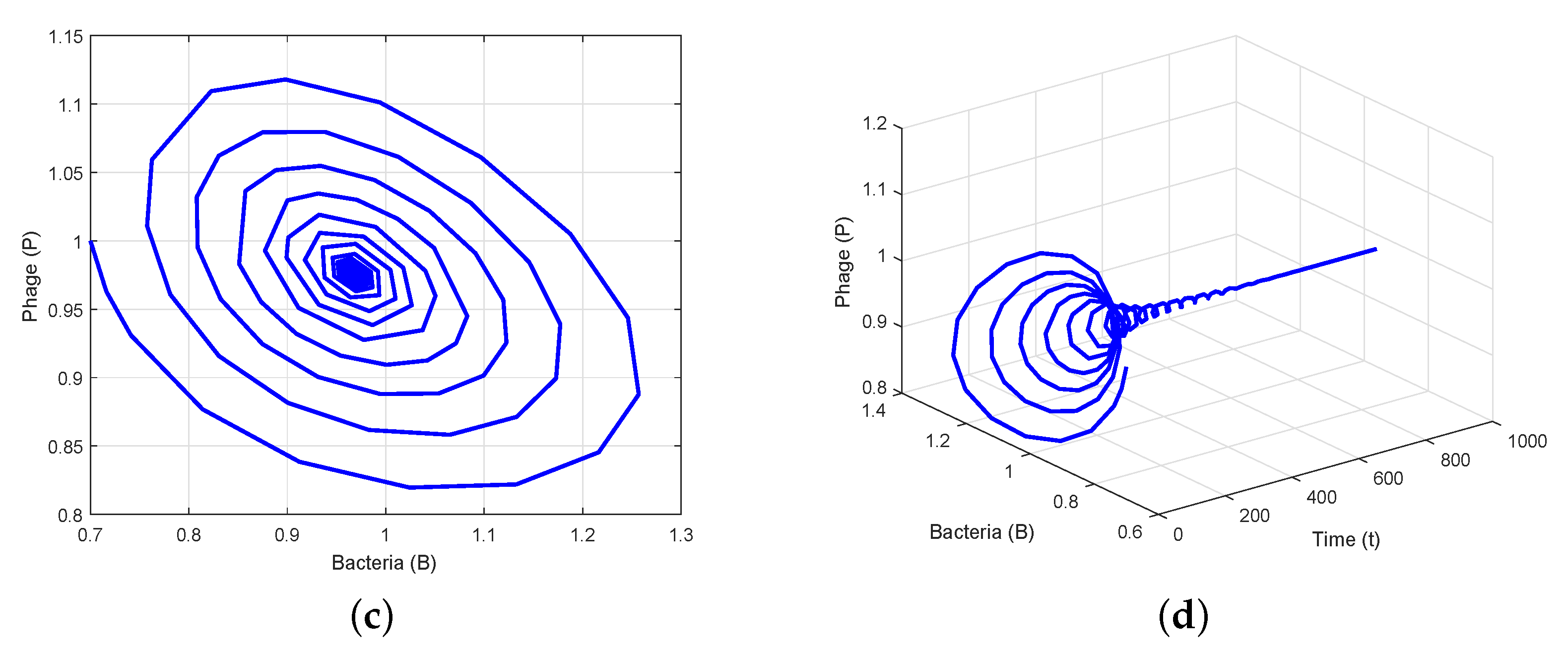
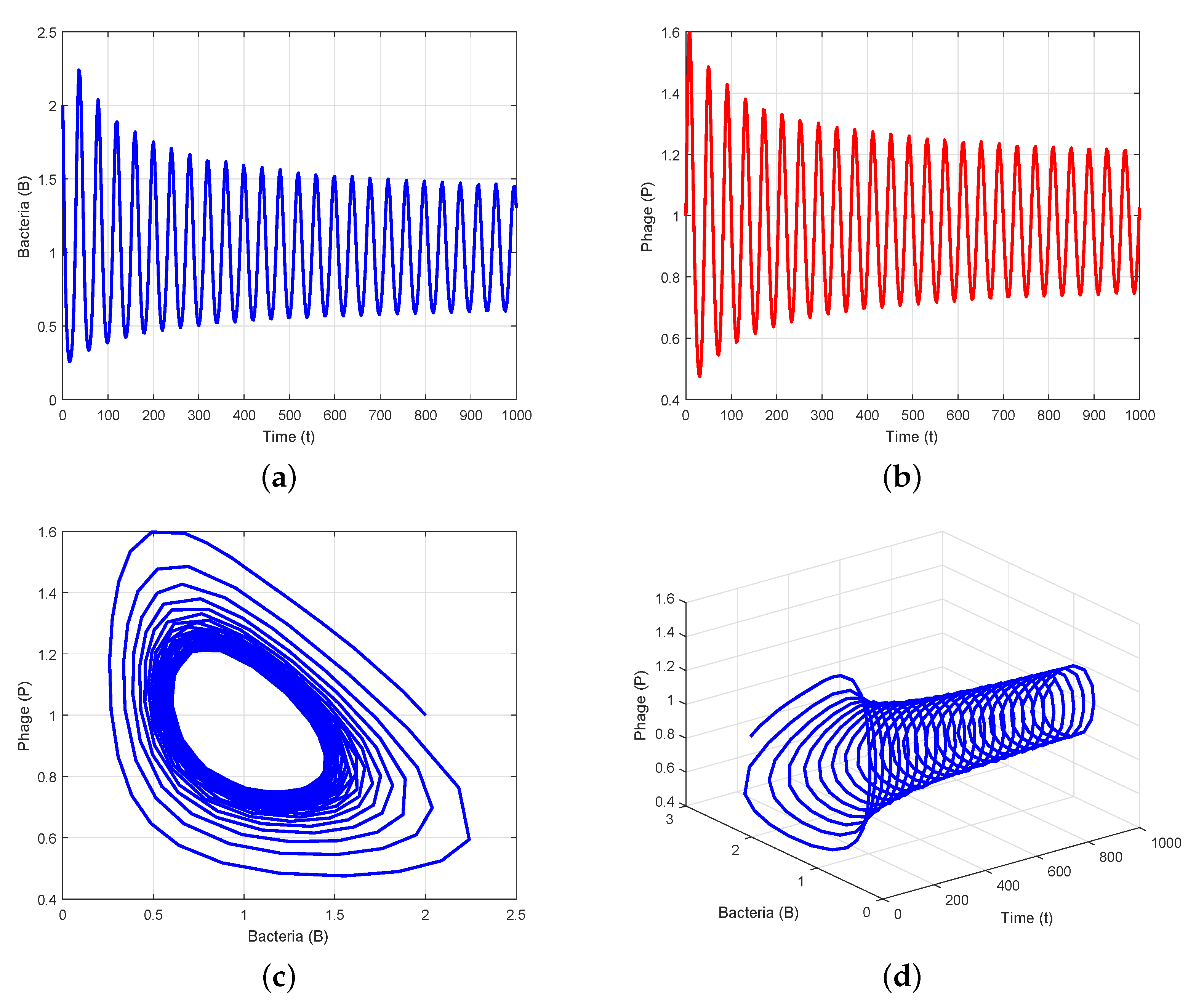
4. Numerical Simulation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beke, G.; Stano, M.; Klucar, L. Modeling the interaction between bacteriophages and their bacterial hosts. Math. Biosci. 2016, 279, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Beretta, E.; Kuang, Y. Modeling and analysis of a marine bacteriophage infection. Math. Biosci. 1998, 149, 57–76. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Grewal, R.K.; Roy, S. Modeling bacteria-phage interactions and its implications for phage therapy. Adv. Appl. Microbiol. 2018, 103, 103–141. [Google Scholar]
- Brives, C.; Pourraz, J. Phage therapy as a potential solution in the fight against AMR: Obstacles and possible futures. Palgrave Commun. 2020, 6, 100. [Google Scholar] [CrossRef]
- Styles, K.M.; Brown, A.T.; Sagona, A.P. A review of using mathematical modeling to improve our understanding of bacteriophage, bacteria, and eukaryotic interactions. Front. Microbiol. 2021, 2021, 2752. [Google Scholar] [CrossRef]
- Al-Darabsah, I. Time Delayed Models in Population Biology and Epidemiology. Ph.D. Dissertation, Memorial University of Newfoundland, St. John’s, NL, Canada, 2018. [Google Scholar]
- Campbell, A. Conditions for the existence of bacteriophage. Evolution 1961, 15, 153–165. [Google Scholar] [CrossRef]
- Levin, B.R.; Stewart, F.M.; Chao, L. Resource-limited growth, competition, and predation: A model and experimental studies with bacteria and bacteriophage. Am. Nat. 1977, 111, 3–24. [Google Scholar] [CrossRef]
- Lenski, R.E.; Levin, B.R. Constraints on the coevolution of bacteria and virulent phage: A model, some experiments, and predictions for natural communities. Am. Nat. 1985, 125, 585–602. [Google Scholar] [CrossRef]
- Smith, H.L.; Trevino, R.T. Bacteriophage infection dynamics: Multiple host binding sites. Math. Model. Nat. Phenom. 2009, 4, 109–134. [Google Scholar] [CrossRef]
- Sahani, S.K.; Gakkhar, S. A mathematical model for phage therapy with impulsive phage dose. Differ. Equ. Dyn. Syst. 2020, 28, 75–86. [Google Scholar] [CrossRef]
- Smith, H.L. Models of virulent phage growth with application to phage therapy. SIAM J. Appl. Math. 2008, 68, 1717–1737. [Google Scholar] [CrossRef]
- Misra, A.K.; Gupta, A.; Venturino, E. Cholera dynamics with bacteriophage infection: A mathematical study. Chaos Solitons Fract. 2016, 91, 610–621. [Google Scholar] [CrossRef]
- Teytsa, H.M.N.; Tsanou, B.; Bowong, S.; Lubuma, J.M. Bifurcation analysis of a phage-bacteria interaction model with prophage induction. Math. Med. Biol. 2021, 38, 28–58. [Google Scholar] [CrossRef]
- Li, X.; Huang, R.; He, M. Dynamics model analysis of bacteriophage infection of bacteria. Adv. Differ. Equ. 2021, 488, 3–11. [Google Scholar] [CrossRef]
- Xu, C.; Tang, X.; Liao, M. Stability and bifurcation analysis of a delayed predator-prey model of prey dispersal in two-patch environments. Appl. Math. Comput. 2010, 216, 2920–2936. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, R. Stability and bifurcation of a Cohen-Grossberg neural network with discrete delays. Appl. Math. Comput. 2011, 218, 2850–2862. [Google Scholar] [CrossRef]
- Li, Y.; Li, C. Stability and Hopf bifurcation analysis on a delayed Leslie–Gower predator-prey system incorporating a prey refuge. Appl. Math. Comput. 2013, 219, 4576–4589. [Google Scholar] [CrossRef]
- Khajanchi, S.; Banerjee, S. Stability and bifurcation analysis of delay induced tumor immune interaction model. Appl. Math. Comput. 2014, 248, 652–671. [Google Scholar] [CrossRef]
- Beretta, E.; Kuang, Y. Modeling and analysis of a marine bacteriophage infection with latency period. Nonlinear Anal. Real World Appl. 2001, 2, 35–74. [Google Scholar] [CrossRef]
- Beretta, E.; Solimano, F. The effect of time delay on stability in a bacteria-bacteriophage model. Sci. Math. Jpn. 2003, 58, 399–405. [Google Scholar]
- Liu, S.; Liu, Z.; Tang, J. A delayed marine bacteriophage infection model. Appl. Math. Lett. 2007, 20, 702–706. [Google Scholar] [CrossRef]
- Gakkhar, S.; Sahani, S.K. A time delay model for bacteria bacteriophage interaction. J. Biol. Syst. 2008, 16, 445–461. [Google Scholar] [CrossRef]
- Smith, H.L.; Thieme, H.R. Persistence of bacteria and phages in a chemostat. J. Math. Biol. 2012, 64, 951–979. [Google Scholar] [CrossRef]
- Aviram, I.; Rabinovitch, A. Bifurcation analysis of bacteria and bacteriophage coexistence in the presence of bacterial debris. Commun. Nonlinear Sci. Numer. Simul. 2012, 17, 242–254. [Google Scholar] [CrossRef]
- Calsina, A.; Palmada, J.M.; Ripoll, J. Optimal latent period in a bacteriophage population model structured by infection-age. Math. Model. Methods Appl. Sci. 2011, 21, 693–718. [Google Scholar] [CrossRef]
- Aviram, I.; Rabinovitch, A. Bacteria and lytic phage coexistence in a chemostat with periodic nutrient supply. Bull. Math. Biol. 2014, 76, 225–244. [Google Scholar] [CrossRef]
- Han, Z.; Smith, H.L. Bacteriophage-resistant and bacteriophage-sensitive bacteria in a chemostat. Math. Biosci. Eng. 2012, 9, 737–765. [Google Scholar]
- Beretta, E.; Solimano, F.; Tang, Y.B. Analysis of a chemostat model for bacteria and virulent bacteriaphage. Discrete Cont. Dyn. Syst. Ser. B 2002, 2, 495–520. [Google Scholar]
- Beretta, E.; Sakakibara, H.; Takeuchi, Y. Stability analysis of time delayed chemostat models for bacteria and virulent phage. Dyn. Syst. Their Appl. Biol. 2003, 36, 45–58. [Google Scholar]
- Czárxaxn, T.; Rattray, F.P.; Möller, C.O.A.; Christensen, B.B. Modelling the influence of metabolite diffusion on non-starter lactic acid bacteria growth in ripening cheddar cheese. Int. Dairy J. 2018, 80, 35–45. [Google Scholar]
- Wang, J.; Zheng, H.; Jia, Y. Dynamical analysis on a bacteria-phages model with delay and diffusion. Chaos Solitons Fract. 2021, 143, 110597. [Google Scholar] [CrossRef]
- Gourley, S.A.; Kuang, Y. A delay reaction-diffusion model of the spread of bacteriophage infection. SIAM J. Appl. Math. 2005, 65, 550–566. [Google Scholar] [CrossRef]
- Carletti, M. On the stability properties of a stochastic model for phage bacteria interaction in open marine environment. Math. Biosci. 2002, 175, 117–131. [Google Scholar] [CrossRef]
- Carletti, M. Mean-square stability of a stochastic model for bacteriophage infection with time delays. Math. Biosci. 2007, 210, 395–414. [Google Scholar] [CrossRef]
- Bardina, X.; Bascompte, D.; Rovira, C.; Tindel, S. An analysis of stochastic model for bacteriophage systems. Math. Biosci. 2013, 241, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Vidurupola, S.W.; Allen, L.J.S. Impact of Variability in Stochastic Models of Bacteria-Phage Dynamics Applicable to Phage Therapy. Stoch. Anal. Appl. 2014, 32, 427–449. [Google Scholar] [CrossRef]
- Vidurupola, S.W. Analysis of deterministic and stochastic mathematical models with resistant bacteria and bacteria debris for bacteriophage dynamics. Appl. Math. Comput. 2018, 316, 215–228. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef]
- Cisek, A.A.; Dabrowska, I.; Gregorczyk, K.P.; Wyzewski, Z. Phage therapy in bacterial infections treatment: One hundred years after the discovery of bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef]
- Leung, C.Y.; Weitz, J.S. Modeling the synergistic elimination of bacteria by phage and the innate immune system. J. Theor. Biol. 2017, 429, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Kyaw, E.E.; Zheng, H.; Wang, J.; Hlaing, H.K. Stability analysis and persistence of a phage therapy model. Math. Biosci. Eng. 2021, 18, 5552–5572. [Google Scholar] [CrossRef] [PubMed]
- Birkhoff, G.; Rota, G.C. Ordinary Differential Equations; John Wiley and Sons: New York, NY, USA, 1982. [Google Scholar]
- Tripathi, J.P.; Tyagi, S.; Abbas, S. Global analysis of a delayed density dependent predator-prey model with Crowley-Martin functional response. Commun. Nonlinear Sci. Numer. Simul. 2016, 30, 45–69. [Google Scholar] [CrossRef]
- Perko, L. Differential Equations and Dynamical Systems; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Freedman, H.I.; Rao, V.S.H. The trade-off between mutual interference and time lags in predator-prey ststems. Bull. Math. Biol. 1983, 45, 991–1004. [Google Scholar] [CrossRef]
- Wiggins, S. Introduction to Applied Nonlinear Dynamical Systems and Chaos; Springer: New York, NY, USA, 1990. [Google Scholar]
- Kuang, Y. Delay Differential Equation with Applications in Population Dynamics; Academic Press: New York, NY, USA, 1993. [Google Scholar]
- Hassard, B.D.; Kazarinoff, N.D.; Wan, Y.H. Theory and Applications of Hopf Bifurcation; University Cambridge: Cambridge, UK, 1981. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Data 1 | Data 2 |
|---|---|---|---|
| adsorption rate of phage | 0.34 | 0.34 | |
| burst size of phage | 0.38 | 0.38 | |
| killing rate of innate immune response | 0.19 | 0.19 | |
| w | decay rate of phage | 0.125 | 0.125 |
| r | intrinsic growth rate of bacteria | 0.25 | 0.5 |
| carrying capacity of bacteria | 7.29 | 5 | |
| bacterial concentration when innate immune | |||
| response is half saturated | 3.5 | 3.5 | |
| carrying capacity of innate immune response | 0.48 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyaw, E.E.; Zheng, H.; Wang, J. Stability and Hopf Bifurcation Analysis for a Phage Therapy Model with and without Time Delay. Axioms 2023, 12, 772. https://doi.org/10.3390/axioms12080772
Kyaw EE, Zheng H, Wang J. Stability and Hopf Bifurcation Analysis for a Phage Therapy Model with and without Time Delay. Axioms. 2023; 12(8):772. https://doi.org/10.3390/axioms12080772
Chicago/Turabian StyleKyaw, Ei Ei, Hongchan Zheng, and Jingjing Wang. 2023. "Stability and Hopf Bifurcation Analysis for a Phage Therapy Model with and without Time Delay" Axioms 12, no. 8: 772. https://doi.org/10.3390/axioms12080772