
MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. DNA Sequence Analysis of MICA
2.3. Determination of Soluble MICA (sMICA) Levels in Serum Samples
2.4. Generation of Human MICA Expression Constructs
2.5. Generation of Cell Lines Expressing MICA Alleles
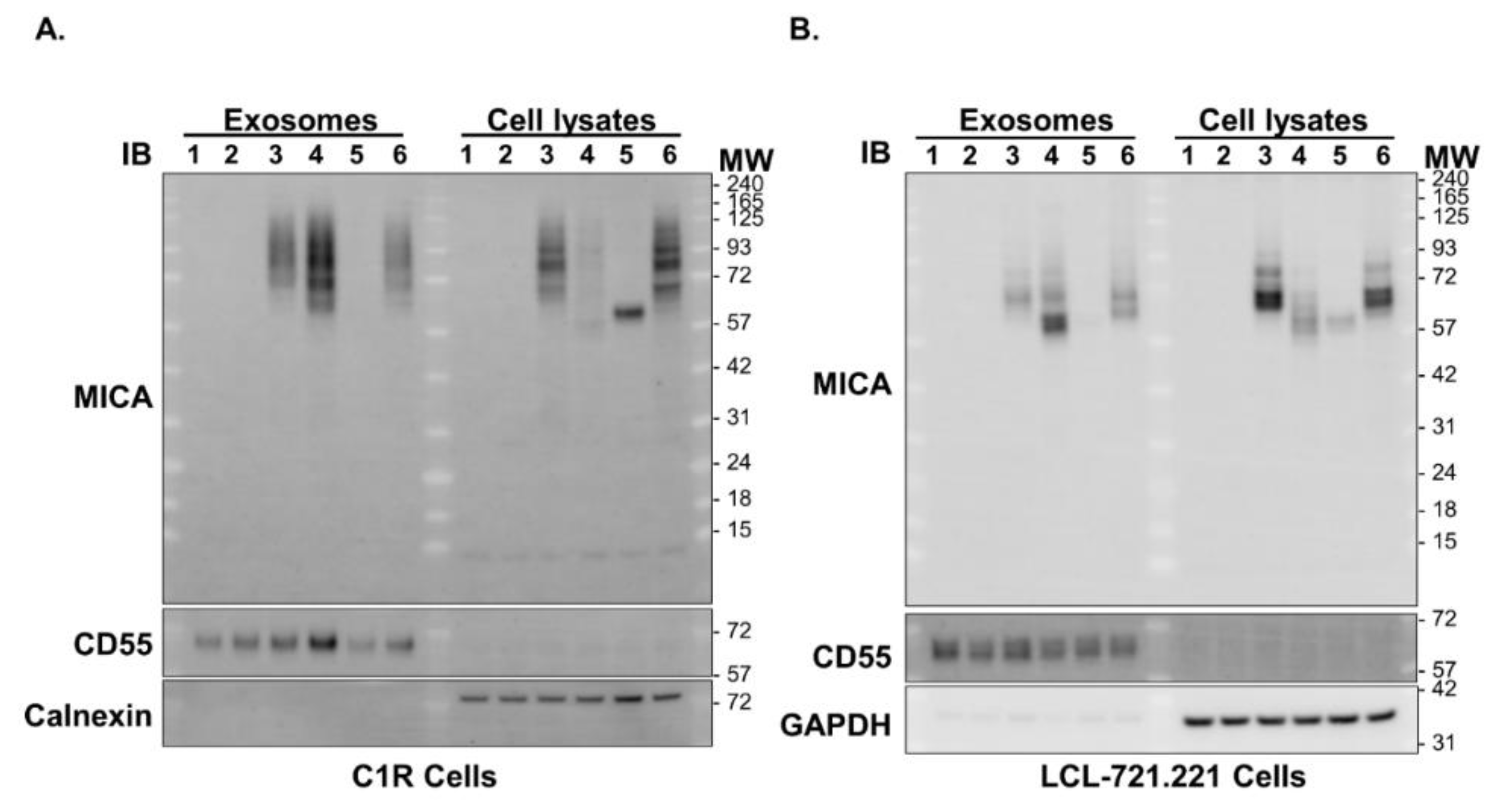
2.6. Western Blot Analyses and Detection of sMICA, Exosomal MICA and Cellular MICA
2.7. Statistical Analysis
2.8. Supplementary Methods
3. Results
3.1. MICA cSNP and Alleles in Taiwanese
3.2. Association of MICA cSNPs with AS Susceptibility in Taiwanese
3.3. Association of MICA Alleles with AS Susceptibility in Taiwanese
3.4. Associations of MICA Alleles with Syndesmophyte Formation in AS Patients
3.5. Associations of MICA Alleles with HLA-B27 Positivity in AS Patients
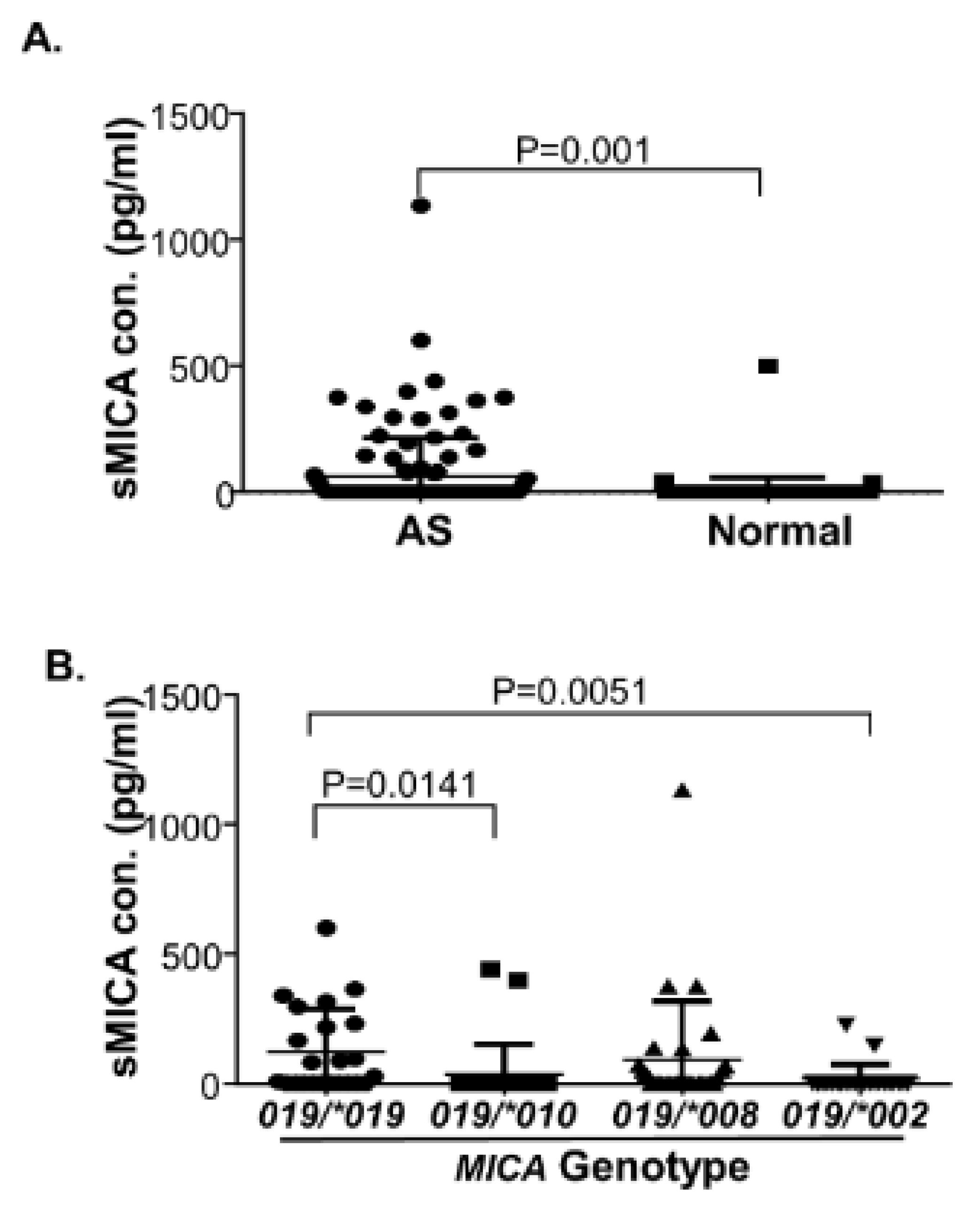
3.6. Increased Levels of Serum sMICA in AS Patients and MICA*019 Homozygous AS Patients
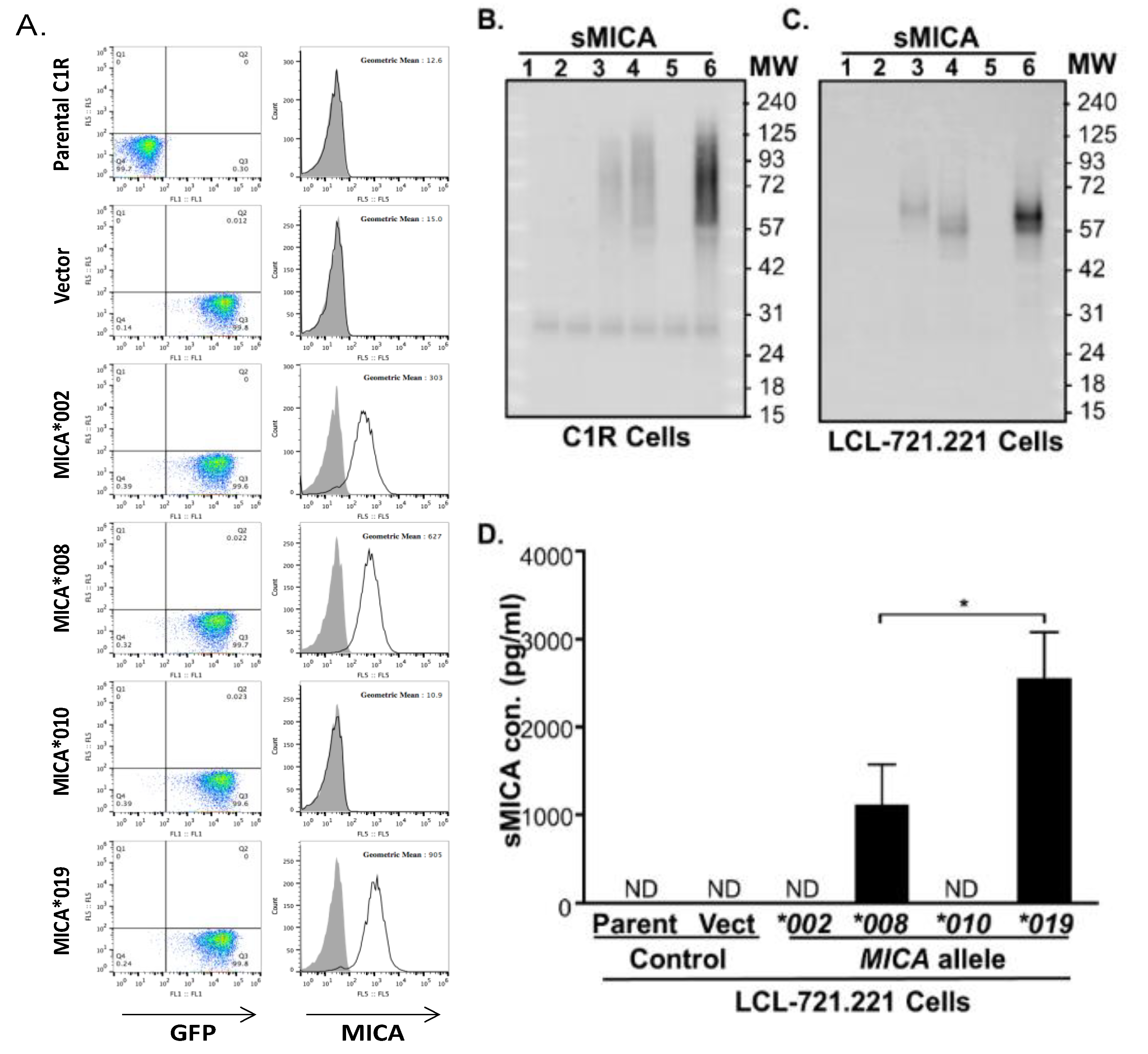
3.7. Cells Expressing MICA*019 Allele Produces the Highest Amount of sMICA In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AS | ankylosing spondylitis |
| SpA | spondyloarthritides; |
| mSASSS | modified Stoke Ankylosing Spondylitis Spinal Score |
| HLA-B27 | human leukocyte antigen B27 |
| MICA | major histocompatibility complex class I chain-related gene A |
| NKG2D | receptor natural killer group 2, member D |
| SNP | single nucleotide polymorphism |
| sMICA | soluble MICA |
References
- Taurog, J.D.; Chhabra, A.; Colbert, R.A. Ankylosing Spondylitis and Axial Spondyloarthritis. N. Engl. J. Med. 2016, 374, 2563–2574. [Google Scholar] [CrossRef] [Green Version]
- Rezaiemanesh, A.; Abdolmaleki, M.; Abdolmohammadi, K.; Aghaei, H.; Pakdel, F.D.; Fatahi, Y.; Soleimanifar, N.; Zavvar, M.; Nicknam, M.H. Immune cells involved in the pathogenesis of ankylosing spondylitis. Biomed. Pharmacother. 2018, 100, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Dalbeth, N.; Gundle, R.; Davies, R.J.; Lee, Y.C.; McMichael, A.J.; Callan, M.F. CD56bright NK cells are enriched at inflammatory sites and can engage with monocytes in a reciprocal program of activation. J. Immunol. 2004, 173, 6418–6426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, N.; Pascal, V.; Fasth, A.E.; Sundström, Y.; Galsgaard, E.D.; Ahern, D.; Andersen, M.; Baslund, B.; Bartels, E.M.; Bliddal, H.; et al. Balance between activating NKG2D, DNAM-1, NKp44 and NKp46 and inhibitory CD94/NKG2A receptors determine natural killer degranulation towards rheumatoid arthritis synovial fibroblasts. Immunology 2014, 142, 581–593. [Google Scholar] [CrossRef]
- Pridgeon, C.; Lennon, G.P.; Pazmany, L.; Thompson, R.N.; Christmas, S.E.; Moots, R.J. Natural killer cells in the synovial fluid of rheumatoid arthritis patients exhibit a CD56bright, CD94bright, CD158negative phenotype. Rheumatology 2003, 42, 870–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollock, R.A.; Chandran, V.; Pellett, F.J.; Thavaneswaran, A.; Eder, L.; Barrett, J.; Rahman, P.; Farewell, V.; Gladman, D.D. The functional MICA-129 polymorphism is associated with skin but not joint manifestations of psoriatic disease independently of HLA-B and HLA-C. Tissue Antigens 2013, 82, 43–47. [Google Scholar] [CrossRef]
- Kim, T.-J.; Lee, S.-J.; Cho, Y.-N.; Park, S.-C.; Jin, H.-M.; Kim, M.-J.; Park, D.-J.; Kee, S.-J.; Lee, S.-S.; Park, Y.-W. Immune cells and bone formation in ankylosing spondylitis. Clin. Exp. Rheumatol. 2012, 30, 469–475. [Google Scholar]
- Szanto, S.; Aleksza, M.; Mihály, E.; Lakos, G.; Szabo, Z.; Végvári, A.; Sipka, S.; Szekanecz, Z. Intracytoplasmic cytokine expression and T cell subset distribution in the peripheral blood of patients with ankylosing spondylitis. J. Rheumatol. 2008, 35, 2372–2375. [Google Scholar] [CrossRef]
- Conigliaro, P.; Scrivo, R.; Valesini, G.; Perricone, R. Emerging role for NK cells in the pathogenesis of inflammatory arthropathies. Autoimmun. Rev. 2011, 10, 577–581. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- 1Zitti, B.; Bryceson, Y.T. Natural killer cells in inflammation and autoimmunity. Cytokine Growth Factor Rev. 2018, 42, 37–46. [Google Scholar]
- Tian, Z.; Gershwin, M.E.; Zhang, C. Regulatory NK cells in autoimmune disease. J. Autoimmun. 2012, 39, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Flodstrom-Tullberg, M.; Bryceson, Y.T.; Shi, F.D.; Hoglund, P.; Ljunggren, H.G. Natural killer cells in human autoimmunity. Curr. Opin. Immunol. 2009, 21, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Perricone, R.; Perricone, C.; De Carolis, C.; Shoenfeld, Y. NK cells in autoimmunity: A two-edg’d weapon of the immune system. Autoimmun. Rev. 2008, 7, 384–390. [Google Scholar] [CrossRef]
- Gianchecchi, E.; Delfino, D.V.; Fierabracci, A. NK cells in autoimmune diseases: Linking innate and adaptive immune responses. Autoimmun. Rev. 2018, 17, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Lunemann, A.; Lunemann, J.D.; Munz, C. Regulatory NK-cell functions in inflammation and autoimmunity. Mol. Med. 2009, 15, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.K.C.; Lam, Q.L.K.; Sun, L.; Wang, S.; Ko, K.-H.; Xu, H.; Wu, C.-Y.; Zheng, B.-J.; Lu, L. Natural killer cell degeneration exacerbates experimental arthritis in mice via enhanced interleukin-17 production. Arthritis Rheum. 2008, 58, 2700–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanovic, A.; Correia, M.P.; Cerwenka, A. The NKG2D/NKG2DL Axis in the Crosstalk Between Lymphoid and Myeloid Cells in Health and Disease. Front. Immunol. 2018, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Groh, V.; Wu, J.; Steinle, A.; Phillips, J.H.; Lanier, L.L.; Spies, T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 1999, 285, 727–729. [Google Scholar] [CrossRef]
- Babic, M.; Romagnani, C. The Role of Natural Killer Group 2, Member D in Chronic Inflammation and Autoimmunity. Front. Immunol. 2018, 9, 1219. [Google Scholar] [CrossRef]
- Chen, D.; Gyllensten, U. MICA polymorphism: Biology and importance in cancer. Carcinogenesis 2014, 35, 2633–2642. [Google Scholar] [CrossRef]
- Yang, X.; Kuang, S.; Wang, L.; Wei, Y. MHC class I chain-related A: Polymorphism, regulation and therapeutic value in cancer. Biomed. Pharmacother. 2018, 103, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Li, H.; Couturier, J.P.; Yang, K.; Guo, X.; He, N.; E Lewis, R.; Zhou, X. Allele Specific Expression of MICA Variants in Human Fibroblasts Suggests a Pathogenic Mechanism. Open Rheumatol. J. 2015, 9, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhou, X. Associations of MICA Polymorphisms with Inflammatory Rheumatic Diseases. Open Rheumatol. J. 2015, 9, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Wang, J.; Zou, H.; Ward, M.M.; Weisman, M.H.; Espitia, M.G.; Xiao, X.; Petersdorf, E.; Mignot, E.; Martin, J.; et al. MICA, a gene contributing strong susceptibility to ankylosing spondylitis. Ann. Rheum. Dis. 2014, 73, 1552–1557. [Google Scholar] [CrossRef]
- Evans, D.M.; A Spencer, C.C.; Pointon, J.J.; Su, Z.; Harvey, D.; Kochan, G.; Oppermann, U.; Dilthey, A.; Pirinen, M.; A Stone, M.; et al. Interaction between ERAP1 and HLA-B27 in ankylosing spondylitis implicates peptide handling in the mechanism for HLA-B27 in disease susceptibility. Nat. Genet. 2011, 43, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Van der Linden, S.; Valkenburg, H.A.; Cats, A. Evaluation of diagnostic criteria for ankylosing spondylitis. A proposal for modification of the New York criteria. Arthritis Rheum. 1984, 27, 361–368. [Google Scholar] [CrossRef]
- Creemers, M.C.; Franssen, M.J.; van’t Hof, M.A.; Gribnau, F.W.; van de Putte, L.B.; van Riel, P.L. Assessment of outcome in ankylosing spondylitis: An extended radiographic scoring system. Ann. Rheum. Dis. 2005, 64, 127–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-M.; Ho, H.-H.; Chang, S.-W.; Wu, Y.-J.J.; Lin, J.-C.; Chang, P.-Y.; Wu, J.; Chen, J.-Y. ERAP1 genetic variations associated with HLA-B27 interaction and disease severity of syndesmophytes formation in Taiwanese ankylosing spondylitis. Arthritis Res. Ther. 2012, 14, R125. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Groh, V.; Strong, R.K.; Spies, T. A single amino acid substitution causes loss of expression of a MICA allele. Immunogenetics 2000, 51, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Bei, J.-X.; Shen, M.; Li, Q.; Liao, Z.; Zhang, Y.; Lv, Q.; Wei, Q.; Low, H.-Q.; Guo, Y.-M.; et al. A genome-wide association study in Han Chinese identifies new susceptibility loci for ankylosing spondylitis. Nat. Genet. 2011, 44, 73–77. [Google Scholar] [CrossRef]
- Cortes, A.; Hadler, J.; Pointon, J.P.; Robinson, P.C.; Karaderi, T.; Leo, P.; Cremin, K.; Pryce, K.; Harris, J.; Lee, S.; et al. Identification of multiple risk variants for ankylosing spondylitis through high-density genotyping of immune-related loci. Nat. Genet. 2013, 45, 730–738. [Google Scholar]
- Reveille, J.D.; Sims, A.M.; Danoy, P.; Evans, D.M.; Leo, P.; Pointon, J.J.; Jin, R.; Zhou, X.; Bradbury, L.A.; Appleton, L.H.; et al. Genome-wide association study of ankylosing spondylitis identifies non-MHC susceptibility loci. Nat. Genet. 2010, 42, 123–127. [Google Scholar]
- Choy, M.K.; Phipps, M.E. MICA polymorphism: Biology and importance in immunity and disease. Trends Mol. Med. 2010, 16, 97–106. [Google Scholar] [CrossRef]
- Zou, Y.; Stastny, P. Role of MICA in the immune response to transplants. Tissue Antigens 2010, 76, 171–176. [Google Scholar] [CrossRef]
- Wensveen, F.M.; Jelencic, V.; Polic, B. NKG2D: A Master Regulator of Immune Cell Responsiveness. Front. Immunol. 2018, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Isernhagen, A.; Malzahn, D.; Bickeboller, H.; Dressel, R. Impact of the MICA-129Met/Val Dimorphism on NKG2D-Mediated Biological Functions and Disease Risks. Front. Immunol. 2016, 7, 588. [Google Scholar] [CrossRef] [Green Version]
- Isernhagen, A.; Malzahn, D.; Viktorova, E.; Elsner, L.; Monecke, S.; Von Bonin, F.; Kilisch, M.; Wermuth, J.M.; Walther, N.; Balavarca, Y.; et al. The MICA-129 dimorphism affects NKG2D signaling and outcome of hematopoietic stem cell transplantation. EMBO Mol. Med. 2015, 7, 1480–1502. [Google Scholar] [CrossRef] [PubMed]
- Isernhagen, A.; Schilling, D.; Monecke, S.; Shah, P.; Elsner, L.; Walter, L.; Multhoff, G.; Dressel, R. The MICA-129Met/Val dimorphism affects plasma membrane expression and shedding of the NKG2D ligand MICA. Immunogenetics 2016, 68, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Zingoni, A.; Vulpis, E.; Cecere, F.; Amendola, M.G.; Fuerst, D.; Saribekyan, T.; Achour, A.; Sandalova, T.; Nardone, I.; Peri, A.; et al. MICA-129 Dimorphism and Soluble MICA Are Associated With the Progression of Multiple Myeloma. Front. Immunol. 2018, 9, 926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, E.; Collins, R.; Kondeatis, E.; Wallace, G.; Graham, E.; Vaughan, R.; Stanford, M. Associations of major histocompatibility complex class I chain-related molecule polymorphisms with Behcet’s disease in Caucasian patients. Tissue Antigens 2005, 66, 195–199. [Google Scholar] [CrossRef]
- Munoz-Saa, I.; Cambra, A.; Pallarés, L.; Espinosa, G.; Juan, A.; Pujalte, F.; Matamoros, N.; Mila, J.; Julià, M.R. Allelic diversity and affinity variants of MICA are imbalanced in Spanish patients with Behcet’s disease. Scand. J. Immunol. 2006, 64, 77–82. [Google Scholar] [CrossRef]
- Cortes, A.; Gladman, D.; Raychaudhuri, S.; Cui, J.; Wheeler, L.; Brown, M.A. Imputation-based analysis of MICA alleles in the susceptibility to ankylosing spondylitis. Ann. Rheum. Dis. 2018, 77, 1691–1692. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Wang, R.; Ward, M.M. Syndesmophyte growth in ankylosing spondylitis. Curr. Opin. Rheumatol. 2015, 27, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Boukouaci, W.; Al-Daccak, R.; Dulphy, N.; Lauden, L.; Amokrane, K.; Fortier, C.; Marzais, F.; Bennabi, M.; de Latour, R.P.; Socie, G.; et al. Soluble MICA-NKG2D interaction upregulates IFN-gamma production by activated CD3-CD56+ NK cells: Potential impact on chronic graft versus host disease. Hum. Immunol. 2013, 74, 1536–1541. [Google Scholar] [CrossRef] [PubMed]
- Mariaselvam, C.M.; Boukouaci, W.; Charron, D.; Krishnamoorthy, R.; Tamouza, R.; Misra, D.P.; Negi, V.S. Association of MICA-129 polymorphism and circulating soluble MICA level with rheumatoid arthritis in a south Indian Tamil population. Int. J. Rheum. Dis. 2018, 21, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Ashiru, O.; López-Cobo, S.; Fernández-Messina, L.; Pontes-Quero, S.; Pandolfi, R.; Reyburn, H.T.; Valés-Gómez, M. A GPI anchor explains the unique biological features of the common NKG2D-ligand allele MICA*008. Biochem. J. 2013, 454, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashiru, O.; Boutet, P.; Fernández-Messina, L.; Agüera-González, S.; Skepper, J.N.; Vales-Gomez, M.; Reyburn, H.T. Natural killer cell cytotoxicity is suppressed by exposure to the human NKG2D ligand MICA*008 that is shed by tumor cells in exosomes. Cancer Res. 2010, 70, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Clayton, A.; Mitchell, J.P.; Court, J.; Linnane, S.; Mason, M.D.; Tabi, Z. Human tumor-derived exosomes down-modulate NKG2D expression. J. Immunol. 2008, 180, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| MICA Allele | Estimated Frequency Trend Test | Logistic Regression | Logistic Regression Adjusted for Sex | ||||
|---|---|---|---|---|---|---|---|
| AS(2N = 1790) | Control (2N = 1792) | p Value | PFDR Value | OR (95% CI) | PFDR Value | OR (95% CI) | |
| MICA*019:01 | 765 (42.74%) | 157 (8.76%) | <0.00001 | 1.91×10−115 | 14.86 (11.80–18.71) | 2.25 × 10−115 | 14.90 (11.83–18.77) |
| MICA*008:01:01 | 332 (18.55%) | 510 (28.46%) | 2.0 × 10−12 | 1.4×10−11 | 0.57 (0.48–0.66) | 8.98 × 10−12 | 0.56 (0.48–0.66) |
| MICA*010:01 | 192 (10.73%) | 287 (16.02%) | 1.86 × 10−6 | 4.77×10−6 | 0.62 (0.50–0.75) | 4.50 × 10−6 | 0.61 (0.50–0.75) |
| MICA*002:01 | 190 (10.61%) | 376 (20.98%) | 3.82 × 10−17 | 8.05×10−16 | 0.45 (0.37–0.54) | 7.33 × 10−16 | 0.44 (0.37–0.54) |
| MICA*004 | 73 (4.08%) | 134 (7.48%) | 1.70 × 10−5 | 4.06×10−5 | 0.53 (0.40–0.71) | 4.98 × 10−5 | 0.54 (0.40–0.72) |
| MICA*012:01 | 55 (3.07%) | 144 (8.04%) | 1.49 × 10−10 | 1.92×10−9 | 0.37 (0.27–0.51) | 2.11 × 10−9 | 0.37 (0.27–0.51) |
| MICA*045 | 51 (2.85%) | 62 (3.46%) | 0.343 | 0.334 | 0.82 (0.57–1.19) | 0.360 | 0.83 (0.57–1.20) |
| MICA*033 | 39 (2.18%) | 2 (0.11%) | 1.17 × 10−8 | 6.46×10−5 | 19.36 (4.66–80.37) | 6.71 × 10−5 | 19.23 (4.63–79.86) |
| MICA*007:01 | 26 (1.45%) | 19 (1.06%) | 0.291 | 0.336 | 1.38 (0.76–2.51) | 0.360 | 1.38 (0.76–2.51) |
| MICA*018:01 | 5 (0.28%) | 4 (0.22%) | 0.738 | 0.738 | 1.25 (0.34–4.68) | 0.784 | 1.20 (0.32–4.51) |
| others | 62 (3.46%) | 97 (5.41%) | |||||
| MICA Allele | Estimated Frequency Trend Test | Logistic Regression | Logistic Regression Adjusted for Sex | ||||
|---|---|---|---|---|---|---|---|
| Synd+ (2N = 732) | Synd− (2N = 1058) | p Value | PFDR Value | OR (95% CI) | PFDR Value | OR (95% CI) | |
| MICA*019:01 | 343 (46.86%) | 422 (39.89%) | 8.72×10−5 | 0.001 | 1.68 (1.29–2.19) | 0.0017 | 1.69 (1.29–2.22) |
| MICA*008:01:01 | 119 (16.26%) | 213 (20.13%) | 0.030 | 0.077 | 0.75 (0.57–0.97) | 0.120 | 0.77 (0.59–1.01) |
| MICA*010:01 | 65 (8.88%) | 127 (12.00%) | 0.029 | 0.077 | 0.70 (0.50–0.97) | 0.120 | 0.71 (0.51–1.00) |
| MICA*002:01 | 72 (9.84%) | 118 (11.15%) | 0.352 | 0.401 | 0.86 (0.63–1.19) | 0.518 | 0.87 (0.63–1.21) |
| MICA*004 | 40 (5.46%) | 33 (3.12%) | 0.0196 | 0.071 | 1.81 (1.13–2.92) | 0.081 | 1.84 (1.12–3.02) |
| MICA*012:01 | 26 (3.55%) | 29 (2.74%) | 0.397 | 0.401 | 1.32 (0.76–2.28) | 0.518 | 1.27 (0.72–2.22) |
| MICA*045 | 14 (1.91%) | 37 (3.50%) | 0.063 | 0.105 | 0.54 (0.29–1.01) | 0.120 | 0.53 (0.28–1.00) |
| MICA*033 | 16 (2.19%) | 23 (2.17%) | 1 | 0.987 | 1.01 (0.53–1.90) | 0.841 | 1.07 (0.55–2.07) |
| MICA*007:01 | 7 (0.96%) | 19 (1.80%) | 0.199 | 0.247 | 0.52 (0.22–1.26) | 0.191 | 0.49 (0.20–1.19) |
| MICA*018:01 | 1 (0.14%) | 4 (0.38%) | 0.641 | 0.401 | 0.36 (0.04–3.23) | 0.540 | 0.45 (0.05–4.31) |
| others | 29 (3.96%) | 33 (3.12%) | |||||
| MICA Allele | Estimated Frequency Trend Test | Logistic Regression | Logistic Regression Adjusted for Sex | ||||
|---|---|---|---|---|---|---|---|
| B27+ (2N = 1568) | B27− (2N = 222) | p Value | PFDR Value | OR (95% CI) | PFDR Value | OR (95% CI) | |
| MICA*019:01 | 746 (47.58%) | 19 (8.56%) | 2.99 × 10−50 | 6.71 × 10−34 | 28.49 (16.72–48.53) | 1.45 × 10−33 | 28.79 (16.83–49.26) |
| MICA*008:01:01 | 274 (17.47%) | 58 (26.13%) | 0.001 | 0.002 | 0.55 (0.39–0.79) | 0.004 | 0.57 (0.40–0.81) |
| MICA*010:01 | 148 (9.44%) | 44 (19.82%) | 1.01 × 10−6 | 1.02 × 10−5 | 0.39 (0.26–0.58) | 1.78 × 10−5 | 0.39 (0.26–0.59) |
| MICA*002:01 | 138 (8.80%) | 52 (23.42%) | 5.47 × 10−12 | 7.18 × 10−10 | 0.27 (0.18–0.41) | 9.17 × 10−10 | 0.27 (0.18–0.41) |
| MICA*004 | 57 (3.64%) | 16 (7.21%) | 0.025 | 0.022 | 0.48 (0.27–0.86) | 0.018 | 0.47 (0.26–0.84) |
| MICA*012:01 | 47 (3.00%) | 8 (3.60%) | 0.724 | 0.688 | 0.82 (0.38–1.79) | 0.692 | 0.79 (0.36–1.73) |
| MICA*045 | 35 (2.23%) | 16 (7.21%) | 0.001 | 0.0003 | 0.29 (0.16–0.55) | 0.0003 | 0.29 (0.16–0.54) |
| MICA*033 | 39 (2.49%) | 0 (0.00%) | 0.001 | 0.999 | (0.00–Inf) | 0.999 | (0.00–Inf) |
| MICA*007:01 | 24 (1.53%) | 2 (0.90%) | 0.720 | 0.665 | 1.72 (0.40–7.38) | 0.692 | 1.68 (0.39–7.25) |
| MICA*018:01 | 4 (0.26%) | 1 (0.45%) | 0.964 | 0.688 | 0.56 (0.06–5.09) | 0.792 | 0.66 (0.07–6.09) |
| other | 56 (3.57%) | 6 (2.70%) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.-M.; Tan, K.-P.; Jan Wu, Y.-J.; Lin, J.-C.; Zheng, J.-W.; Yu, A.L.; Wu, J.-M.; Chen, J.-Y. MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese. J. Pers. Med. 2021, 11, 564. https://doi.org/10.3390/jpm11060564
Wang C-M, Tan K-P, Jan Wu Y-J, Lin J-C, Zheng J-W, Yu AL, Wu J-M, Chen J-Y. MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese. Journal of Personalized Medicine. 2021; 11(6):564. https://doi.org/10.3390/jpm11060564
Chicago/Turabian StyleWang, Chin-Man, Keng-Poo Tan, Yeong-Jian Jan Wu, Jing-Chi Lin, Jian-Wen Zheng, Alice L. Yu, Jian-Ming Wu, and Ji-Yih Chen. 2021. "MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese" Journal of Personalized Medicine 11, no. 6: 564. https://doi.org/10.3390/jpm11060564
APA StyleWang, C. -M., Tan, K. -P., Jan Wu, Y. -J., Lin, J. -C., Zheng, J. -W., Yu, A. L., Wu, J. -M., & Chen, J. -Y. (2021). MICA*019 Allele and Soluble MICA as Biomarkers for Ankylosing Spondylitis in Taiwanese. Journal of Personalized Medicine, 11(6), 564. https://doi.org/10.3390/jpm11060564