Differential Microbial Pattern Description in Subjects with Autoimmune-Based Thyroid Diseases: A Pilot Study

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Anthropometric and Laboratory Measurements
2.3. DNA Extraction from Faecal Samples
2.4. Sequencing of 16S rRNA and Bioinformatic Analysis
2.5. Statistical Analysis
3. Results
3.1. Clinical Data Study
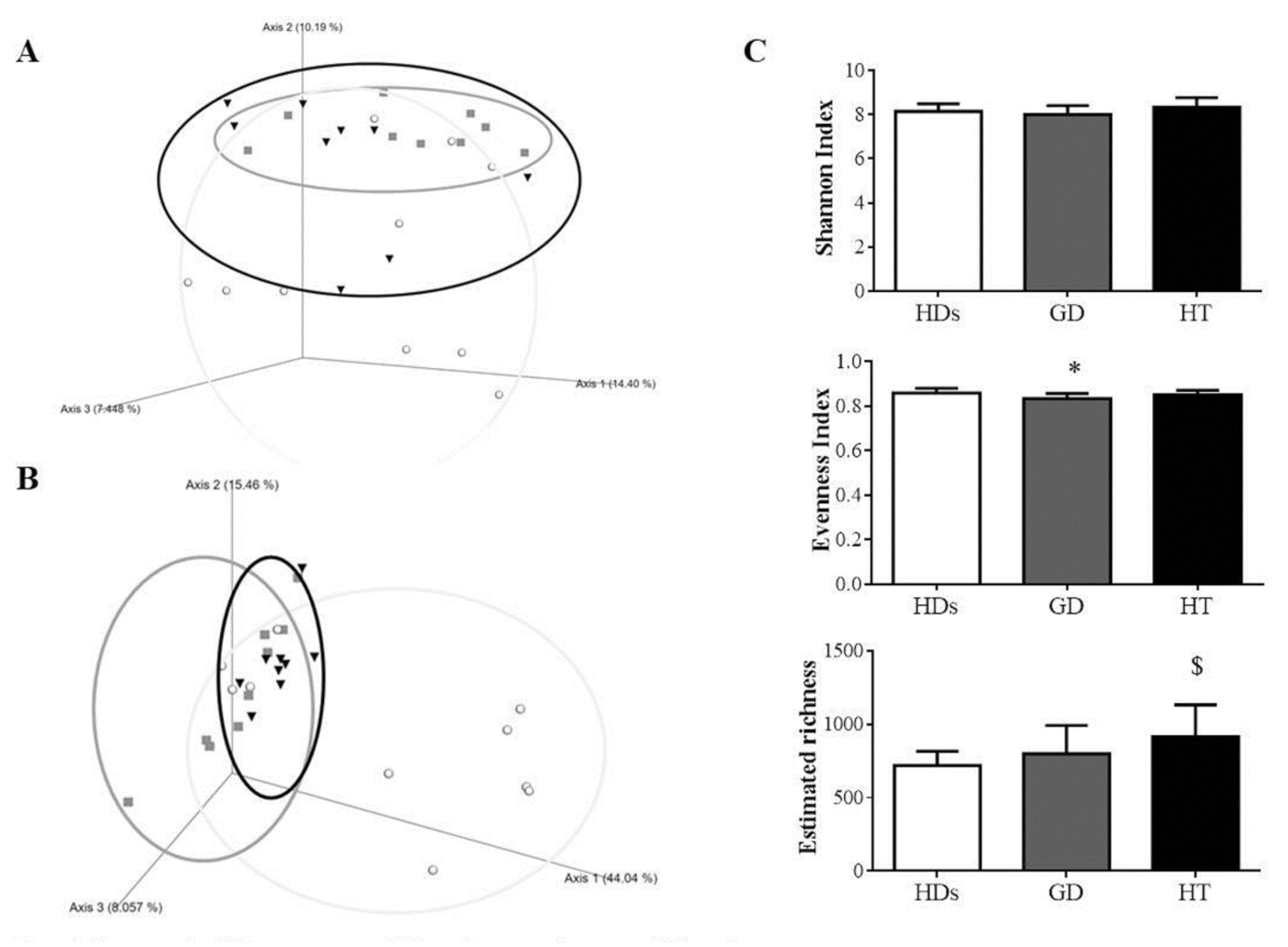
3.2. Gut Microbiota Diversity in AITD Patients
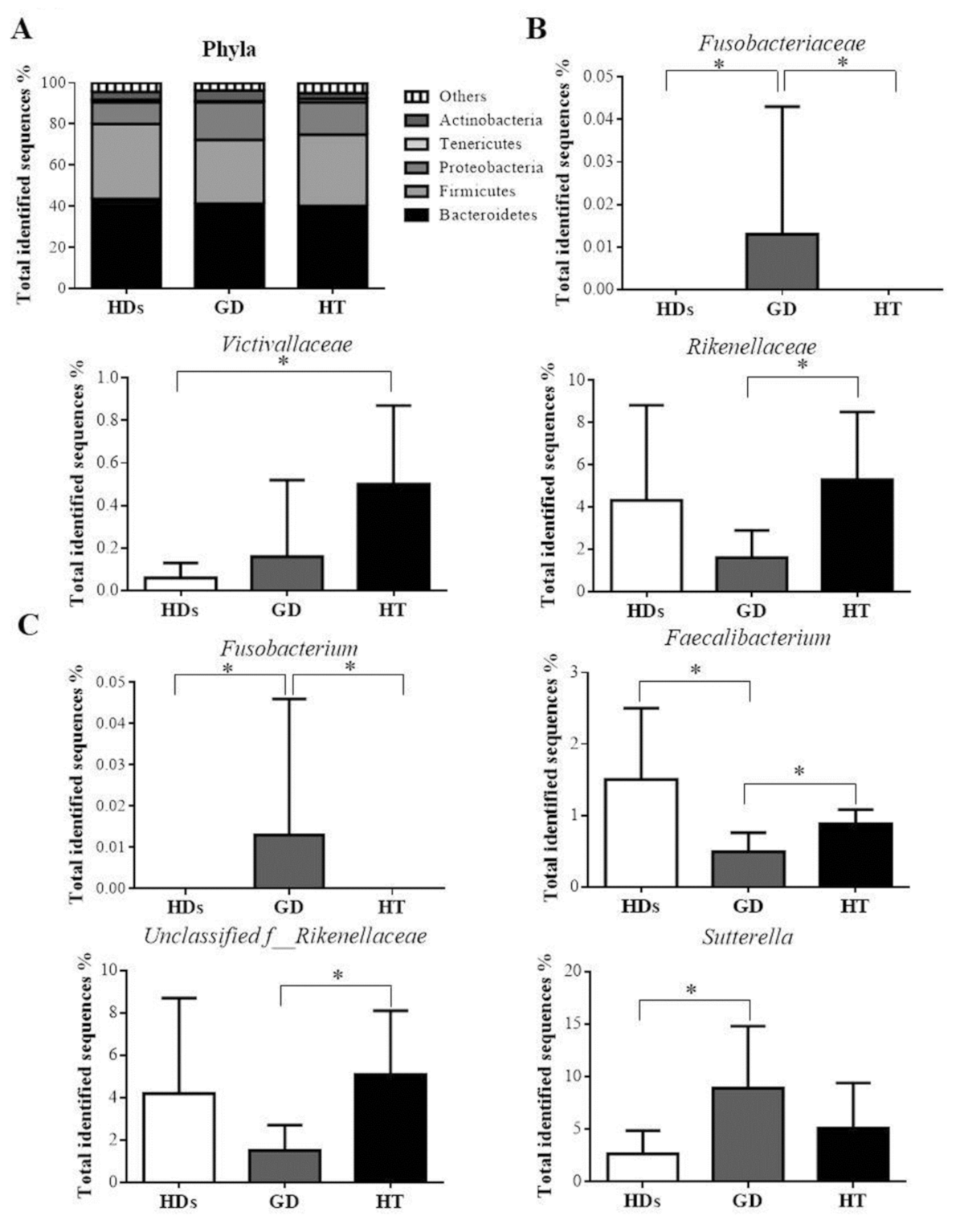
3.3. Gut Microbiota Profile in AITD Patients
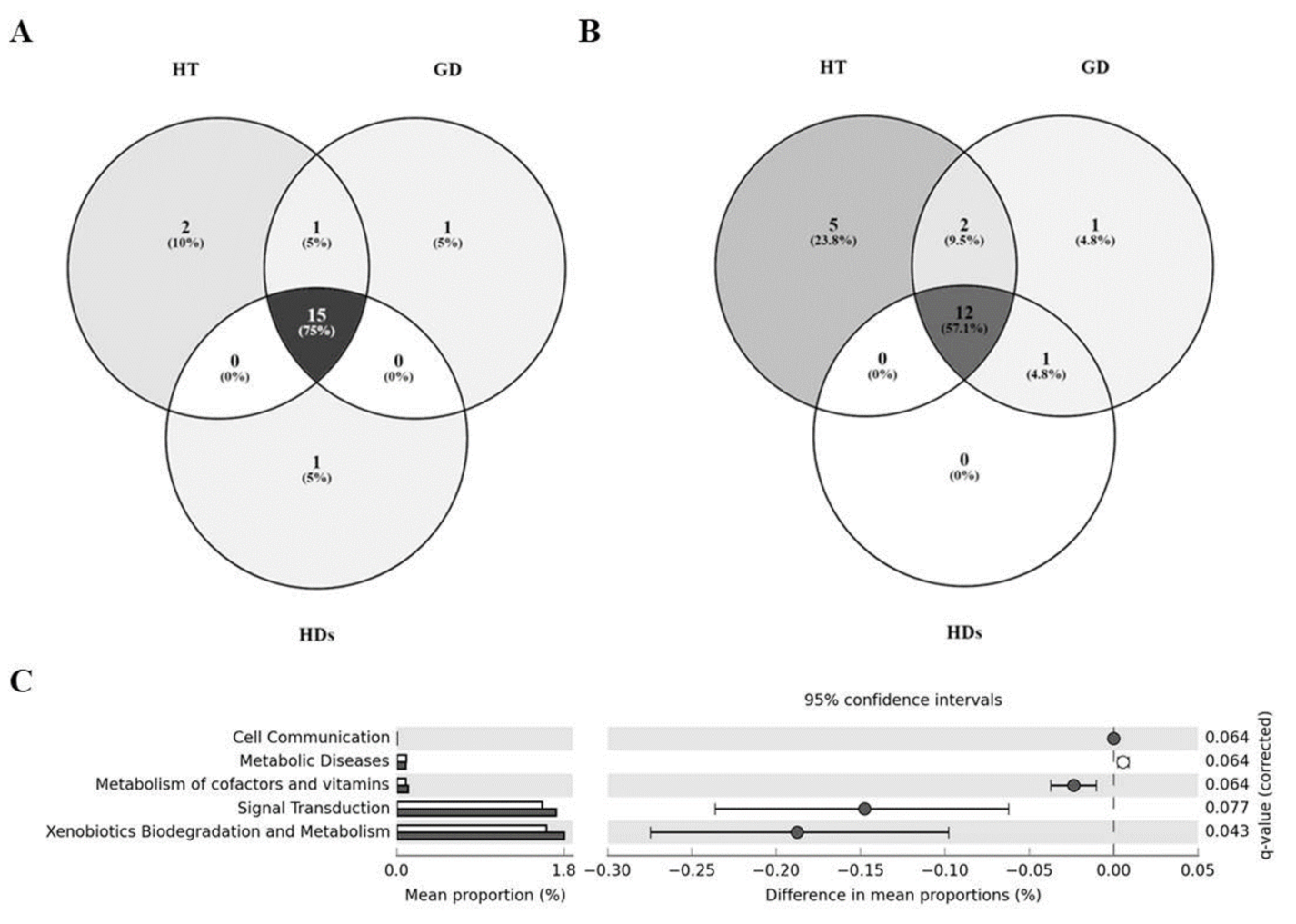
3.4. Core Microbiome in AITD Patients and HDs
3.5. Differences in the Metabolic Profiles of Gut Microbiota between AITD Patients and HDs
3.6. AITDs Could Be Related to Bacterial Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pearce, E.N.; Farwell, A.P.; Braverman, L.E. Thyroiditis. N. Engl. J. Med. 2003, 348, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Devdhar, M.; Ousman, Y.H.; Burman, K.D. Hypothyroidism. Endocrinol. Metab. Clin. North Am. 2007, 36, 595–615. [Google Scholar] [CrossRef] [PubMed]
- De Leo, S.; Lee, S.Y.; Braverman, L.E. Hyperthyroidism. Lancet 2016, 388, 906–918. [Google Scholar] [CrossRef] [Green Version]
- Hasham, A.; Tomer, Y. Genetic and epigenetic mechanisms in thyroid autoimmunity. Immunol. Res. 2012, 54, 204–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozzoli, R.; Barzilai, O.; Ram, M.; Villalta, D.; Bizzaro, N.; Sherer, Y.; Shoenfeld, Y. Infections and autoimmune thyroid diseases: Parallel detection of antibodies against pathogens with proteomic technology. Autoimmun. Rev. 2008, 8, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Prummel, M.F.; Strieder, T.; Wiersinga, W.M. The environment and autoimmune thyroid diseases. Eur. J. Endocrinol. 2004, 150, 605–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, E.C. The Thyroid and the Gut. J. Clin. Gastroenterol. 2010, 44, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Behrouzi, A.; Nafari, A.H.; Siadat, S.D. The significance of microbiome in personalized medicine. Clin. Transl. Med. 2019, 8, 16. [Google Scholar] [CrossRef]
- Leiva-Gea, I.; Sánchez-Alcoholado, L.; Martín-Tejedor, B.; Castellano-Castillo, D.; Moreno-Indias, I.; Urda-Cardona, A.; Tinahones, F.J.; Fernández-García, J.C.; Queipo-Ortuño, M.I. Gut Microbiota Differs in Composition and Functionality Between Children With Type 1 Diabetes and MODY2 and Healthy Control Subjects: A Case-Control Study. Diabetes Care 2018, 41, 2385–2395. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, N.M.; Ruiz-Limón, P.; Moreno-Indias, I.; Manrique-Arija, S.; Tinahones, F.J.; Fernandez-Nebro, A. Expansion of Rare and Harmful Lineages is Associated with Established Rheumatoid Arthritis. J. Clin. Med. 2020, 9, 1044. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.J.; Costenbader, K. Epigenetics and the microbiome: Developing areas in the understanding of the aetiology of lupus. Lupus 2014, 23, 505–506. [Google Scholar] [CrossRef] [PubMed]
- Sellitto, M.; Bai, G.; Serena, G.; Fricke, W.F.; Sturgeon, C.; Gajer, P.; White, J.R.; Koenig, S.S.K.; Sakamoto, J.; Boothe, D.; et al. Proof of Concept of Microbiome-Metabolome Analysis and Delayed Gluten Exposure on Celiac Disease Autoimmunity in Genetically At-Risk Infants. PLoS ONE 2012, 7, e33387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.S.; Gill, H.S. Immunostimulatory probiotic Lactobacillus rhamnosus HN001 and Bifidobacterium lactis HN019 do not induce pathological inflammation in mouse model of experimental autoimmune thyroiditis. Int. J. Food Microbiol. 2005, 103, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Penhale, W.J.; Young, P.R. The influence of the normal microbial flora on the susceptibility of rats to experimental autoimmune thyroiditis. Clin. Exp. Immunol. 1988, 72, 288–292. [Google Scholar]
- Ishaq, H.M.; Mohammad, I.S.; Guo, H.; Shahzad, M.; Hou, Y.J.; Ma, C.; Naseem, Z.; Wu, X.; Shi, P.; Xu, J. Molecular estimation of alteration in intestinal microbial composition in Hashimoto’s thyroiditis patients. Biomed. Pharmacother. 2017, 95, 865–874. [Google Scholar] [CrossRef]
- Zhao, F.; Feng, J.; Li, J.; Zhao, L.; Liu, Y.; Chen, H.; Jin, Y.; Zhu, B.; Wei, Y. Alterations of the Gut Microbiota in Hashimoto’s Thyroiditis Patients. Thyroid. 2018, 28, 175–186. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Benjamini-1995.pdf. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasso, F.C.; Carbonara, O.; Torella, R.; Mezzogiorno, A.; Esposito, V.; DeMagistris, L.; Secondulfo, M.; Carratu’, R.; Iafusco, D.; Cartenì, M. Ultrastructural changes in enterocytes in subjects with Hashimoto’s thyroiditis. Gut 2004, 53, 1878–1880. [Google Scholar] [CrossRef] [Green Version]
- Virili, C.; Fallahi, P.; Antonelli, A.; Benvenga, S.; Centanni, M. Gut microbiota and Hashimoto’s thyroiditis. Rev. Endocr. Metab. Disord. 2018, 19, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut As a Danger Signal for Autoimmune Diseases. Front. Immunol. 2017, 8, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, E.C.; Bilotta, A.L.; Gabrielli, M.; Scarpellini, E.; Lupascu, A.; Laginestra, A.; Novi, M.; Sottili, S.; Serricchio, M.; Cammarota, G.; et al. Association between Hypothyroidism and Small Intestinal Bacterial Overgrowth. J. Clin. Endocrinol. Metab. 2007, 92, 4180–4184. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, U.C.; Shukla, R.; Ghoshal, U.; Gwee, K.-A.; Ng, S.C.; Quigley, E.M.M. The Gut Microbiota and Irritable Bowel Syndrome: Friend or Foe? Int. J. Inflamm. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Ishaq, H.M.; Mohammad, I.S.; Shahzad, M.; Ma, C.; Raza, M.A.; Wu, X.; Guo, H.; Shi, P.; Xu, J. Molecular Alteration Analysis of Human Gut Microbial Composition in Graves’ disease Patients. Int. J. Biol. Sci. 2018, 14, 1558–1570. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Soldan, M.M.P.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Redline, R.W.; Han, Y.W. Fusobacterium nucleatum Induces Fetal Death in Mice via Stimulation of TLR4-Mediated Placental Inflammatory Response. J. Immunol. 2007, 179, 2501–2508. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, X.; Ahmed, A.; Wu, D.; Liu, L.; Qiu, J.; Yan, Y.; Jin, M.; Xin, Y. Gut Microbe Analysis Between Hyperthyroid and Healthy Individuals. Curr. Microbiol. 2014, 69, 675–680. [Google Scholar] [CrossRef]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, A.; Lennon, G.; O’Sullivan, O.; Docherty, N.; Balfe, A.; Maguire, A.; Mulcahy, H.E.; Doherty, G.; O’Donoghue, D.; Hyland, J.; et al. Spatial variation of the colonic microbiota in patients with ulcerative colitis and control volunteers. Gut 2015, 64, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Hiippala, K.; Kainulainen, V.; Kalliomäki, M.; Arkkila, P.; Satokari, R. Mucosal Prevalence and Interactions with the Epithelium Indicate Commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar] [CrossRef] [Green Version]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.C.; Wang, Z.; Usyk, M.; Sotres-Alvarez, D.; Daviglus, M.L.; Schneiderman, N.; Talavera, G.A.; Gellman, M.D.; Thyagarajan, B.; Moon, J.-Y.; et al. Gut microbiome composition in the Hispanic Community Health Study/Study of Latinos is shaped by geographic relocation, environmental factors, and obesity. Genome Biol. 2019, 20, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Gevers, D.; Siljander, H.; Vatanen, T.; Hyötyläinen, T.; Hämäläinen, A.-M.; Peet, A.; Tillmann, V.; Pöhö, P.; Mattila, I.; et al. The Dynamics of the Human Infant Gut Microbiome in Development and in Progression toward Type 1 Diabetes. Cell Host Microbe 2015, 17, 260–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croxford, J.L.; Anger, H.A.; Miller, S.D. Viral Delivery of an Epitope fromHaemophilus influenzaeInduces Central Nervous System Autoimmune Disease by Molecular Mimicry. J. Immunol. 2005, 174, 907–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 1–11. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Prummel, M.F.; Wiersinga, W.M. Thyroid peroxidase autoantibodies in euthyroid subjects. Best Pr. Res. Clin. Endocrinol. Metab. 2005, 19, 1–15. [Google Scholar] [CrossRef]
- Gelfand, R.A.; Hutchinson-Williams, K.A.; Bonde, A.A.; Castellino, P.; Sherwin, R.S. Catabolic effects of thyroid hormone excess: The contribution of adrenergic activity to hypermetabolism and protein breakdown. Metabolism 1987, 36, 562–569. [Google Scholar] [CrossRef]
- Koppel, N.; Rekdal, V.M.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, eaag2770. [Google Scholar] [CrossRef] [PubMed]
- Punzalan, C.; Qamar, A. Probiotics for the Treatment of Liver Disease. In The Microbiota in Gastrointestinal Pathophysiology: Implications for Human Health, Prebiotics, Probiotics, and Dysbiosis, 1st ed.; Elsevier Inc.: London, UK, 2017; pp. 373–381. ISBN 9780128040621. [Google Scholar]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira-Halder, C.V.; Faria, A.V.D.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | HT Patients (n = 9) | GD Patients (n = 9) | HDs (n = 11) | p-Value |
|---|---|---|---|---|
| Sex (M/F, % F) | (0/10, 100) | (2/7, 77.8) | (4/7, 63.5) | 0.113 |
| Age (years, mean ± SD) | 40.3 ± 9.6 | 46.2 ± 8.6 | 48.8 ± 6.2 | 0.062 |
| Smokers (%) | 30 | 44.4 | ----- | 0.515 |
| Family history of Thyroid disease (%) | 50 | 33.3 | ----- | 0.484 |
| Time of evolution of thyroid disease (months, mean ± SD) | 134.4 ± 101.3 | 16.4 ± 22.6 | ----- | 0.001 * |
| Anthropometry | ||||
| Weight (kg, mean ± SD) | 63.4 ± 11.7 | 66.6 ± 13.0 | 69.5 ± 8.1 | 0.354 |
| BMI (kg/m2, mean ± SD) | 24.9 ± 5.8 | 25.2 ± 4.7 | 25.0 ± 2.0 | 0.831 |
| Waist c. (cm, mean ± SD) | 81.4 ± 13.8 | 86.1 ± 10.9 | 87.9 ± 8.4 | 0.321 |
| Hip c. (cm, mean ± SD) | 98.3 ± 10.7 | 101.6 ± 11.3 | 97.0 ± 4.1 | 0.391 |
| Blood pressure | ||||
| Systolic (mmHg, mean ± SD) | 121.6 ± 11.3 | 119.6 ± 16.2 | 128.8 ± 17.3 | 0.636 |
| Diastolic (mmHg, mean ± SD) | 75.3 ± 5.1 | 75.0 ± 15.4 | 78.6 ± 8.7 | 0.449 |
| Analytical metabolic | ||||
| Glucose (mg/dL, mean ± SD) | 86.6 ± 6.5 | 93.4 ± 4.1 | 91.0 ± 8.3 | 0.078 |
| HbA1c (%, mean ± SD) | 5.1 ± 0.3 | 5.3 ± 0.4 | 5.3 ± 0.3 | 0.29 |
| Total-C (mg/dL, mean ± SD) | 178.5 ± 44.7 | 192.2 ± 29.6 | 192.4 ± 47.8 | 0.568 |
| LDL-c (mg/dL, mean ± SD) | 107.7 ± 39.1 | 109.4 ± 10.2 | 110.1 ± 38.1 | 0.758 |
| HDL-c (mg/dL, mean ± SD) | 56.8 ± 11.7 | 65.6 ± 22.2 | 60.5 ± 12.9 | 0.666 |
| TGs (mg/dL, mean ± SD) | 70.1 ± 16.6 | 85.2 ± 31.9 | 108.8 ± 59.0 | 0.132 |
| CRP (mg/dL, mean ± SD) | 3.1 ± 0.0 | 3.2 ± 0.4 | 4.1 ± 1.4 | 0.116 |
| Thyroid profile | ||||
| TSH (µIU/mL, mean ± SD) | 2.6 ± 2.8 | 3.3 ± 8.5 | 2.2 ± 1.0 | 0.030 * |
| FT4 (pmol/L, mean ± SD) | 15.4 ± 2.2 | 15.2 ± 3.1 | 15.2 ± 1.3 | 0.76 |
| FT3 (pmol/L, mean ± SD) | 3.8 ± 0.2 | 5.5 ± 2.3 | 4.8 ± 0.4 | 0.002 * |
| TPO-Ab (IU/mL, mean ± SD) | 1186.7 ± 358.4 | 792.0 ± 621.7 | 160.3 ± 381.3 | 0.001 * |
| TPO-Ab > 60 IU/mL (P/N, % P) | (10/0, 100) | (6/3, 66.7) | (2/9, 18.2) | 0.000 * |
| TSI-Ab (IU/mL, mean ± SD) | 3.5 ± 7.2 | 16.8 ± 34.1 | 0.8 ± 0.1 | 0.000 * |
| TSI-Ab > 2 IU/mL (P/N, % P) | (1/9, 10) | (9/0, 100) | (0/11, 0) | 0.000 * |
| Plylum Level | Family or Genus Level | TPO-Ab | TSI-Ab |
|---|---|---|---|
| Bacteroidetes | ----- | ----- | |
| Alistipes | r = 0.432; p = 0.019 | ----- | |
| Firmicutes | ----- | ----- | |
| Lactobacillaceae | ----- | r = 0.517; p = 0.006 | |
| Lactobacillus | ----- | r = 0.517; p = 0.006 | |
| Faecalibacterium | r = −0.453; p = 0.014 | r = −0.406; p = 0.036 | |
| Ruminococcaceae unclasificated | r = 0.408; p = 0.028 | ----- | |
| Proteobacteria | ----- | ----- | |
| Enterobacteriaceae | r = 0.416; p = 0.025 | ----- | |
| Pasteurellaceae | ----- | r = 0.441; p = 0.021 |
| Positive TPO Autoimmunity (TPO-Ab >60 IU/mL) | ||
|---|---|---|
| Ruminococcaeae unclassified | ||
| OR (CI) | p | |
| Crude model | 1.494 (1.041–2.145) | 0.029 |
| Model 1 | 1.474 (1.026–2.413) | 0.038 |
| Positive TSI Autoimmunity (TSI-Ab >2 IU/mL) | ||||
|---|---|---|---|---|
| Sutterella | Faecalibacterium | |||
| OR (CI) | p | OR (CI) | p | |
| Crude model | 1.351 (1.053–1.734) | 0.018 | 0.059 (0.004–0.958) | 0.047 |
| Model 1 | 1.499 (1.041–2.158) | 0.030 | 0.025 (0.001–0.900) | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cornejo-Pareja, I.; Ruiz-Limón, P.; Gómez-Pérez, A.M.; Molina-Vega, M.; Moreno-Indias, I.; Tinahones, F.J. Differential Microbial Pattern Description in Subjects with Autoimmune-Based Thyroid Diseases: A Pilot Study. J. Pers. Med. 2020, 10, 192. https://doi.org/10.3390/jpm10040192
Cornejo-Pareja I, Ruiz-Limón P, Gómez-Pérez AM, Molina-Vega M, Moreno-Indias I, Tinahones FJ. Differential Microbial Pattern Description in Subjects with Autoimmune-Based Thyroid Diseases: A Pilot Study. Journal of Personalized Medicine. 2020; 10(4):192. https://doi.org/10.3390/jpm10040192
Chicago/Turabian StyleCornejo-Pareja, Isabel, Patricia Ruiz-Limón, Ana M. Gómez-Pérez, María Molina-Vega, Isabel Moreno-Indias, and Francisco J. Tinahones. 2020. "Differential Microbial Pattern Description in Subjects with Autoimmune-Based Thyroid Diseases: A Pilot Study" Journal of Personalized Medicine 10, no. 4: 192. https://doi.org/10.3390/jpm10040192
APA StyleCornejo-Pareja, I., Ruiz-Limón, P., Gómez-Pérez, A. M., Molina-Vega, M., Moreno-Indias, I., & Tinahones, F. J. (2020). Differential Microbial Pattern Description in Subjects with Autoimmune-Based Thyroid Diseases: A Pilot Study. Journal of Personalized Medicine, 10(4), 192. https://doi.org/10.3390/jpm10040192