Early Prediction of Tumor Response to Neoadjuvant Chemotherapy and Clinical Outcome in Breast Cancer Using a Novel FDG-PET Parameter for Cancer Stem Cell Metabolism


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Study Design
2.2. Histopathologic Analysis
2.3. 18F-FDG PET/CT Procedures
2.4. Novel PET Parameters for CSC Metabolism
2.5. Statistical Analysis
3. Results
3.1. Patient Characteristics
3.2. Association between the Achievement of pCR and Clinicopathologic/Metabolic Parameters in HER2-positive and TN Subtypes
3.3. Prediction of the Pathologic Response with MTVcsc
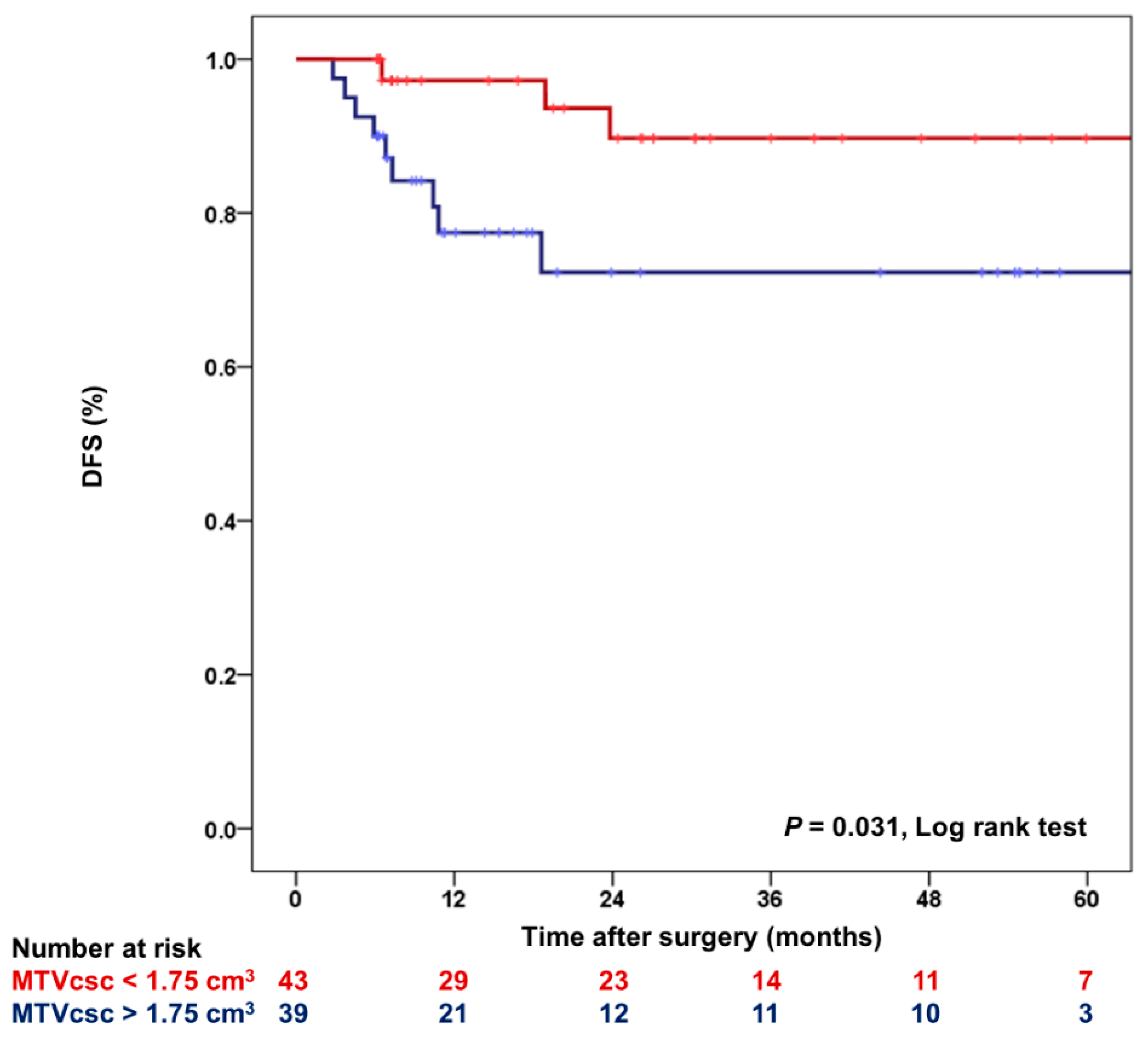
3.4. The Relation between MTVcsc and DFS in HER2-Positive and TN Subtypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shackleton, M.; Quintana, E.; Fearon, E.R.; Morrison, S.J. Heterogeneity in cancer: Cancer stem cells versus clonal evolution. Cell 2009, 138, 822–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, A.; Chang, H.; Huang, S. Non-genetic heterogeneity—A mutation-independent driving force for the somatic evolution of tumours. Nat. Rev. Genet. 2009, 10, 336–342. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Nassar, D.; Blanpain, C. Cancer stem cells: Basic concepts and therapeutic implications. Annu. Rev. Pathol. 2016, 11, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Prager, B.C.; Xie, Q.; Bao, S.; Rich, J.N. Cancer stem cells: The architects of the tumor ecosystem. Cell Stem Cell 2019, 24, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatt, M.; Le Rest, C.C.; Tixier, F.; Badic, B.; Schick, U.; Visvikis, D. Radiomics: Data are also images. J. Nucl. Med. 2019, 60, 38S–44S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C.; Bodet-Milin, C.; Bourgeois, M.; Gouard, S.; Ansquer, C.; Barbaud, M.; Sebille, J.C.; Cherel, M.; Kraeber-Bodere, F.; Carlier, T. Exploring tumor heterogeneity using PET imaging: The big picture. Cancers 2019, 11, 1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traverso, A.; Wee, L.; Dekker, A.; Gillies, R. Repeatability and reproducibility of radiomic features: A systematic review. Int. J. Radiat. Oncol. Biol. Phys. 2018, 102, 1143–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menendez, J.A.; Joven, J.; Cufi, S.; Corominas-Faja, B.; Oliveras-Ferraros, C.; Cuyas, E.; Martin-Castillo, B.; Lopez-Bonet, E.; Alarcon, T.; Vazquez-Martin, A. The Warburg effect version 2.0: Metabolic reprogramming of cancer stem cells. Cell Cycle 2013, 12, 1166–1179. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Suda, T. Metabolic requirements for the maintenance of self-renewing stem cells. Nat. Rev. Mol. Cell Biol. 2014, 15, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, J.; Lima, J. Metabolic traits of cancer stem cells. Dis. Models Mech. 2018, 11, dmm033464. [Google Scholar] [CrossRef] [Green Version]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Pece, S.; Tosoni, D.; Confalonieri, S.; Mazzarol, G.; Vecchi, M.; Ronzoni, S.; Bernard, L.; Viale, G.; Pelicci, P.G.; Di Fiore, P.P. Biological and molecular heterogeneity of breast cancers correlates with their cancer stem cell content. Cell 2010, 140, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riester, M.; Xu, Q.; Moreira, A.; Zheng, J.; Michor, F.; Downey, R.J. The Warburg effect: Persistence of stem-cell metabolism in cancers as a failure of differentiation. Ann. Oncol. 2018, 29, 264–270. [Google Scholar] [CrossRef]
- Fujii, T.; Kogawa, T.; Dong, W.; Sahin, A.A.; Moulder, S.; Litton, J.K.; Tripathy, D.; Iwamoto, T.; Hunt, K.K.; Pusztai, L.; et al. Revisiting the definition of estrogen receptor positivity in HER2-negative primary breast cancer. Ann. Oncol. 2017, 28, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Nioche, C.; Orlhac, F.; Boughdad, S.; Reuze, S.; Goya-Outi, J.; Robert, C.; Pellot-Barakat, C.; Soussan, M.; Frouin, F.; Buvat, I. LIFEx: A freeware for radiomic feature calculation in multimodality imaging to accelerate advances in the characterization of tumor heterogeneity. Cancer Res. 2018, 78, 4786–4789. [Google Scholar] [CrossRef] [Green Version]
- Seo, A.N.; Lee, H.J.; Kim, E.J.; Jang, M.H.; Kim, Y.J.; Kim, J.H.; Kim, S.W.; Ryu, H.S.; Park, I.A.; Im, S.A.; et al. Expression of breast cancer stem cell markers as predictors of prognosis and response to trastuzumab in HER2-positive breast cancer. Br. J. Cancer 2016, 114, 1109–1116. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.R.; Dawson, S.-J.; Blows, F.M.; Provenzano, E.; Pharoah, P.D.; Caldas, C. Cancer stem cell markers in breast cancer: Pathological, clinical and prognostic significance. Breast Cancer Res. 2011, 13, R118. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Nenutil, R.; Appleyard, M.V.; Murray, K.; Boylan, M.; Thompson, A.M.; Coates, P.J. Lack of correlation of stem cell markers in breast cancer stem cells. Br. J. Cancer 2014, 110, 2063–2071. [Google Scholar] [CrossRef] [Green Version]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Intlekofer, A.M.; Finley, L.W.S. Metabolic signatures of cancer cells and stem cells. Nat. Metab. 2019, 1, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.G. Understanding cancer stem cell heterogeneity and plasticity. Cell Res. 2012, 22, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Samanta, D.; Gilkes, D.M.; Chaturvedi, P.; Xiang, L.; Semenza, G.L. Hypoxia-inducible factors are required for chemotherapy resistance of breast cancer stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, E5429–E5438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Liu, F.; Han, L.; Zhao, L.; Chen, J.; Olopade, O.I.; He, M.; Wei, M. HIF-2alpha promotes conversion to a stem cell phenotype and induces chemoresistance in breast cancer cells by activating Wnt and Notch pathways. J. Exp. Clin. Cancer Res. 2018, 37, 256. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factors: Coupling glucose metabolism and redox regulation with induction of the breast cancer stem cell phenotype. EMBO J. 2017, 36, 252–259. [Google Scholar] [CrossRef]
- Peng, F.; Wang, J.H.; Fan, W.J.; Meng, Y.T.; Li, M.M.; Li, T.T.; Cui, B.; Wang, H.F.; Zhao, Y.; An, F.; et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene 2018, 37, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Keith, B.; Simon, M.C. Hypoxia-inducible factors, stem cells, and cancer. Cell 2007, 129, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.P.; Liao, J.; Tang, Z.J.; Wu, W.J.; Yang, J.; Zeng, Z.L.; Hu, Y.; Wang, P.; Ju, H.Q.; Xu, R.H.; et al. Metabolic regulation of cancer cell side population by glucose through activation of the Akt pathway. Cell Death Differ. 2014, 21, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Ciavardelli, D.; Rossi, C.; Barcaroli, D.; Volpe, S.; Consalvo, A.; Zucchelli, M.; De Cola, A.; Scavo, E.; Carollo, R.; D’Agostino, D.; et al. Breast cancer stem cells rely on fermentative glycolysis and are sensitive to 2-deoxyglucose treatment. Cell Death Dis. 2014, 5, e1336. [Google Scholar] [CrossRef] [Green Version]
- Vlashi, E.; Lagadec, C.; Vergnes, L.; Reue, K.; Frohnen, P.; Chan, M.; Alhiyari, Y.; Dratver, M.B.; Pajonk, F. Metabolic differences in breast cancer stem cells and differentiated progeny. Breast Cancer Res. Treat. 2014, 146, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, J.; Schwab, J.; Schwenck, J.; Chen, Q.; Quintanilla-Martinez, L.; Hahn, M.; Wietek, B.; Schwenzer, N.; Staebler, A.; Kohlhofer, U.; et al. Decoding intratumoral heterogeneity of breast cancer by multiparametric in vivo imaging: A translational study. Cancer Res. 2016, 76, 5512–5522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortazar, P.; Zhang, L.; Untch, M.; Mehta, K.; Costantino, J.P.; Wolmark, N.; Bonnefoi, H.; Cameron, D.; Gianni, L.; Valagussa, P.; et al. Pathological complete response and long-term clinical benefit in breast cancer: The CTNeoBC pooled analysis. Lancet 2014, 384, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Asselain, B.; Barlow, W.; Bartlett, J.; Bergh, J.; Bergsten-Nordström, E.; Bliss, J.; Bradley, R. Long-term outcomes for neoadjuvant versus adjuvant chemotherapy in early breast cancer: Meta-analysis of individual patient data from ten randomised trials. Lancet Oncol. 2018, 19, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Humbert, O.; Riedinger, J.M.; Charon-Barra, C.; Berriolo-Riedinger, A.; Desmoulins, I.; Lorgis, V.; Kanoun, S.; Coutant, C.; Fumoleau, P.; Cochet, A.; et al. Identification of biomarkers including 18FDG-PET/CT for early prediction of response to neoadjuvant chemotherapy in triple-negative breast cancer. Clin. Cancer Res. 2015, 21, 5460–5468. [Google Scholar] [CrossRef] [Green Version]
- Groheux, D.; Hindie, E.; Giacchetti, S.; Hamy, A.S.; Berger, F.; Merlet, P.; de Roquancourt, A.; de Cremoux, P.; Marty, M.; Hatt, M.; et al. Early assessment with 18F-fluorodeoxyglucose positron emission tomography/computed tomography can help predict the outcome of neoadjuvant chemotherapy in triple negative breast cancer. Eur. J. Cancer 2014, 50, 1864–1871. [Google Scholar] [CrossRef]
- Humbert, O.; Berriolo-Riedinger, A.; Riedinger, J.M.; Coudert, B.; Arnould, L.; Cochet, A.; Loustalot, C.; Fumoleau, P.; Brunotte, F. Changes in 18F-FDG tumor metabolism after a first course of neoadjuvant chemotherapy in breast cancer: Influence of tumor subtypes. Ann. Oncol. 2012, 23, 2572–2577. [Google Scholar] [CrossRef]
- Tan, W.; Yang, M.; Yang, H.; Zhou, F.; Shen, W. Predicting the response to neoadjuvant therapy for early-stage breast cancer: Tumor-, blood-, and imaging-related biomarkers. Cancer Manag. Res. 2018, 10, 4333–4347. [Google Scholar] [CrossRef] [Green Version]
- Coudert, B.; Pierga, J.Y.; Mouret-Reynier, M.A.; Kerrou, K.; Ferrero, J.M.; Petit, T.; Kerbrat, P.; Dupre, P.F.; Bachelot, T.; Gabelle, P.; et al. Use of [(18)F]-FDG PET to predict response to neoadjuvant trastuzumab and docetaxel in patients with HER2-positive breast cancer, and addition of bevacizumab to neoadjuvant trastuzumab and docetaxel in [(18)F]-FDG PET-predicted non-responders (AVATAXHER): An open-label, randomised phase 2 trial. Lancet Oncol. 2014, 15, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Heil, J.; Kuerer, H.M.; Pfob, A.; Rauch, G.; Sinn, H.P.; Golatta, M.; Liefers, G.J.; Vrancken Peeters, M.J. Eliminating the breast cancer surgery paradigm after neoadjuvant systemic therapy: Current evidence and future challenges. Ann. Oncol. 2020, 31, 61–71. [Google Scholar] [CrossRef] [Green Version]
- D’Alterio, C.; Scala, S.; Sozzi, G.; Roz, L.; Bertolini, G. Paradoxical effects of chemotherapy on tumor relapse and metastasis promotion. Semin. Cancer Biol. 2020, 60, 351–361. [Google Scholar] [CrossRef]
- Creighton, C.J.; Li, X.; Landis, M.; Dixon, J.M.; Neumeister, V.M.; Sjolund, A.; Rimm, D.L.; Wong, H.; Rodriguez, A.; Herschkowitz, J.I.; et al. Residual breast cancers after conventional therapy display mesenchymal as well as tumor-initiating features. Proc. Natl. Acad. Sci. USA 2009, 106, 13820–13825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klintman, M.; Buus, R.; Cheang, M.C.; Sheri, A.; Smith, I.E.; Dowsett, M. Changes in expression of genes representing key biologic processes after neoadjuvant chemotherapy in breast cancer, and prognostic implications in residual disease. Clin. Cancer Res. 2016, 22, 2405–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Giuliano, M.; Trivedi, M.V.; Schiff, R.; Osborne, C.K. Metastasis dormancy in estrogen receptor-positive breast cancer. Clin. Cancer Res. 2013, 19, 6389–6397. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.D.; Burness, M.L.; Wicha, M.S. Therapeutic implications of cellular heterogeneity and plasticity in breast cancer. Cell Stem Cell 2015, 17, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Shang, L.; Brooks, M.D.; Jiagge, E.; Zhu, Y.; Buschhaus, J.M.; Conley, S.; Fath, M.A.; Davis, A.; Gheordunescu, E.; et al. Targeting breast cancer stem cell state equilibrium through modulation of redox signaling. Cell Metab. 2018, 28, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.; Lu, M.; Jung, K.H.; Park, J.H.; Yu, L.; Onuchic, J.N.; Kaipparettu, B.A.; Levine, H. Elucidating cancer metabolic plasticity by coupling gene regulation with metabolic pathways. Proc. Natl. Acad. Sci. USA 2019, 116, 3909–3918. [Google Scholar] [CrossRef] [Green Version]
- Gawrzak, S.; Rinaldi, L.; Gregorio, S.; Arenas, E.J.; Salvador, F.; Urosevic, J.; Figueras-Puig, C.; Rojo, F.; Del Barco Barrantes, I.; Cejalvo, J.M.; et al. MSK1 regulates luminal cell differentiation and metastatic dormancy in ER(+) breast cancer. Nat. Cell Biol. 2018, 20, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Fluegen, G.; Avivar-Valderas, A.; Wang, Y.; Padgen, M.R.; Williams, J.K.; Nobre, A.R.; Calvo, V.; Cheung, J.F.; Bravo-Cordero, J.J.; Entenberg, D.; et al. Phenotypic heterogeneity of disseminated tumour cells is preset by primary tumour hypoxic microenvironments. Nat. Cell Biol. 2017, 19, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Kreso, A.; O’Brien, C.A.; van Galen, P.; Gan, O.I.; Notta, F.; Brown, A.M.; Ng, K.; Ma, J.; Wienholds, E.; Dunant, C.; et al. Variable clonal repopulation dynamics influence chemotherapy response in colorectal cancer. Science 2013, 339, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.S. Molecular pathways: Targeting cancer stem cells awakened by chemotherapy to abrogate tumor repopulation. Clin. Cancer Res. 2016, 22, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lewis, M.T.; Huang, J.; Gutierrez, C.; Osborne, C.K.; Wu, M.F.; Hilsenbeck, S.G.; Pavlick, A.; Zhang, X.; Chamness, G.C.; et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J. Natl. Cancer Inst. 2008, 100, 672–679. [Google Scholar] [CrossRef]
- Malta, T.M.; Sokolov, A.; Gentles, A.J.; Burzykowski, T.; Poisson, L.; Weinstein, J.N.; Kaminska, B.; Huelsken, J.; Omberg, L.; Gevaert, O.; et al. Machine learning identifies stemness features associated with oncogenic dedifferentiation. Cell 2018, 173, 338–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
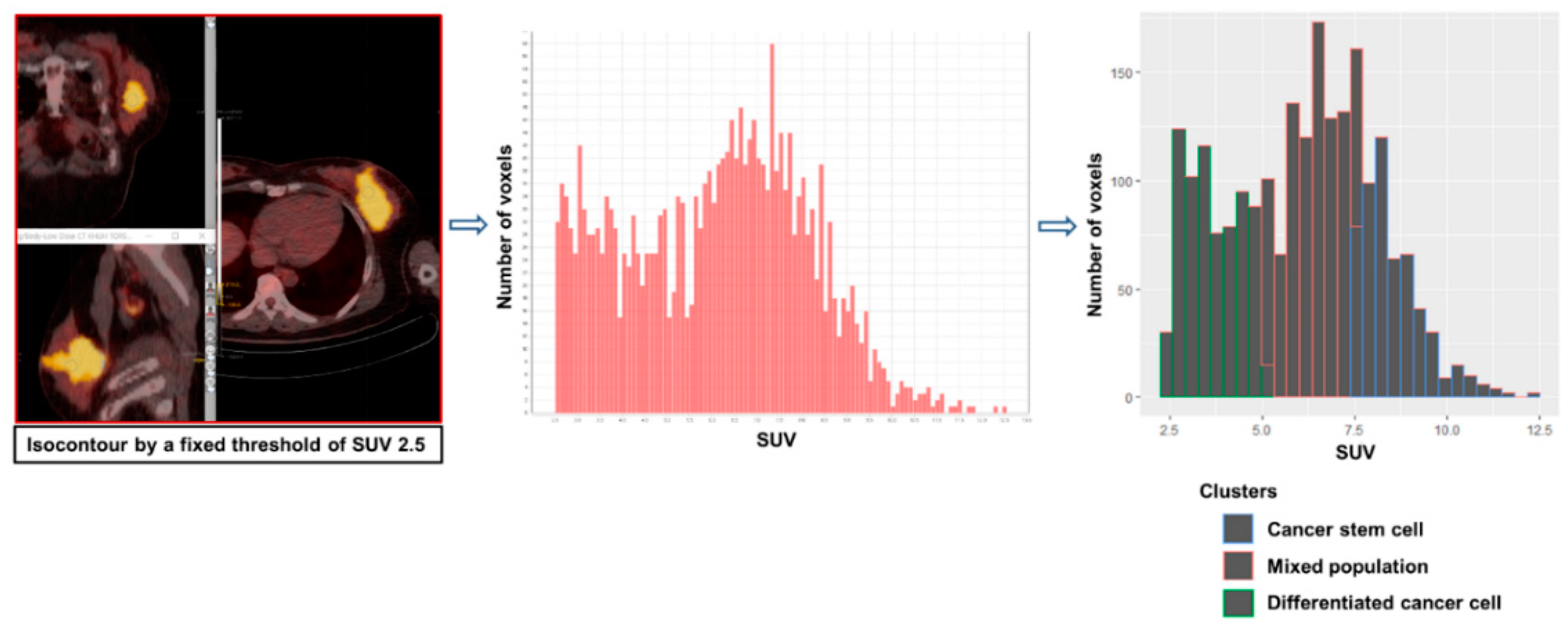




{kind=link}
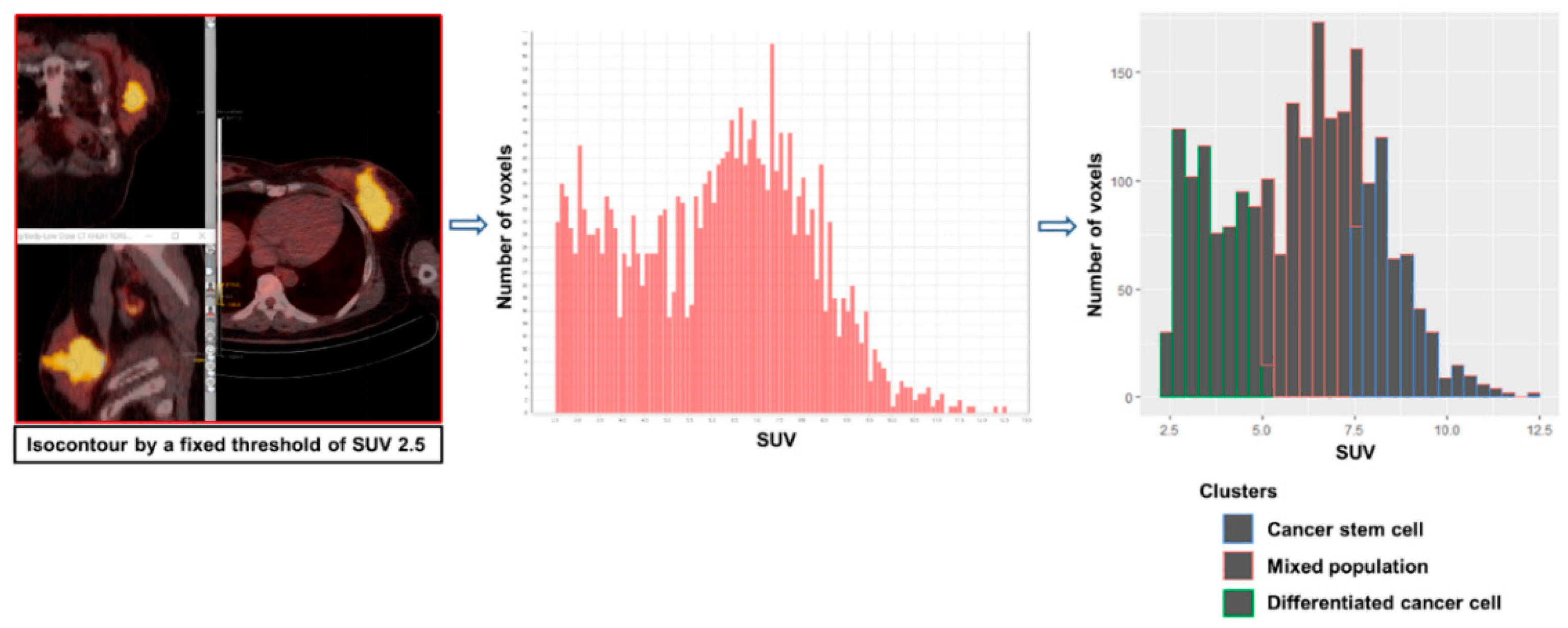
{kind=link}
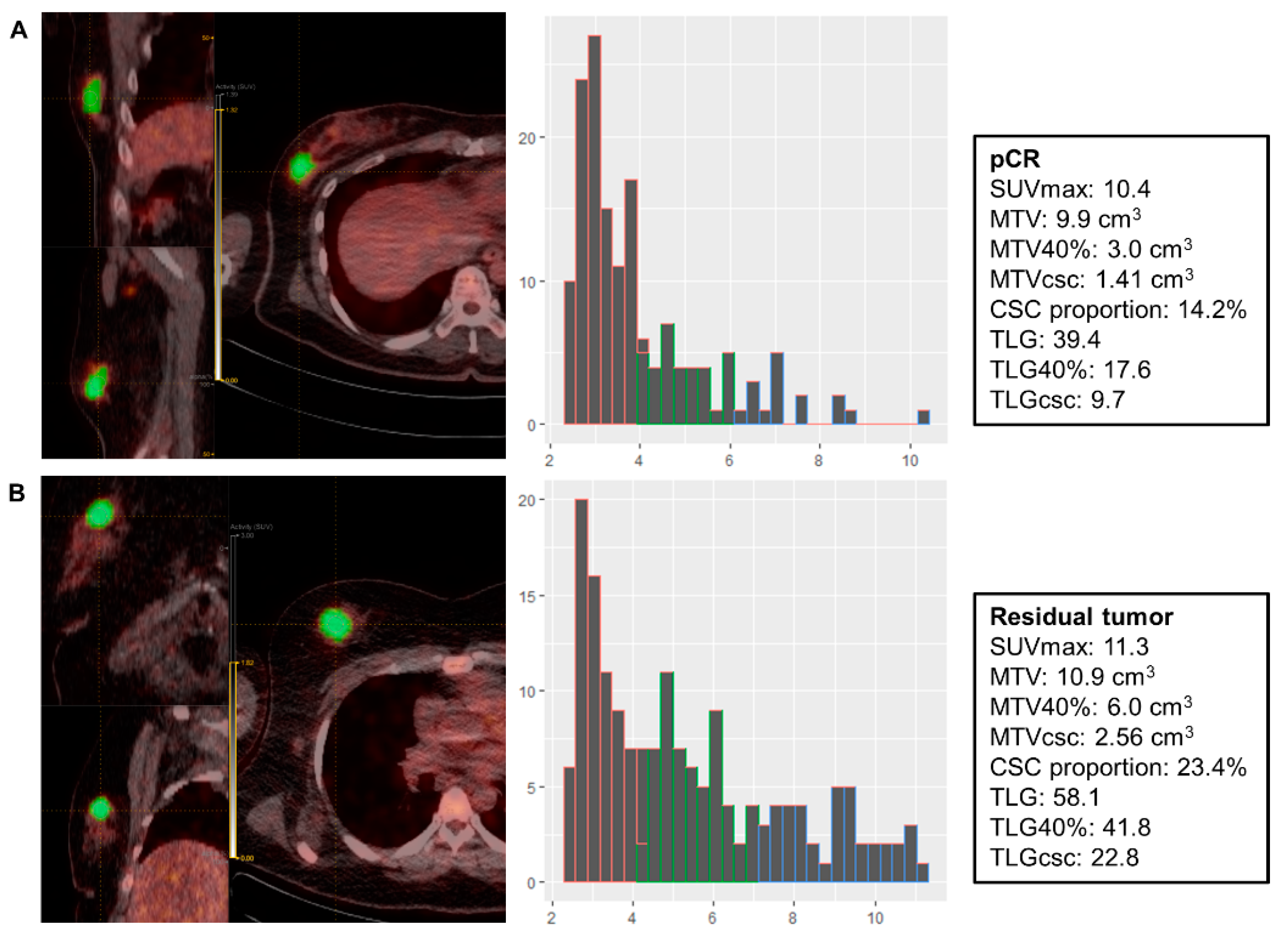
{kind=link}
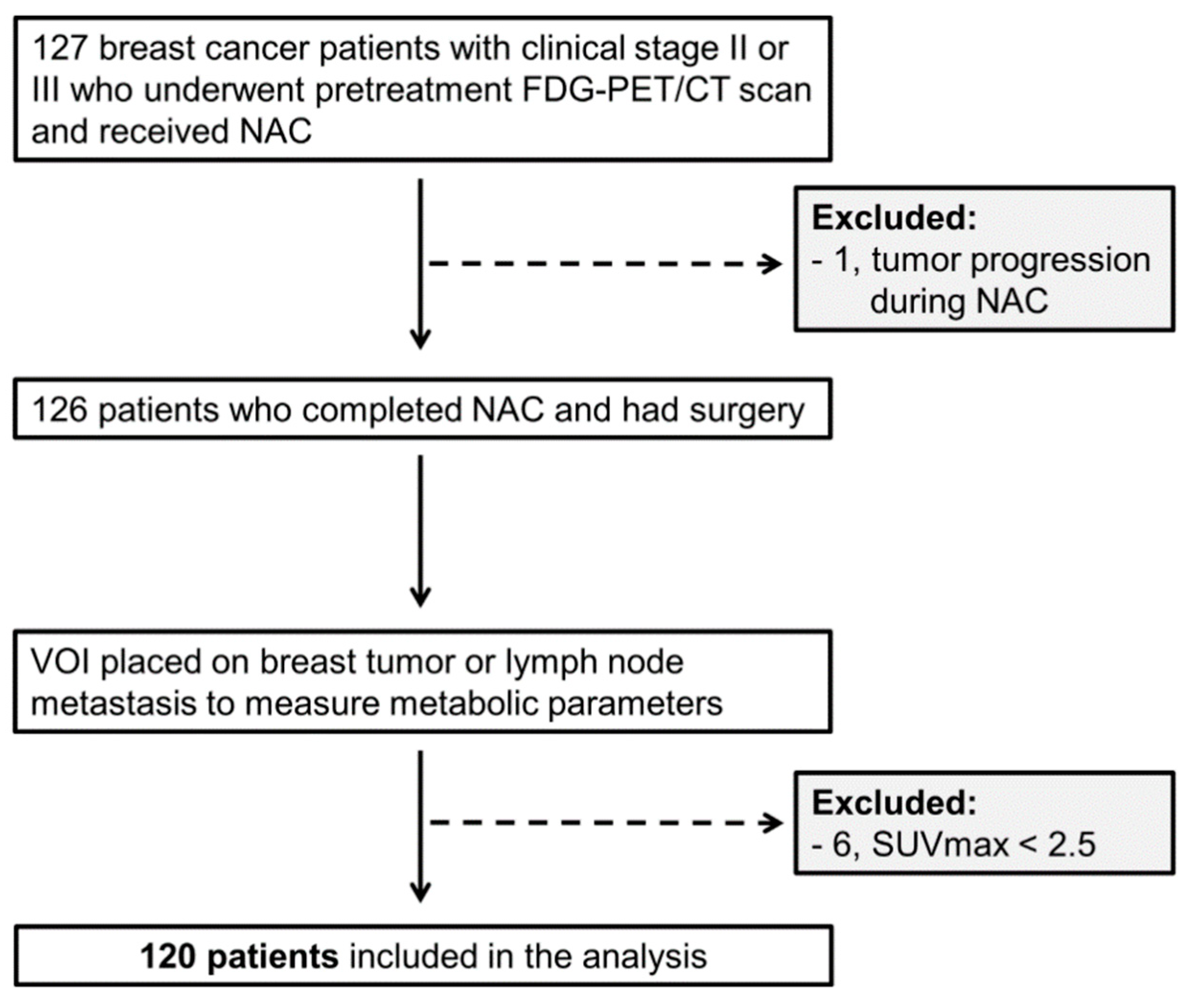
{kind=link}
| Characteristics | N = 120 |
|---|---|
| Age (years), median (range) | 49 (25–72) |
| Histology | |
| IDC | 115 (96%) |
| ILC/other | 5 (4%) |
| Clinical T stage | |
| T1-2 | 79 (66%) |
| T3-4 | 41 (34%) |
| Clinical anatomic stage | |
| IIA-IIIA | 97 (81%) |
| IIIB-IIIC | 23 (19%) |
| ER status | |
| Positive | 65 (54%) |
| Negative | 55 (46%) |
| Molecular subtype | |
| Luminal (HER2-negative) | 38 (32%) |
| HER2-positive | 57 (47%) |
| Triple negative | 25 (21%) |
| NAC regimen | |
| Anthracycline and taxane based | 92 (77%) |
| Anthracycline based | 10 (8%) |
| Taxane based | 18 (15%) |
| Anti-HER2 therapy in 57 HER2-positive patients | |
| Neoadjuvant | 31 (55%) |
| Adjuvant only | 19 (33%) |
| None | 7 (12%) |
| NAC response | |
| pCR (ypT0/is ypN0) | 22 (18%) |
| Residual tumor | 98 (82%) |
| Surgery | |
| Breast-conserving surgery | 66 (55%) |
| Mastectomy | 54 (45%) |
| Recurrence | |
| Yes | 16 (13%) |
| No | 104 (87%) |
| Parameters | HER2-Positive/TN | Univariable Analysis | Multivariable Analysis | |||||
|---|---|---|---|---|---|---|---|---|
| pCR (N = 22) | Residual Tumor (N = 60) | OR | 95% CI | p Value | OR | 95% CI | p Value | |
| T stage | ||||||||
| 1–2 | 20 (38%) | 33 (62%) | 8.18 | 1.75–38.16 | 0.007 | |||
| 3–4 | 2 (7%) | 27 (93%) | 1.00 | |||||
| Clinical anatomic stage | ||||||||
| IIA-IIIA | 20 (30%) | 46 (70%) | 3.04 | 0.63–14.66 | 0.165 | |||
| IIIB-IIIC | 2 (12%) | 14 (88%) | 1.00 | |||||
| Histologic grade | ||||||||
| 1-2 | 8 (21%) | 30 (79%) | 1.00 | |||||
| 3 | 11 (28%) | 28 (72%) | 1.47 | 0.52–4.19 | 0.468 | |||
| Missing | 3 | 2 | ||||||
| Ki-67 | ||||||||
| Low, <30% | 8 (24%) | 25 (76%) | 1.00 | |||||
| High, ≥30% | 9 (20%) | 35 (80%) | 0.80 | 0.27–2.37 | 0.692 | |||
| Missing | 5 | 0 | ||||||
| ER status | ||||||||
| Positive | 4 (15%) | 23 (85%) | 1.00 | 1.00 | ||||
| Negative | 18 (33%) | 37 (67%) | 2.80 | 0.84–9.31 | 0.093 | 8.37 | 1.75–40.1 | 0.008 |
| Metabolic parameters a | ||||||||
| MTVcsc (cm3) | 0.9 (0.3–1.7) | 2.8 (0.1–38.0) | 0.37 | 0.19–0.72 | 0.003 | 0.12 | 0.02–0.74 | 0.022 |
| TLGcsc | 5.0 (1.4–38.5) | 19.5 (0.2–593.4) | 0.92 | 0.86–0.98 | 0.010 | |||
| CSC proportion (%) | 15.9 (6.1–31.8) | 21.5 (5.6–39.1) | 0.94 | 0.87–1.00 | 0.068 | |||
| SUVmax | 6.3 (3.3–28.7) | 8.2 (3.4–23.7) | 0.93 | 0.82–1.06 | 0.271 | |||
| MTV (cm3) | 4.6 (1.7–13.8) | 13.3 (1.2–170.7) | 0.85 | 0.76–0.95 | 0.005 | |||
| TLG | 17.1 (4.7–111.3) | 57.6 (3.3–1386.9) | 0.98 | 0.96–0.99 | 0.011 | |||
| MTV40% (cm3) | 3.6 (1.7–13.8) | 9.1 (1.2–90.3) | 0.78 | 0.65–0.93 | 0.005 | |||
| TLG40% | 16.0 (4.7–66.3) | 47.0 (3.3–1006.1) | 0.96 | 0.94–0.99 | 0.008 | |||
| HER2-Positive/TN (N = 82) | Luminal (N = 38) | |||
|---|---|---|---|---|
| pCR (N = 22) | Residual Tumor (N = 60) | pCR (N = 0) | Residual Tumor (N = 38) | |
| MTVcsc < 1.75 cm3 | 22 | 21 | 0 | 27 |
| MTVcsc > 1.75 cm3 | 0 | 39 | 0 | 11 |
| Prediction accuracy | 74% (61/82) | 29% (11/38) | ||
| Anthracycline and Taxane Based (N = 57) | Taxane Based (N = 18) | Anthracycline Based (N = 7) | ||||
| pCR (N = 16) | Residual Tumor (N = 41) | pCR (N = 6) | Residual Tumor (N = 12) | pCR (N = 0) | Residual Tumor (N = 7) | |
| MTVcsc < 1.75 cm3 | 16 | 14 | 6 | 5 | 0 | 2 |
| MTVcsc > 1.75 cm3 | 0 | 27 | 0 | 7 | 0 | 5 |
| Prediction accuracy | 75% (43/57) | 72% (13/18) | 71% (5/7) | |||
| with Anti-HER2 NAC (N = 31) | without Anti-HER2 NAC (N = 26) | |||||
| pCR (N = 12) | Residual Tumor (N = 19) | pCR (N = 4) | Residual Tumor (N = 22) | |||
| MTVcsc < 1.75 cm3 | 12 | 5 | 4 | 13 | ||
| MTVcsc > 1.75 cm3 | 0 | 14 | 0 | 9 | ||
| Prediction accuracy | 84% (26/31) | 50% (13/26) | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Han, S.-A.; Won, K.Y.; Hong, I.K.; Kim, D.Y. Early Prediction of Tumor Response to Neoadjuvant Chemotherapy and Clinical Outcome in Breast Cancer Using a Novel FDG-PET Parameter for Cancer Stem Cell Metabolism. J. Pers. Med. 2020, 10, 132. https://doi.org/10.3390/jpm10030132
Kim C, Han S-A, Won KY, Hong IK, Kim DY. Early Prediction of Tumor Response to Neoadjuvant Chemotherapy and Clinical Outcome in Breast Cancer Using a Novel FDG-PET Parameter for Cancer Stem Cell Metabolism. Journal of Personalized Medicine. 2020; 10(3):132. https://doi.org/10.3390/jpm10030132
Chicago/Turabian StyleKim, Chanwoo, Sang-Ah Han, Kyu Yeoun Won, Il Ki Hong, and Deog Yoon Kim. 2020. "Early Prediction of Tumor Response to Neoadjuvant Chemotherapy and Clinical Outcome in Breast Cancer Using a Novel FDG-PET Parameter for Cancer Stem Cell Metabolism" Journal of Personalized Medicine 10, no. 3: 132. https://doi.org/10.3390/jpm10030132
APA StyleKim, C., Han, S.-A., Won, K. Y., Hong, I. K., & Kim, D. Y. (2020). Early Prediction of Tumor Response to Neoadjuvant Chemotherapy and Clinical Outcome in Breast Cancer Using a Novel FDG-PET Parameter for Cancer Stem Cell Metabolism. Journal of Personalized Medicine, 10(3), 132. https://doi.org/10.3390/jpm10030132