
Novel Directions for Neuromorphic Machine Intelligence Guided by Functional Connectivity: A Review


Abstract
:1. Introduction
- We provide an accessible resource for researchers in the related fields of neuroscience and computer science that explains FC in terms of network organisation, multi-tasking, and learning methods.
- This is the first review, to our knowledge, that gives a comprehensive background into key areas of neuroscience to give novel directions for neuromorphic machine intelligence.
- We discuss key neuroimaging techniques, giving computer scientists an insight into the types of data available for consideration in the development of neuromorphic AI.
- The review also refers to key application domains, such as human augmentation that will benefit from FC-inspired neural network design and artificial learning.
Organisation of the Review
- Section 2: We begin by providing a background into key concepts in neuroscience and AI that are referred to throughout this review. We provide descriptive definitions for FC, executive function, and neuromorphic AI. In addition, we discuss popular neuroimaging techniques that are being used in both domains and how these have been used in combination to understand FC.
- Section 3: We then look at how FC can inspire artificial network design and the potentials of considering multi-network integration for computing systems. Modelling AI systems from known FC networks may be a step toward making AI explainable and more human in decision-making.
- Section 4: We then discuss learning in humans and the role of FC in memory retrieval. After also discussing various learning paradigms in AI, we suggest research directions for improving the efficiency and memory capabilities of neuromorphic artificial learning.
- Section 5: The next section covers the dilemmas faced by a robot trying to cook pasta. We discuss the human ability to multi-task and allocate attention in response to an unexpected stimulus and how FC could aid in areas of research relating to temporal abstraction, transfer learning, and swarm intelligence.
- Section 6: Potential applications for NMAI: we discuss areas around non-invasive neurotechnology, human augmentation, and collaborative robots across different domains.
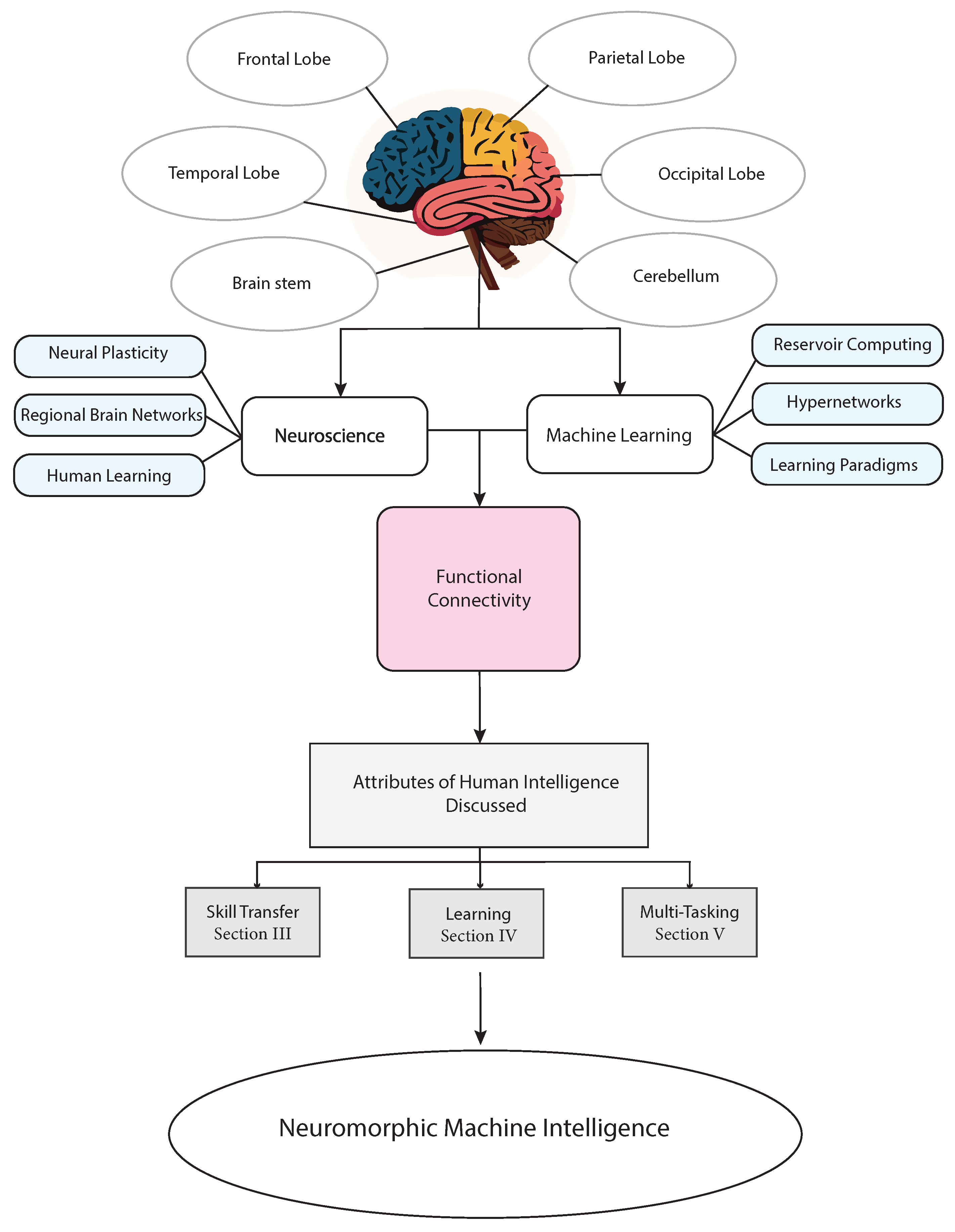
2. Background
2.1. Explanations of Key Concepts in Neuroscience for This Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Network (Abbreviation) | Primary Brain Regions | Role They Play | Section |
|---|---|---|---|
| Frontoparietal Network (FPN) |
| Executive functions such as working memory, goal-oriented cognition, inhibition, and task-switching. | Section 3, Section 4 and Section 5 |
| Dorsal Attention Network (DAN) |
| Typically involved in visuo-spatial attention and attention allocation. | Section 5 |
| Salience Network (SN) |
| This network is involved in determining if information is important or not. Furthermore, it plays a role in individualistic goal-directed behaviour. | Section 5 |
| Default Mode Network (DMN) |
| This network can be seen as active during cognitive functions that are focused on internal conditions rather than in response to the external environment. It plays a role in semantic cognition and language comprehension, as well as conscious thought and decision-making. | Section 3, Section 4 and Section 5 |

2.1.1. Defining Functional Connectivity
2.1.2. Executive Function and Cognitive Control
2.2. Key Concepts in Computer Science for this Review
2.2.1. Neuromorphic Artificial Intelligence and Synaptic Models
2.2.2. Neuromorphic Directions for Artificial Learning
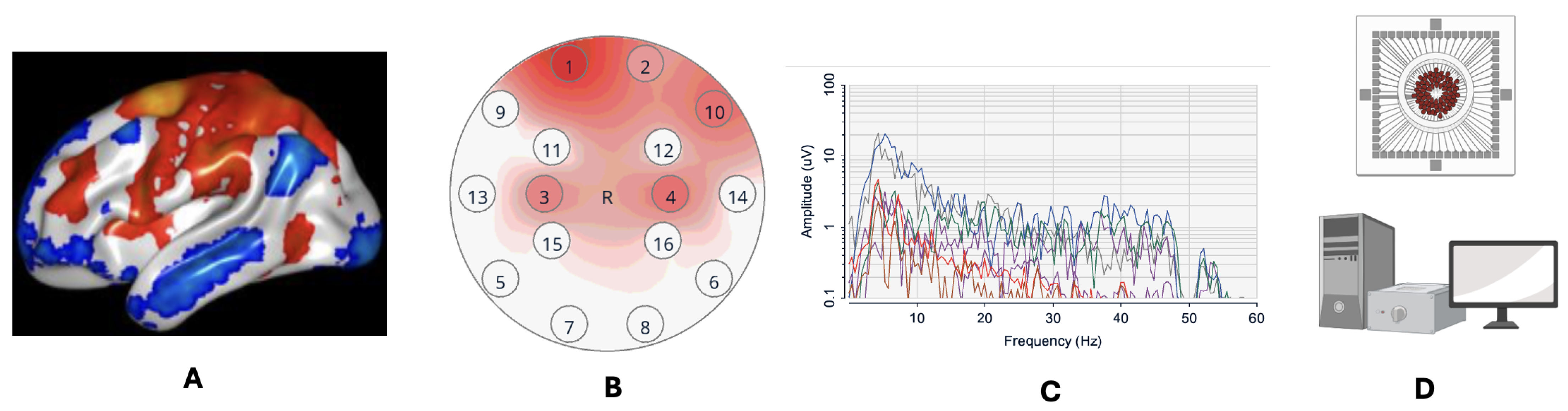
2.3. Neuroimaging Techniques Used in Observing FC
2.3.1. Functional Magnetic Resonance Imaging
2.3.2. Electroencephalography
2.3.3. Microelectrode Recording and Arrays
2.3.4. Combining EEG and fMRI to Predict Functional Brain States
3. Functional Network Design for Neuromorphic Machine Intelligence
3.1. Network Organisation and Information Storage in the Brain
3.1.1. Network Integration and Segregation for Executive Functions in Humans
3.1.2. Neural Plasticity in the Brain for Energy-Efficient Processing
3.1.3. Synaptic Information Storage Capacity Measured with Information Theory
3.1.4. Networks for Skill Transfer in Humans
3.1.5. Hypergraphs for Modelling Brain Connectivity from fMRI
3.1.6. Subsection Summary
3.2. Perceiving FC from a Deep Learning Perspective
3.2.1. Hypernetworks as Artificial Functional Networks
3.2.2. Hypernetworks for Continual Learning
3.2.3. Transfer Learning
3.2.4. Organising Organic Neural Networks for Neuromorphic Machine Intelligence
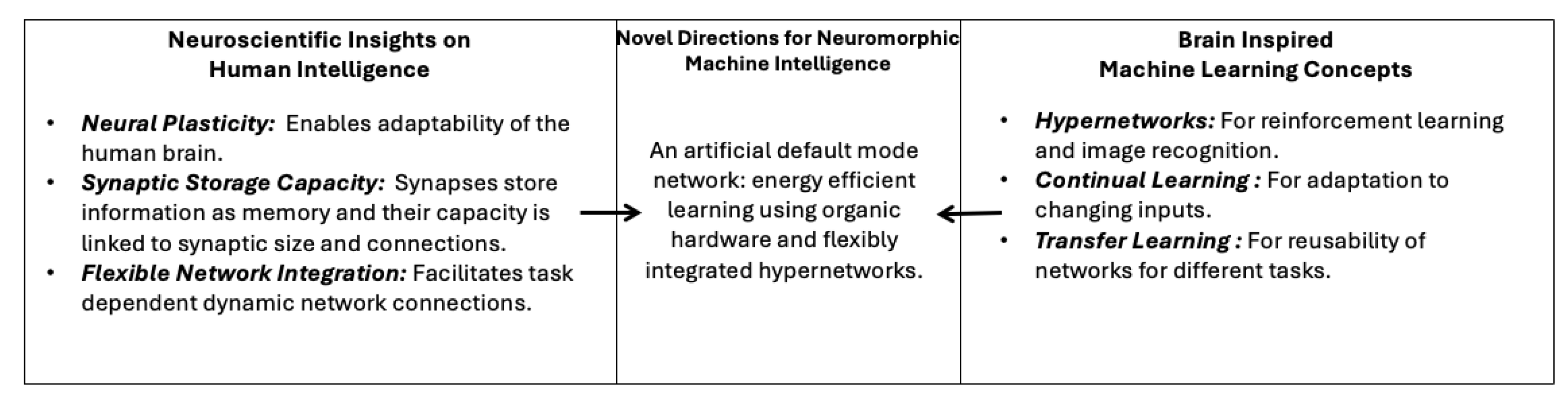
3.3. Research-Inspired Directions for Neuromorphic Machine Intelligence
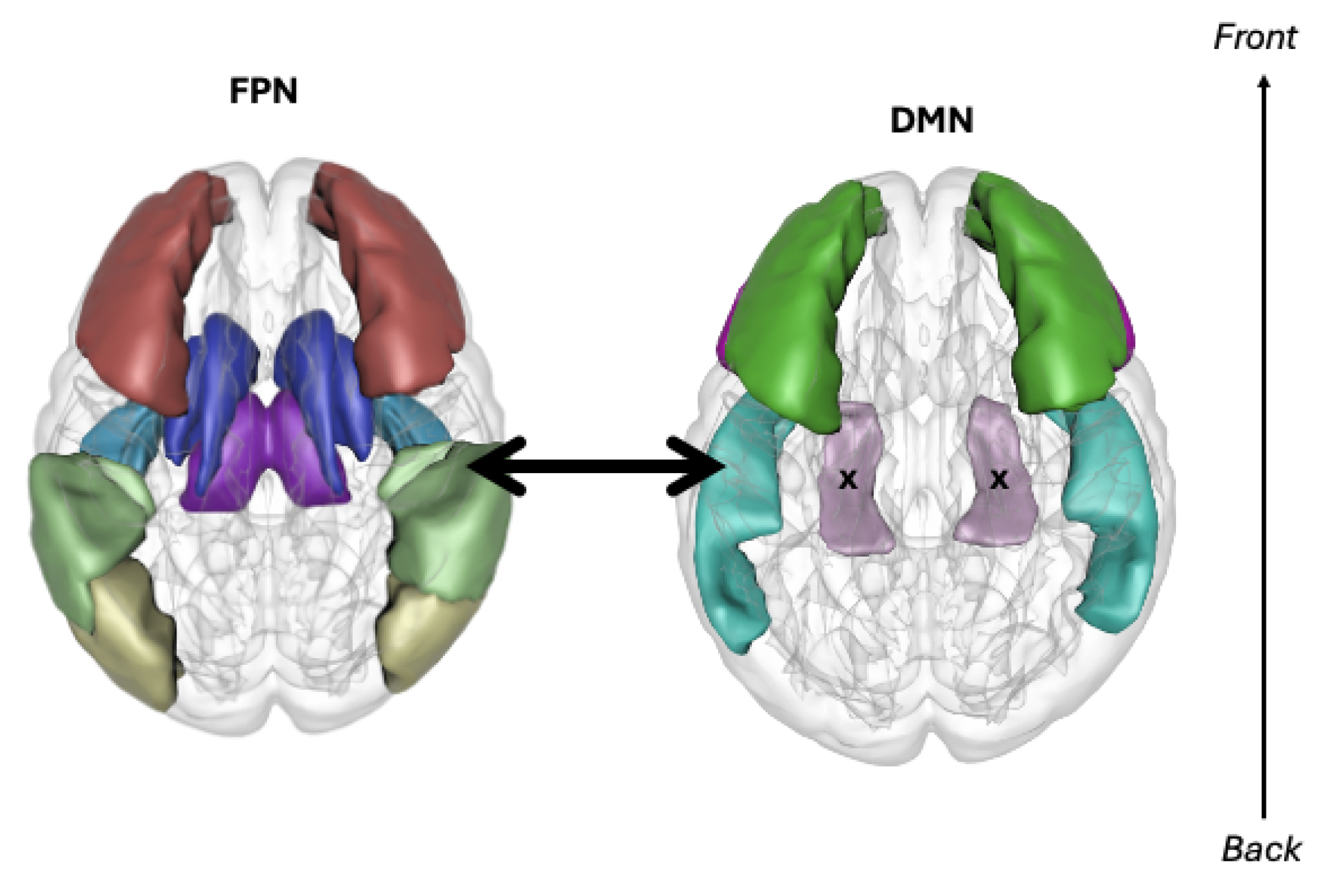
- Given the importance of the human DMN and its integration with other networks, a neuromorphic default mode network using a hypergraph structure could create a base for artificial architectures to be built upon.
- The use of brain organoid hardware in this hypergraph DMN may provide a sustainable path for energy-efficient neuromorphic systems, making the feasibility of NMI in the future coincide with environmental preservation goals in society.
4. Learning: The Key to Human-like Intelligence
- Errorless and trial-and-error learning in humans: FC dynamics facilitating learning in humans.
- Memory retrieval in humans and how FC contributes to this ability.
- Artificial learning methods such as reinforcement learning and brain-inspired methods of incorporating learning and memory into AI agents.
4.1. Learning in Humans
4.1.1. Errorless Learning and Trial-and-Error Learning
4.1.2. Human Memory Models
4.1.3. FC for Memory Retrieval
4.1.4. Subsection Summary
4.2. Current Brain-Inspired Machine Learning Concepts for Artificial Learning
4.2.1. Prospective Learning vs. Backpropagation for Artificial Learning
4.2.2. Artificial Learning Methods: Reinforcement Learning
4.3. Research-Inspired Directions for Neuromorphic Machine Intelligence
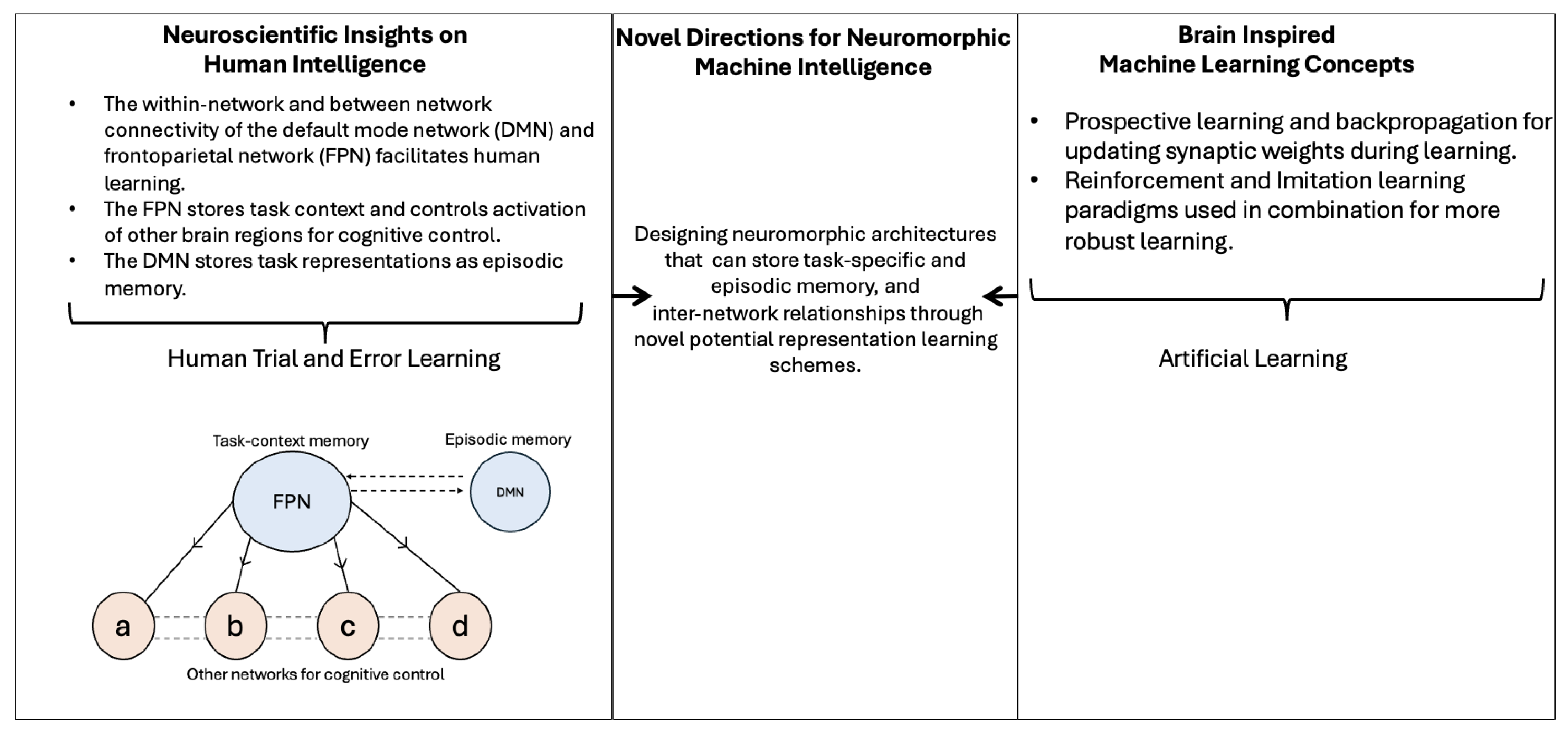
- A neuromorphic machine intelligence architecture with a task-specific context and episodic memory storage could limit the effects of catastrophic forgetting in continual learning systems.
- Facilitating inter-network relationships when designing neuromorphic models with multiple networks could give machines the ability to learn faster. This would lead to reduced model training times and less energy consumption for sustainable NMI.
5. The Pasta Problem: How FC Can Inspire Task-Prioritisation Systems
5.1. The Role of Different FC Networks in Human Multi-Tasking
5.1.1. Dorsal Attention Network: For Goal-Directed Attention to Environmental Stimuli
5.1.2. Salience Network: A Biological Switch for Identifying Relevant Stimuli
5.1.3. The Default Mode Network: The Introspective Mind at Rest
5.1.4. The Frontoparietal Network: For Task-Planning and Decision-Making
5.2. FC to Enable Task-Switching and Multi-Task Completion
- Attention allocation during multi-tasking.
- Performance evaluation during task completion.
- Mind wandering due to feeling overwhelmed by multiple stimuli.
- Imagining the task components in order to facilitate the state of attention allocation.
Subsection Summary
5.3. Areas within Machine Learning That Could Benefit from Attention and Task Prioritisation
5.3.1. Human-Inspired Multi-Task Learning
5.3.2. Multi-Task Reinforcement Learning for Generalisability
5.3.3. Salience Interest for Temporal Abstraction
5.3.4. The Need for Task-Prioritisation in Co-Operative Multi-Agent Swarms
5.4. Research-Inspired Directions for Neuromorphic Machine Intelligence
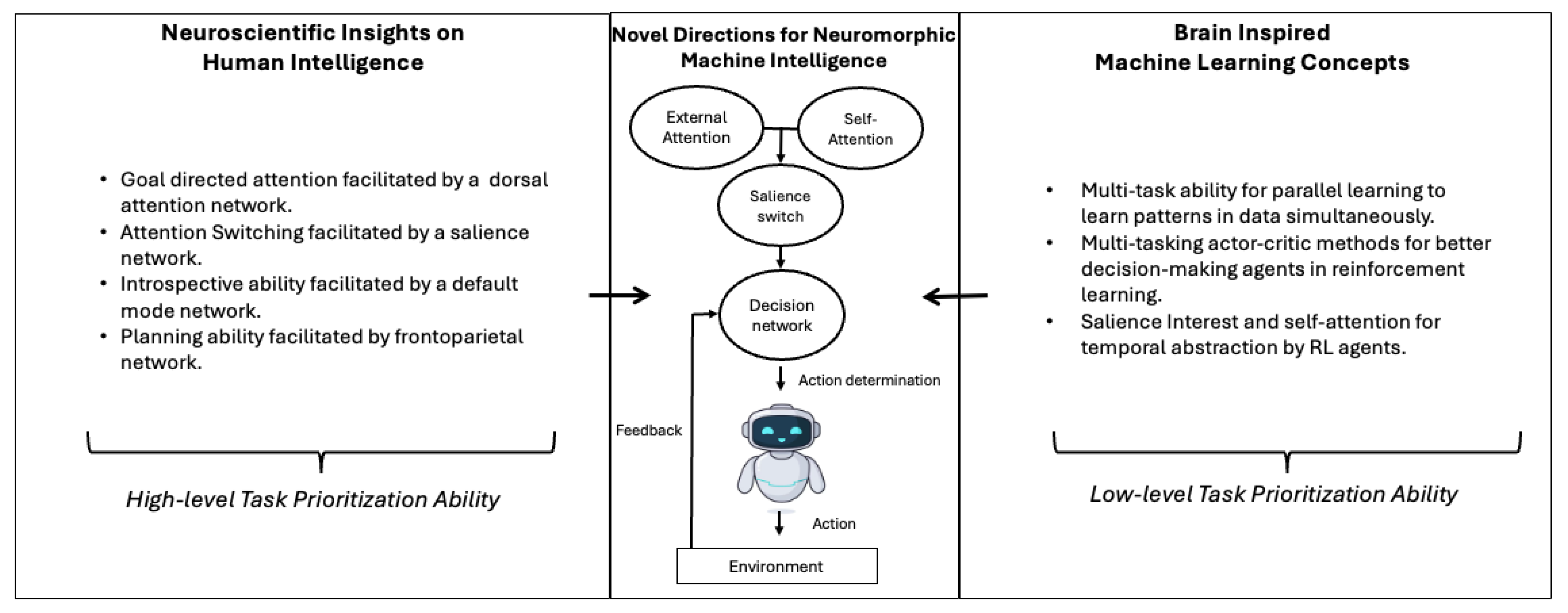
- Having an external attention mechanism to store task-specific context and a self-attention mechanism storing episodic memory, inspired by FPN and DMN, could give agents the ability to prioritise as intuitively as humans do whilst preserving task-related memory.
- For neuromorphic machines, a dedicated decision network could fit into this architecture, for the final decision-making ability based on the evaluation of any unexpected situations in the environment made by the salience switch.
6. Potential Application Domains for Neuromorphic Machine Intelligence
6.1. Autonomous Driverless Vehicles
6.2. Human-Compatible Assistive Neuroprosthetics
6.3. Development and Integration of Energy-Efficient Sensors
6.4. Collaborative Robots for Human–Robot Interaction
6.5. Robust Swarms of Autonomous Machines
6.6. Customisable Non-Invasive Human Augmentation Tools Using BCI
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Frenkel, C.; Bol, D.; Giacomo, I. Bottom-Up and Top-Down Neural Processing Systems Design: Neuromorphic Intelligence as the Convergence of Natural and Artificial Intelligence. IEEE Proc. 2023. [Google Scholar] [CrossRef]
- Eickhoff, S.; Muller, V. Functional Connectivity. Brain Mapp. 2015, 2, 187–201. [Google Scholar] [CrossRef]
- McIntosh, T.; Susnjak, T.; Liu, T.; Watters, P.; Ng, A.; Halgamuge, M. A Game-Theoretic Approach to Containing Artificial General Intelligence: Insights from Highly Autonomous Aggressive Malware. IEEE Trans. Artif. Intell. 2024. [Google Scholar] [CrossRef]
- Wan, Q.; Hu, S.; Zhang, Y.; Wang, P.; Wen, B.; Lu, Z. “It Felt Like Having a Second Mind”: Investigating Human-AI Co-creativity in Prewriting with Large Language Models. Proc. Acm -Hum.-Comput. Interact. 2024. [Google Scholar] [CrossRef]
- Rasnayaka, S.; Wang, G.; Shariffdeen, R.; Lyer, G. An Empirical Study on Usage and Perceptions of LLMs in a Software Engineering Project. In Proceedings of the 46th International Conference on Software Engineering, Lisbon, Portugal, 12–21 April 2024. [Google Scholar]
- Obrenovic, B.; Gu, X.; Wang, G.; Godinic, D.; Jakhongirov, I. Generative AI and human—Robot interaction: Implications and future agenda for business, society and ethics. AI Soc. 2024. [Google Scholar] [CrossRef]
- Cheng, B.; Lin, H.; Kong, Y. Challenge or hindrance? How and when organizational artificial intelligence adoption influences employee job crafting. J. Bus. Res. 2023, 164, 113987. [Google Scholar] [CrossRef]
- Oniani, D.; Hilsman, J.; Peng, Y.; Poropatich, R.; Pamplin, J.; Legault, G.; Wang, Y. Adopting and expanding ethical principles for generative artificial intelligence from military to healthcare. npj Digit. Med. 2023, 6, 225. [Google Scholar] [CrossRef]
- Wang, D.; Churchill, E.; Maes, P.; Fan, X.; Shneiderman, B.; Shi, Y.; Wang, Q. From Human-Human Collaboration to Human-AI Collaboration: Designing AI Systems That Can Work Together with People. In Proceedings of the Extended Abstracts of the 2020 CHI Conference on Human Factors in Computing Systems, CHI EA ’20, Honolulu, HI, USA, 25–30 April 2020; pp. 1–6. [Google Scholar] [CrossRef]
- Wu, C.J.; Raghavendra, R.; Gupta, U.; Acun, B.; Ardalani, N.; Maeng, K.; Chang, G.; Aga, F.; Huang, J.; Bai, C.; et al. Sustainable AI: Environmental Implications, Challenges and Opportunities. Proc. Mach. Learn. Syst. 2022, 4, 795–813. [Google Scholar]
- Macpherson, T.; Churchland, A.; Sejnowski, T.; DiCarlo, J.; Kamitani, Y.; Takahashi, H.; Hikida, T. Natural and Artificial Intelligence: A brief introduction to the interplay between AI and neuroscience research. Neural Netw. 2021, 144, 603–613. [Google Scholar] [CrossRef]
- Sutton, R.; Barto, A. Reinforcement Learning: An Introduction; MIT Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Cai, H.; Ao, Z.; Tian, C.; Wu, Z.; Liu, H.; Tchieu, J.; Gu, M.; Mackie, K.; Guo, F. Brain organoid reservoir computing for artificial intelligence. Nat. Electron. 2023, 6, 1032–1039. [Google Scholar] [CrossRef]
- Suarez, L.; Richards, B.; Lajoie, G.; Misic, B. Learning function from structure in neuromorphic networks. Nat. Mach. Intell. 2021, 3, 771–786. [Google Scholar] [CrossRef]
- Benisty, H.; Barson, D.; Moberly, A.; Lohani, S.; Tang, L.; Coifman, R.; Crair, M.; Mishne, G.; Cardin, J.; Higley, M. Rapid fluctuations in functional connectivity of cortical networks encode spontaneous behavior. Nat. Neurosci. 2024, 27, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Maslow, A.H. A theory of human motivation. Psychol. Rev. 1943, 50, 370–396. [Google Scholar] [CrossRef]
- Rybnicek, R.; Bergner, S.; Gutschelhofer, A. How individual needs influence motivation effects: A neuroscientific study on McClelland’s need theory. Rev. Manag. Sci. 2019, 13, 443–482. [Google Scholar] [CrossRef]
- Demir, M.; McNeese, N.J.; Cooke, N.J. The Impact of Perceived Autonomous Agents on Dynamic Team Behaviors. IEEE Trans. Emerg. Top. Comput. Intell. 2018, 2, 258–267. [Google Scholar] [CrossRef]
- Hartikainen, M.; Spurava, G.; Vaananen, K. Human-AI Collaboration in Smart Manufacturing: Key Concepts and Framework for Design. Front. Artif. Intell. Appl. 2024, 386, 162–172. [Google Scholar]
- La Fata, C.; Adelfio, L.; Micale, R.; La Scalia, G. Human error contribution to accidents in the manufacturing sector: A structured approach to evaluate the interdependence among performance shaping factors. Saf. Sci. 2023, 161, 106067. [Google Scholar] [CrossRef]
- Hassija, V.; Chamola, V.; Mahapatra, A.; Singal, A.; Goel, D.; Huang, K.; Scardapane, S.; Spinelli, I.; Mahmud, M.; Hussain, A. Interpreting Black-Box Models: A Review on Explainable Artificial Intelligence. Cogn. Comput. 2024, 16, 45–74. [Google Scholar] [CrossRef]
- Pina, R.; Artaud, C.; Liu, X.; De Silva, V. Staged Reinforcement Learning for Complex Tasks Through Decomposed Environments. Intell. Syst. Pattern Recognit. 2023. [Google Scholar]
- Kaur, S.; Singla, J.; Nkenyereye, L.; Jha, S.; Prashar, D.; Prasad, G. Medical Diagnostic Systems Using Artificial Intelligence (AI) Algorithms: Principles and Perspectives. IEEE Access 2020. [Google Scholar] [CrossRef]
- Luccioni, S.; Jernite, Y.; Strubell, E. Power Hungry Processing: Watts Driving the Cost of AI Deployment? In Proceedings of the 2024 ACM Conference on Fairness, Accountability, and Transparency, FAccT ’24, Rio de Janeiro, Brazil, 3–6 June 2024; pp. 85–99. [Google Scholar] [CrossRef]
- EPA. United States Environmental Protection Agency Green—House Gas Equivalencies Calculator; EPA: New York, NY, USA, 2021.
- Greif, L.; Kimmig, A.; Bobbou, S.; Jurisch, P.; Ovtcharova, J. Strategic view on the current role of AI in advancing environmental sustainability: A SWOT analysis. Discov. Artif. Intell. 2024, 4, 45. [Google Scholar] [CrossRef]
- Wu, Y.; Moon, J.; Zhu, X.; Lu, W. Neural Functional Connectivity Reconstruction with Second-Order Memristor Network. Adv. Intell. Syst. 2021, 3, 2000276. [Google Scholar] [CrossRef]
- Uddin, L. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Uddin, L.; Yeo, T.; Spreng, N. Towards a Universal Taxonomy of Macro-scale Functional Human Brain Networks. Brain Topogr. 2019, 32, 926–942. [Google Scholar] [CrossRef]
- Fan, L.; Li, H.; Zhuo, J.; Zhang, Y.; Wang, J.; Chen, L.; Yang, Z.; Chu, C.; Xie, S.; Laird, A.; et al. The Human Brainnetome Atlas: A New Brain Atlas Based on Connectional Architecture. Cereb. Cortex 2016, 26, 3508–3526. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; Psychology Press: London, UK, 2005. [Google Scholar]
- Eickhoff, S.B.; Jbabdi, S.; Caspers, S.; Laird, A.R.; Fox, P.T.; Zilles, K.; Behrens, T.E.J. Anatomical and functional connectivity of cytoarchitectonic areas within the human parietal operculum. J. Neurosci. 2010, 30, 6409–6421. [Google Scholar] [CrossRef]
- Bear, M.; Connors, B.W.; Paradiso, M. Neuroscience: Exploring the Brain, 3rd ed.; Lippincott Williams & Wilkins Publishers: Philadelphia, PA, USA, 2007; pp. 293–331. [Google Scholar]
- Javed, K.; Reddy, V.; Lui, F. Neuroanatomy, Cerebral Cortex; StatPearls: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK537247/ (accessed on 26 March 2024).
- Diamond, A. Executive Functions. Annu Rev Psychol. 2013, 64, 35–168. [Google Scholar] [CrossRef] [PubMed]
- Vereshchaka, A.; Yang, F.; Suresh, A.; Olokodana, I.L.; Dong, W. Predicting Cognitive Control in Older Adults Using Deep Learning and EEG Data. In Proceedings of the 2020 International Conference on Social Computing, Behavioral-Cultural Modeling & Prediction and Behavior Representation in Modeling and Simulation (SBP-BRiMS 2020), Washington, DC, USA, 19–22 October 2020; pp. 19–22. [Google Scholar]
- Miranda, E.; Sune, J. Memristors for Neuromorphic Circuits and Artificial Intelligence Applications. Materials 2020, 13, 938. [Google Scholar] [CrossRef]
- Bile, A.; Tari, H.; Pepino, R.; Nabizada, A.; Fazio, E. Solitonic Neural Network: A novel approach of Photonic Artificial Intelligence based on photorefractive solitonic waveguides. In Proceedings of the EPJ Web of Conferences. EDP Sciences, Kaifeng, China, 21–23 April 2023; Volume 287, p. 13003. [Google Scholar]
- Yaremkevich, D.D.; Scherbakov, A.V.; De Clerk, L.; Kukhtaruk, S.M.; Nadzeyka, A.; Campion, R.; Rushforth, A.W.; Savel’ev, S.; Balanov, A.G.; Bayer, M. On-chip phonon-magnon reservoir for neuromorphic computing. Nat. Commun. 2023, 14, 8296. [Google Scholar] [CrossRef]
- Wilson, H.; Cowan, J. Excitatory and Inhibitory Interac- tions in Localized Populations of Model Neurons. Biophys. J. 1972, 12, 1–24. [Google Scholar] [CrossRef]
- Gerstner, W.; Kistler, W.M. Spiking Neuron Models: Single Neurons, Populations, Plasticity; Cambridge University: Cambridge, UK, 2002. [Google Scholar]
- Yamazaki, K.; Vo-Ho, V.K.; Bulsara, D.; Le, N. Spiking Neural Networks and Their Applications: A Review. Brain Sci. 2022, 12, 863. [Google Scholar] [CrossRef]
- Menon, V.; D’Esposito, M. The role of PFC networks in cognitive control and executive function. Neuropsychopharmacology 2022, 47, 90–103. [Google Scholar] [CrossRef]
- Hilman, E. Coupling Mechanism and Significance of the BOLD Signal: A Status Report. Annu. Rev. Neurosci. 2014, 37, 161–181. [Google Scholar] [CrossRef] [PubMed]
- Matthews, P.M.; Jezzard, P. Functional Magnetic Resonance Imaging. J. Neurol. Neurosurg. Psychiatry 2004, 75, 6–12. [Google Scholar]
- Scrivener, C.L.; Reader, A.T. Variability of EEG electrode positions and their underlying brain regions: Visualizing gel artifacts from a simultaneous EEG-fMRI dataset. Brain Behav. 2022, 12, e2476. [Google Scholar] [CrossRef]
- Schomer, D.; Silva, F.D. Niedermeyer’s Electroencephalography; Oxford University Press: Oxford, UK, 2017; Volume 1. [Google Scholar] [CrossRef]
- Mohammedi, M.; Omar, M.; Bouabdallah, A. Methods for detecting and removing ocular artifacts from EEG signals in drowsy driving warning systems: A survey. Multimed. Tools Appl. 2023, 82, 17687–17714. [Google Scholar] [CrossRef]
- Seyedkhani, S.; Mohammadpour, R.; Irajizad, A. Principles and Advancements of Microelectrode Arrays in Brain–Machine Interfaces; Intechopen: London, UK, 2024. [Google Scholar] [CrossRef]
- Bradley, J.; Luithardt, H.; Metea, M.; Stock, C.J. In vitro screening for seizure liability using microelectrode array technology. Toxicol. Sci. 2018, 163, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.; Rolston, J.; Potter, S. How to culture, record and stimulate neuronal networks on micro-electrode arrays (MEAs). J. Vis. Exp. 2010, 39, e2056. [Google Scholar] [CrossRef]
- Maccione, A.; Garofalo, M.; Nieus, T.; Tedesco, M.; Berdondini, L.; Martinoia, S. Multiscale functional connectivity estimation on low-density neuronal cultures recorded by high-density CMOS micro electrode arrays. J. Neurosci. Methods 2012, 207, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Viventi, J.; Kim, D.H.; Vigeland, L.; Frechette, E.; Blanco, J.; Kim, Y.S. Flexible, foldable, actively multiplexed, high-density electrode array for mapping brain activity in vivo. Nat. Neurosci. 2011, 14, 1599–1605. [Google Scholar] [CrossRef]
- Abreu, R.; Jorge, J.; Leal, A. EEG Microstates Predict Concurrent fMRI Dynamic Functional Connectivity States. Brain Topogr. 2021, 34, 41–55. [Google Scholar] [CrossRef]
- Cole, M.; Bassett, D.; Power, J.; Braver, T.; Petersen, S. Intrinsic and task-evoked network architectures of the human brain. Neuron 2014, 83, 238–251. [Google Scholar] [CrossRef]
- Ryali, S.; Supekar, K.; Chen, T.; Kockalka, J.; Cai, W.; Nicholas, J.; Padmanabhan, A.; Menon, V. Temporal Dynamics and Developmental Maturation of Salience, Default and Central-Executive Network Interactions Revealed by Variational Bayes Hidden Markov Modeling. PLoS Comput. Biol. 2016, 12, e1005138. [Google Scholar] [CrossRef]
- Yamada, T.; Watanabe, T.; Sasaki, Y. Plasticity—Stability dynamics during post-training processing of learning. Trends Cogn. Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Hebb, D. The Organization of Behaviour; John Wiley and Sons: Hoboken, NJ, USA, 1949. [Google Scholar]
- Ólafsdóttir, H.; Bush, D.; Barry, C. The role of Hippocampal Replay in Memory and Planning. Curr. Biol. 2018, 28, R37–R50. [Google Scholar] [CrossRef]
- Samavat, M.; Bartol, T.; Harris, K.; Sejnowski, T. Synaptic Information Storage Capacity Measured with Information Theory. Neural Comput. 2024, 36, 781–802. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Scripture, E.; Smith, T.; Brown, E. On the education of muscular control and power. Stud. Yale Psychol. Lab. 1894, 2, 114–119. Available online: http://echo.mpiwg-berlin.mpg.de/MPIWG:47EYCE88 (accessed on 15 August 2024).
- Ossmy, O.; Mukamel, R. Neural Network Underlying Intermanual SKill Transfer in Humans. Cell Rep. 2016, 17, 2891–2900. [Google Scholar] [CrossRef]
- Alahmadi, A.A.S. Investigating the sub-regions of the superor parietal cortex using functional magnetic resonance imaging connectivity. Insights Into Imaging 2021, 12, 47. [Google Scholar] [CrossRef]
- Agarwal, S.; Branson, K.; Belongie, S. Higher Order learning with graphs. In Proceedings of the 23rd International Conference on Machine Learning, Pittsburgh, PA, USA, 25–29 June 2006. [Google Scholar]
- Serrano, N.; Jaimes-Reategui, R.; Pisarchik, A. Hypergraph of Functional Connectivity Based on Event-Related Coherence: Magnetoencephalography Data Analysis. Appl. Sci. 2024, 14, 2343. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, J.; Kassani, P.; Zhang, Y.; Bai, Y.; Stephen, J.M.; Wilson, T.W.; Calhoun, V.D.; Wang, Y.P. Multi-Hypergraph Learning-Based Brain Functional Connectivity Analysis in fMRI Data. IEEE Trans. Med. Imaging 2020, 39, 1746–1758. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.; Dai, A.; Le, Q.V. HyperNetworks. arXiv 2016, arXiv:1609.09106. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, P.B.; Prabhu, S.R.; Upadhyay, Y.; Teja, N.V.; Swamy, P.A. Advanced Multi-task Reinforcement Learning Utilising Task-Adaptive Episodic Memory with Hypernetwork Integration. In Proceedings of the 2024 2nd International Conference on Intelligent Data Communication Technologies and Internet of Things (IDCIoT), Bengaluru, India, 4–6 January 2024. [Google Scholar]
- Wang, L.; Zhang, X.; Su, H. A Comprehensive Survey of Continual Learning: Theory, Method and Application. IEEE Trans. Pattern Anal. Mach. Intell. 2024, 46, 5362–5383. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Xu, C.; Liu, H.; Zhou, B.; Zhou, H. Bridging pre-trained models to continual learning: A hypernetwork based framework with parameter-efficient fine-tuning techniques. Inf. Sci. 2024, 674, 120710. [Google Scholar] [CrossRef]
- Judd, C. Generalized Experience. In Psychology of Secondary Education; Ginn & Company: Boston, MA, USA, 1927. [Google Scholar] [CrossRef]
- Zhuang, F.; Qi, Z.; Duan, K.; Xi, D.; Zhu, Y.; Zhu, H.; Xiong, H.; He, Q. A comprehensive survey on transfer learning. Proc. IEEE 2020, 109, 43–76. [Google Scholar] [CrossRef]
- McKissock, S.; Ward, J. Do errors matter? Errorless and errorful learning in anomic picture naming. Neuropsychol. Rehabil. 2007, 17, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Shimokawa, T.; Peper, F.; Tanemura, R. Functional network activity during errorless and trial-and-error color-name association learning. Brain Behav. 2020, 10, e01723. [Google Scholar] [CrossRef] [PubMed]
- Zárate-Rochín, A.M. Contemporary neurocognitive models of memory: A descriptive comparative analysis. Neuropsychologia 2024, 108846. [Google Scholar] [CrossRef]
- Pyc, M.A.; Rawson, K. Testing the retrieval effort hypothesis: Does greater difficulty correctly recalling information lead to higher levels of memory? J. Mem. Lang. 2009, 60, 437–447. [Google Scholar] [CrossRef]
- Bjork, R. Memory and metamemory considerations in the training of human beings. In Metacognition: Knowing About Knowing; MIT Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Van Buuren, M.; Wagner, I.; Fernandez, G. Functional network interactions at rest underlie individual differences in memory ability. Learn. Mem. 2019, 26, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gerraty, R.T.; Davidow, J.Y.; Wimmer, G.E.; Kahn, I.; Shohamy, D. Transfer of Learning Relates to Intrinsic Connectivity between Hippocampus, Ventromedial Prefrontal Cortex, and Large-Scale Networks. J. Neurosci. 2014, 34, 11297–11303. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R. The backpropagation Algorithm. In Neural Networks: A Systematic Introduction; Springer: Cham, Swtizerland, 1996. [Google Scholar] [CrossRef]
- Sacramento, J.; Costa, R.P.; Bengio, Y.; Senn, W. Dendritic cortical microcircuits approximate the backpropagation algorithm. Adv. Neural Inf. Process. Syst. 2018, 31. [Google Scholar]
- Song, Y.; Lukasiewicz, T.; Xu, Z.; Bogacz, R. Can the brain do backpropagation? Exact implementation of backpropagation in predictive coding networks. Adv. Neural Inf. Process. Syst. 2020, 33, 22566–22579. [Google Scholar]
- Song, Y.; Millidge, B.; Salvatori, T.; Lukasiewicz, T.; Xu, Z.; Bogacz, R. Inferring neural activity before plasticity as a foundation for learning beyond backpropagation. Nat. Neurosci. 2023, 27, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.; Kebria, P.; Khosravi, A.; Nahavandi, S. A Survey of Imitation Learning: Algorithms, Recent Developments, and Challenges. IEEE Trans. Cybern. 2023. [Google Scholar] [CrossRef] [PubMed]
- Leiva, F.; Ruiz-del Solar, J. Combining RL and IL using a dynamic, performance-based modulation over learning signals and its application to local planning. arXiv, 2024; arXiv:2405.09760. [Google Scholar] [CrossRef]
- Sestieri, C.; Shulman, G.; Corbetta, M. Orienting to the environment: Separate contributions of dorsal and ventral frontoparietal attention networks. In The Neuroscience of Attention: Attentional Control and Selection; Oxford Academic: Oxford, UK, 2012. [Google Scholar] [CrossRef]
- Seeley, W.; Menon, V.; Scchatzberg, A.; Keller, J.; Glover, G.; Kenna, H. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Schimmelpfennig, J.; Topczewski, J.; Zajkowski, W.; Jankowiak-Siuda, K. The role of the salience network in cognitive and affective defecits. Front. Hum. Neurosci. 2023, 17, 1133367. [Google Scholar] [CrossRef] [PubMed]
- Grecius, M.; Krasnow, B.; Reiss, A.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Koch, I.; Poljac, E.; Muller, H.; Kiesel, A. Cognitive structure, flexibility, and plasticity in human multitasking—An integrative review of dual-task and task-switching research. Psychol. Bull. 2018, 144, 557. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.; Vartanian, O.; Hollands, J. The brain under cognitive workload: Neural networks underlying multitasking performance in the multi-attribute task battery. Neuropsychologia 2022, 174, 108350. [Google Scholar] [CrossRef] [PubMed]
- Garrison, K.; Scheinost, D.; Worhunsky, P.; Elwafi, H.; Thornhill, T.; Thompson, E.; Saron, C.; Desbordes, G.; Kober, H.; Hampson, M.; et al. Real-time fMRI links ssubjective experience with brain activity during focused attention. Neuroimage 2013, 81, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, A.; Tang, W.; Heilbronner, S. Connecting Circuits with Networks in Addiction Neuroscience: A Salience Network Perspective. Int. J. Mol. Sci. 2023, 24, 9083. [Google Scholar] [CrossRef] [PubMed]
- Crawshaw, M. Multi-task learning with deep neural networks: A survey. arXiv 2020, arXiv:2009.09796. [Google Scholar]
- Agliari, E.; Alessandrelli, A.; Barra, A.; Ricci-Tersenghi, F. Parallel learning by multitasking neural networks. J. Stat. Mech. 2023, 2023, 113401. [Google Scholar] [CrossRef]
- Parisi, G.; Kemker, R.; Part, J.; Kanan, C.; Wermter, S. Continual Learning with neural networks: A review. Neural Netw. 2018, 113, 54–71. [Google Scholar] [CrossRef]
- Wu, H.; Khetarpal, K.; Precup, D. Self-supervised Attention-Awware Reinforcement Learning. In Proceedings of the AAAI conference on Artificial Intelligence, Virtual, 2–9 February 2021. [Google Scholar]
- Vaswani, A.; Shazeer, N.; Parmar, N.; Uszkoreit, J.; Jones, L.; Gomez, A.; Kaiser, L.; Polosukhin, i. Attention is all you need. In Proceedings of the 31st Conference on Neural Information Processing Systems (NIPS 2017), Long Beach, CA, USA, 4–9 December 2023. [Google Scholar]
- Zhu, X.; Zhao, L.; Zhu, W. Salience Interest Option: Temporal abstraction with salience interest functions. Neural Netw. 2024, 176, 106342. [Google Scholar] [CrossRef] [PubMed]
- Pina, R.; De Silva, V.; Artaud, C. Towards Self-Adaptive Resilient Swarms Using Multi-Agent Reinforcement Learning. In Proceedings of the 13th International Conference on Pattern Recognition Applications and Methods (ICPRAM 2024), Rome, Italy, 24–26 February 2024. [Google Scholar]
- Bissell, D.; Birtchnell, T.; Elliott, A.; Hsu, E.L. Autonomous automobilities: The social impacts of driverless vehicles. Curr. Sociol. 2020, 68, 116–134. [Google Scholar] [CrossRef]
- Khan, M.A.; Sayed, H.E.; Malik, S.; Zia, T.; Khan, J.; Alkaabi, N.; Ignatious, H. Level-5 autonomous driving—Are we there yet? a review of research literature. ACM Comput. Surv. CSUR 2022, 55, 1–38. [Google Scholar] [CrossRef]
- Wang, J.; Huang, H.; Li, K.; Li, J. Towards the unified principles for level 5 autonomous vehicles. Engineering 2021, 7, 1313–1325. [Google Scholar] [CrossRef]
- Wong, K.; Gu, Y.; Kamijo, S. Mapping for autonomous driving: Opportunities and challenges. IEEE Intell. Transp. Syst. Mag. 2020, 13, 91–106. [Google Scholar] [CrossRef]
- Barabas, I.; Todoruţ, A.; Cordoş, N.; Molea, A. Current challenges in autonomous driving. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Barcelona, Spain, 14–16 July 2017; Volume 252, p. 012096. [Google Scholar]
- Lin, J.; Zhou, W.; Wang, H.; Cao, Z.; Yu, W.; Zhao, C.; Zhao, D.; Yang, D.; Li, J. Road traffic law adaptive decision-making for self-driving vehicles. In Proceedings of the 2022 IEEE 25th International Conference on Intelligent Transportation Systems (ITSC), Macau, China, 8–12 October 2022; pp. 2034–2041. [Google Scholar]
- Erkent, Ö.; Laugier, C. Semantic segmentation with unsupervised domain adaptation under varying weather conditions for autonomous vehicles. IEEE Robot. Autom. Lett. 2020, 5, 3580–3587. [Google Scholar] [CrossRef]
- Galántai, P. Assessment of dangerous traffic situations for autonomous vehicles. Period. Polytech. Transp. Eng. 2022, 50, 260–266. [Google Scholar] [CrossRef]
- Huang, Y.; Eden, J.; Ivanova, E.; Burdet, E. Can Training Make Three Arms Better Than Two Heads for Trimanual Coordination? IEEE Open J. Eng. Med. Biol. 2023, 4, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Amirthanayagam, A.; Zecca, M.; Barber, S.; Singh, B.; Moss, E. Impact of minimally invasive surgery on surgeon health (ISSUE) study: Protocol of a single-arm observational study conducted in the live surgery setting. Br. Med. J. 2023, 13, e066765. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Moreno, Y.; Echevarria, S.; Vidal-Valderrama, C.; Pianetti, L.; Cordova-Guilarte, J.; Navarro-Gonzalez, J.; Acevedo-Rodríguez, J.; Dorado-Avila, G.; Osorio-Romero, L.; Chavez-Campos, C.; et al. Robotic surgery: A comprehensive review of the literature and current trends. Cureus 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kosmyna, N.; Hauptmann, E.; Hmaidan, Y. A Brain-Controlled Quadruped Robot: A Proof-of-Concept Demonstration. Sensors 2023, 24, 80. [Google Scholar] [CrossRef] [PubMed]
- Shajari, S.; Kuruvinashetti, K.; Komeili, A.; Sundararaj, U. The emergence of AI-based wearable sensors for digital health technology: A review. Sensors 2023, 23, 9498. [Google Scholar] [CrossRef] [PubMed]
- Rault, T.; Bouabdallah, A.; Challal, Y.; Marin, F. A survey of energy-efficient context recognition systems using wearable sensors for healthcare applications. Pervasive Mob. Comput. 2017, 37, 23–44. [Google Scholar] [CrossRef]
- Tawakuli, A.; Kaiser, D.; Engel, T. Synchronized preprocessing of sensor data. In Proceedings of the 2020 IEEE International Conference on Big Data (Big Data), Atlanta, GA, USA, 10–13 December 2020; pp. 3522–3531. [Google Scholar]
- Tubis, A.A.; Rohman, J. Intelligent Warehouse in Industry 4.0—Systematic Literature Review. Sensors 2023, 23, 4105. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, F.; Zou, Z.; Duffy, V. Application, Development and Future Opportunities of Collaborative Robots (Cobots) in Manufacturing: A Literature Review. Int. J. Hum. Comput. Interact. 2024, 40, 915–932. [Google Scholar] [CrossRef]
- Inam, R.; Fersman, E.; Raizer, K.; Souza, R.; Nascimento, A.; Hata, A. Safety for Automated Warehouse exhibiting collaborative robots. In Safety and Reliability—Safe Societies in a Changing World; CRC Press: Boca Raton, FL, USA, 2018; pp. 2021–2028. [Google Scholar]
- Shi, D.; Tong, J.; Liu, Y.; Fan, W. Knowledge Reuse of Multi-Agent Reinforcement Learning in Cooperative Tasks. Entropy 2022, 24, 470. [Google Scholar] [CrossRef]
- Gao, Z.; Xu, K.; Ding, B.; Wang, H. Knowru: Knowledge reuse via knowledge distillation in multi-agent reinforcement learning. Entropy 2021, 23, 1043. [Google Scholar] [CrossRef]
- Musk, E.; Neuralink. An Integrated Brain–Machine Interface Platform with Thousands of Channels. J. Med. Internet Res. 2019, 21, e16194. [Google Scholar] [CrossRef]
- Karikari, E.; Koshechkin, K. Review on brain–computer interface technologies in healthcare. Biophys. Rev. 2023, 15, 1351–1358. [Google Scholar] [CrossRef]
- Parui, S.; Samanta, D.; Chakravorty, N. An Advanced Healthcare System Where Internet of Things meets Brain-Computer Interface Using Event-Related Potential. In Proceedings of the 24th International Conference on Distributed Computing and Networking, Kharagpur, India, 4–7 January 2023; pp. 438–443. [Google Scholar] [CrossRef]
- Zhao, Z.P.; Nie, C.; Jiang, C.T.; Cao, S.H.; Tian, K.X.; Yu, S.; Gu, J.W. Modulating Brain Activity with Invasive Brain-Computer Interface: A Narrative Review. Brain Sci. 2023, 13, 134. [Google Scholar] [CrossRef]
- Prattichizzo, D.; Pozzi, M.; Baldi, T.L.; Malvezzi, M.; Hussain, I.; Rossi, S.; Salvietti, G. Human augmentation by wearable supernumerary robotic limbs: Review and perspectives. Prog. Biomed. Eng. 2021, 3, 042005. [Google Scholar] [CrossRef]
- Penaloza, C.; Hernandez-Carmona, D.; Nishio, S. Towards intelligent brain-controlled body augmentation robotic limbs. In Proceedings of the 2018 IEEE International Conference on Systems, Man, and Cybernetics (SMC), Miyazaki, Japan, 7–10 October 2018; pp. 1011–1015. [Google Scholar]
- Zhou, Y.; Yu, T.; Gao, W.; Huang, W.; Lu, Z.; Huang, Q.; Li, Y. Shared three-dimensional robotic arm control based on asynchronous BCI and computer vision. IEEE Trans. Neural Syst. Rehabil. Eng. 2023. [Google Scholar] [CrossRef]
- Drew, L. Neuralink brain chip: Advance sparks safety and secrecy concerns. Nature 2024, 627, 19. [Google Scholar] [CrossRef] [PubMed]
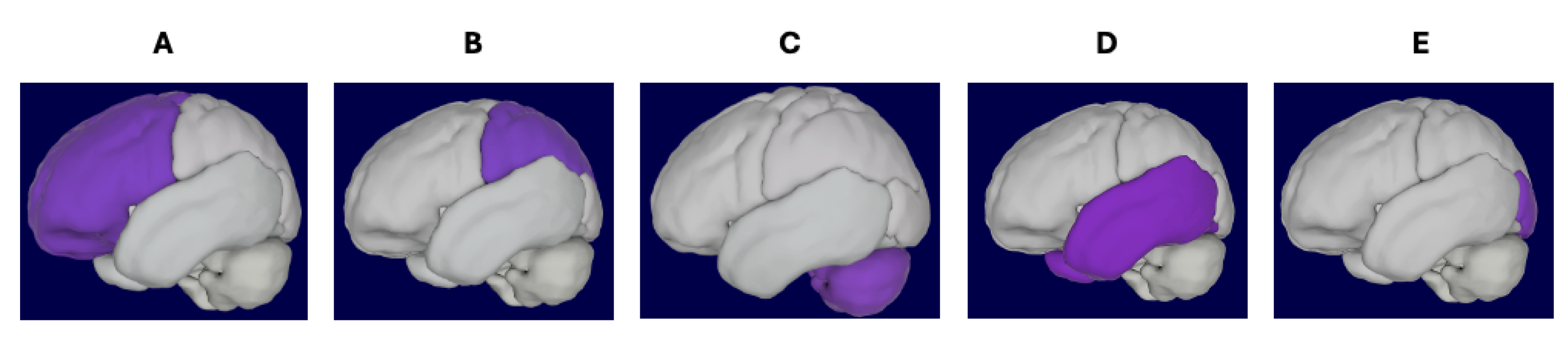

| Brain Region | Attributes |
|---|---|
| Frontal Lobe | Plays a crucial role in higher cognitive functions such as reasoning, planning, decision-making, and voluntary movement. |
| Temporal Lobe | Involved in auditory processing, memory, language comprehension, and emotional responses. |
| Brain Stem | Regulation of autonomous bodily functions such as breathing and digestion. |
| Parietal Lobe | Processing sensory information, spatial awareness, perception, and integrating sensory input with motor function for coordinated movement and orientation in space. |
| Occipital Lobe | Visual information processing and integrating it with higher-level visual functions such as object recognition, color discrimination, and depth perception. |
| Cerebellum | Coordinating voluntary movements such as balance and motor learning. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Illeperuma, M.; Pina, R.; De Silva, V.; Liu, X. Novel Directions for Neuromorphic Machine Intelligence Guided by Functional Connectivity: A Review. Machines 2024, 12, 574. https://doi.org/10.3390/machines12080574
Illeperuma M, Pina R, De Silva V, Liu X. Novel Directions for Neuromorphic Machine Intelligence Guided by Functional Connectivity: A Review. Machines. 2024; 12(8):574. https://doi.org/10.3390/machines12080574
Chicago/Turabian StyleIlleperuma, Mindula, Rafael Pina, Varuna De Silva, and Xiaolan Liu. 2024. "Novel Directions for Neuromorphic Machine Intelligence Guided by Functional Connectivity: A Review" Machines 12, no. 8: 574. https://doi.org/10.3390/machines12080574