
Calcium-Sensing Receptor Polymorphisms at rs1801725 Are Associated with Increased Risk of Secondary Malignancies

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Clinical Laboratory Measurements
2.3. Genotyping and Quality Control Pipeline
2.4. Statistical Analysis
2.5. Statistical Power and Correction for Multiple Comparisons
3. Results
3.1. Characteristics of European and African Datasets
3.2. Allele Frequencies for rs1801725 and rs1801726
3.3. Circulating Calcium, Vitamin D, and PTH Levels in Individuals of European and African Ancestries
3.4. Association of Circulating Calcium, Vitamin D, and PTH with Primary Cancer Phenotypes
3.5. Association of CASR SNPS with Circulating Calcium, Vitamin D, and PTH
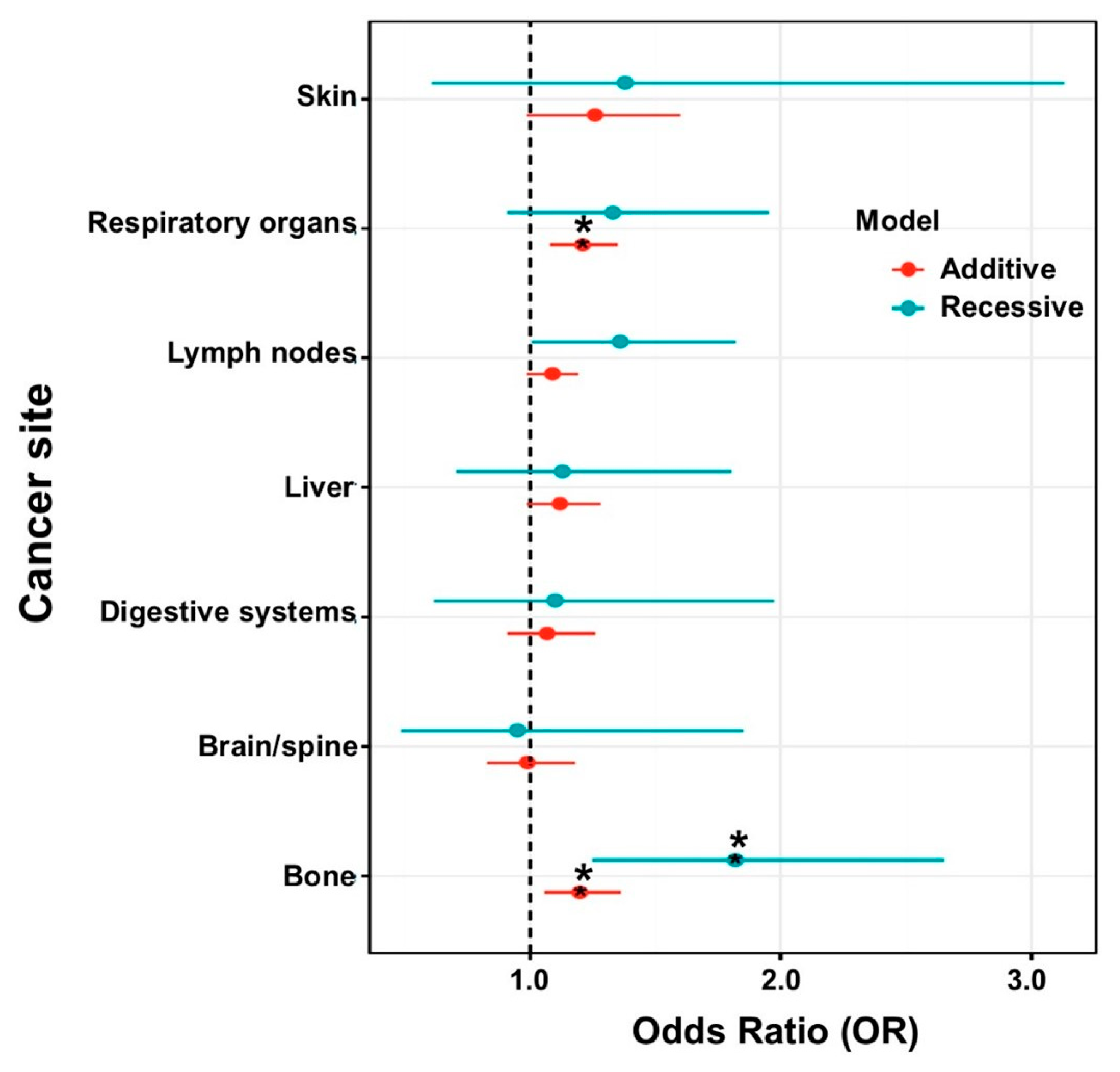
3.6. Association of CASR SNPs with Secondary Cancer Types
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aggarwal, A.; Prinz-Wohlgenannt, M.; Tennakoon, S.; Höbaus, J.; Boudot, C.; Mentaverri, R.; Brown, E.M.; Baumgartner-Parzer, S.; Kállay, E. The Calcium-Sensing Receptor: A Promising Target for Prevention of Colorectal Cancer. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 2158–2167. [Google Scholar] [CrossRef] [Green Version]
- Tennakoon, S.; Aggarwal, A.; Kállay, E. The Calcium-Sensing Receptor and the Hallmarks of Cancer. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Wysolmerski, J.J. Calcium-Sensing Receptor in Breast Physiology and Cancer. Front. Physiol. 2016, 7, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masvidal, L.; Iniesta, R.; García, M.; Casalà, C.; Lavarino, C.; Mora, J.; de Torres, C. Genetic Variants in the Promoter Region of the Calcium-Sensing Receptor Gene Are Associated with Its down-Regulation in Neuroblastic Tumors. Mol. Carcinog. 2017, 56, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Bedolla, M.A.; Carretero-Ortega, J.; Valadez-Sánchez, M.; Vázquez-Prado, J.; Reyes-Cruz, G. Chemotactic and Proangiogenic Role of Calcium Sensing Receptor Is Linked to Secretion of Multiple Cytokines and Growth Factors in Breast Cancer MDA-MB-231 Cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 166–182. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Hu, X.; Chakrabarty, S. Calcium Sensing Receptor Down-Regulates Malignant Cell Behavior and Promotes Chemosensitivity in Human Breast Cancer Cells. Cell Calcium 2009, 45, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Aslam, M.N.; Varani, J.; Chakrabarty, S. Induction of Calcium Sensing Receptor in Human Colon Cancer Cells by Calcium, Vitamin D and Aquamin: Promotion of a More Differentiated, Less Malignant and Indolent Phenotype. Mol. Carcinog. 2015, 54, 543–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tfelt-Hansen, J.; Brown, E.M. The Calcium-Sensing Receptor in Normal Physiology and Pathophysiology: A Review. Crit Rev. Clin. Lab. Sci. 2005, 42, 35–70. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Hediger, M.A.; Lytton, J.; Hebert, S.C. Cloning and Characterization of an Extracellular Ca(2+)− Sensing Receptor from Bovine Parathyroid. Nature 1993, 366, 575–580. [Google Scholar] [CrossRef]
- Ming, J.; Cronin, S.J.F.; Penninger, J.M. Targeting the RANKL/RANK/OPG Axis for Cancer Therapy. Front. Oncol. 2020, 10, 1283. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in Bone Modeling and Remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Guise, T.A.; Kozlow, W.M.; Heras-Herzig, A.; Padalecki, S.S.; Yin, J.J.; Chirgwin, J.M. Molecular Mechanisms of Breast Cancer Metastases to Bone. Clin. Breast Cancer 2005, 5, S46–S53. [Google Scholar] [CrossRef]
- Mundy, G.R. Metastasis to Bone: Causes, Consequences and Therapeutic Opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.L.; Chattopadhyay, N.; Kifor, O.; Yamaguchi, T.; Butters, R.R.; Brown, E.M. Extracellular Calcium-Sensing Receptor Expression and Its Potential Role in Regulating Parathyroid Hormone-Related Peptide Secretion in Human Breast Cancer Cell Lines. Endocrinology 2000, 141, 4357–4364. [Google Scholar] [CrossRef]
- Ralston, S.H.; Gallacher, S.J.; Patel, U.; Campbell, J.; Boyle, I.T. Cancer-Associated Hypercalcemia: Morbidity and Mortality. Clinical Experience in 126 Treated Patients. Ann. Intern. Med. 1990, 112, 499–504. [Google Scholar] [CrossRef]
- Stewart, A.F. Hypercalcemia Associated with Cancer. N. Engl. J. Med. 2005, 352, 373–379. [Google Scholar] [CrossRef]
- Almquist, M.; Anagnostaki, L.; Bondeson, L.; Bondeson, A.-G.; Borgquist, S.; Landberg, G.; Malina, J.; Malm, J.; Manjer, J. Serum Calcium and Tumour Aggressiveness in Breast Cancer: A Prospective Study of 7847 Women. Eur. J. Cancer Prev. 2009, 18, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Thaw, S.S.H.; Sahmoun, A.; Schwartz, G.G. Serum Calcium, Tumor Size, and Hormone Receptor Status in Women with Untreated Breast Cancer. Cancer Biol. Ther. 2012, 13, 467–471. [Google Scholar] [CrossRef]
- Hendy, G.N.; D’Souza-Li, L.; Yang, B.; Canaff, L.; Cole, D.E. Mutations of the Calcium-Sensing Receptor (CASR) in Familial Hypocalciuric Hypercalcemia, Neonatal Severe Hyperparathyroidism, and Autosomal Dominant Hypocalcemia. Hum. Mutat. 2000, 16, 281–296. [Google Scholar] [CrossRef]
- Pidasheva, S.; D’Souza-Li, L.; Canaff, L.; Cole, D.E.C.; Hendy, G.N. CASRdb: Calcium-Sensing Receptor Locus-Specific Database for Mutations Causing Familial (Benign) Hypocalciuric Hypercalcemia, Neonatal Severe Hyperparathyroidism, and Autosomal Dominant Hypocalcemia. Hum. Mutat. 2004, 24, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Thakker, R.V. Diseases Associated with the Extracellular Calcium-Sensing Receptor. Cell Calcium 2004, 35, 275–282. [Google Scholar] [CrossRef]
- Pratt, J.H.; Ambrosius, W.T.; Wagner, M.A.; Maharry, K. Molecular Variations in the Calcium-Sensing Receptor in Relation to Sodium Balance and Presence of Hypertension in Blacks and Whites. Am. J. Hypertens. 2000, 13, 654–658. [Google Scholar] [CrossRef] [Green Version]
- Muddana, V.; Lamb, J.; Greer, J.B.; Elinoff, B.; Hawes, R.H.; Cotton, P.B.; Anderson, M.A.; Brand, R.E.; Slivka, A.; Whitcomb, D.C. Association between Calcium Sensing Receptor Gene Polymorphisms and Chronic Pancreatitis in a US Population: Role of Serine Protease Inhibitor Kazal 1type and Alcohol. World J. Gastroenterol. 2008, 14, 4486. [Google Scholar] [CrossRef]
- Lorch, G.; Viatchenko-Karpinski, S.; Ho, H.-T.; Dirksen, W.P.; Toribio, R.E.; Foley, J.; Györke, S.; Rosol, T.J. The Calcium-Sensing Receptor Is Necessary for the Rapid Development of Hypercalcemia in Human Lung Squamous Cell Carcinoma. Neoplasia 2011, 13, 428–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Widatalla, S.E.; Whalen, D.S.; Ochieng, J.; Sakwe, A.M. Association of Calcium Sensing Receptor Polymorphisms at Rs1801725 with Circulating Calcium in Breast Cancer Patients. BMC Cancer 2017, 17, 511. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.G.; John, E.M.; Rowland, G.; Ingles, S.A. Prostate Cancer in African-American Men and Polymorphism in the Calcium-Sensing Receptor. Cancer Biol. Ther. 2010, 9, 994–999. [Google Scholar] [CrossRef] [Green Version]
- März, W.; Seelhorst, U.; Wellnitz, B.; Tiran, B.; Obermayer-Pietsch, B.; Renner, W.; Boehm, B.O.; Ritz, E.; Hoffmann, M.M. Alanine to Serine Polymorphism at Position 986 of the Calcium-Sensing Receptor Associated with Coronary Heart Disease, Myocardial Infarction, All-Cause, and Cardiovascular Mortality. J. Clin. Endocrinol. Metab. 2007, 92, 2363–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scillitani, A.; Guarnieri, V.; De Geronimo, S.; Muscarella, L.A.; Battista, C.; D’Agruma, L.; Bertoldo, F.; Florio, C.; Minisola, S.; Hendy, G.N.; et al. Blood Ionized Calcium Is Associated with Clustered Polymorphisms in the Carboxyl-Terminal Tail of the Calcium-Sensing Receptor. J. Clin. Endocrinol. Metab. 2004, 89, 5634–5638. [Google Scholar] [CrossRef] [Green Version]
- Cole, D.E.; Vieth, R.; Trang, H.M.; Wong, B.Y.; Hendy, G.N.; Rubin, L.A. Association between Total Serum Calcium and the A986S Polymorphism of the Calcium-Sensing Receptor Gene. Mol. Genet. Metab 2001, 72, 168–174. [Google Scholar] [CrossRef]
- Vezzoli, G.; Scillitani, A.; Corbetta, S.; Terranegra, A.; Dogliotti, E.; Guarnieri, V.; Arcidiacono, T.; Macrina, L.; Mingione, A.; Brasacchio, C.; et al. Risk of Nephrolithiasis in Primary Hyperparathyroidism Is Associated with Two Polymorphisms of the Calcium-Sensing Receptor Gene. J. Nephrol. 2015, 28, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Roden, D.M.; Pulley, J.M.; Basford, M.A.; Bernard, G.R.; Clayton, E.W.; Balser, J.R.; Masys, D.R. Development of a Large-Scale de-Identified DNA Biobank to Enable Personalized Medicine. Clin. Pharmacol. Ther. 2008, 84, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Denny, J.C.; Bastarache, L.; Ritchie, M.D.; Carroll, R.J.; Zink, R.; Mosley, J.D.; Field, J.R.; Pulley, J.M.; Ramirez, A.H.; Bowton, E.; et al. Systematic Comparison of Phenome-Wide Association Study of Electronic Medical Record Data and Genome-Wide Association Study Data. Nat. Biotechnol. 2013, 31, 1102–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, J.K.; Sealock, J.M.; Straub, P.; Lee, Y.H.; Hucks, D.; Actkins, K.; Faucon, A.; Feng, Y.-C.A.; Ge, T.; Goleva, S.B.; et al. Clinical Laboratory Test-Wide Association Scan of Polygenic Scores Identifies Biomarkers of Complex Disease. Genome Med. 2021, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Bien, S.A.; Wojcik, G.L.; Zubair, N.; Gignoux, C.R.; Martin, A.R.; Kocarnik, J.M.; Martin, L.W.; Buyske, S.; Haessler, J.; Walker, R.W.; et al. Strategies for Enriching Variant Coverage in Candidate Disease Loci on a Multiethnic Genotyping Array. PLoS ONE 2016, 11, e0167758. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G.; Qiu, Y.; Inouye, M. FlashPCA2: Principal Component Analysis of Biobank-Scale Genotype Datasets. Bioinformatics 2017, 33, 2776–2778. [Google Scholar] [CrossRef]
- Kumar, R.; Tebben, P.J.; Thompson, J.R. Vitamin D and the Kidney. Arch. Biochem. Biophys. 2012, 523, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Silva, B.C.; Bilezikian, J.P. Parathyroid Hormone: Anabolic and Catabolic Actions on the Skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Li, T.; Liao, D.; Zhu, Z.; Dong, Y. Quantitative Assessment of the Clinical Susceptibility of Calcium-Sensing Receptor Polymorphisms in Cancer Patients. Cancer Manag. Res. 2018, 10, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Kapur, K.; Johnson, T.; Beckmann, N.D.; Sehmi, J.; Tanaka, T.; Kutalik, Z.; Styrkarsdottir, U.; Zhang, W.; Marek, D.; Gudbjartsson, D.F.; et al. Genome-Wide Meta-Analysis for Serum Calcium Identifies Significantly Associated SNPs near the Calcium-Sensing Receptor (CASR) Gene. PLoS Genet. 2010, 6, e1001035. [Google Scholar] [CrossRef] [Green Version]
- Abid, H.A.; Inoue, A.; Gorvin, C.M. Heterogeneity of G-Protein Activation by the Calcium-Sensing Receptor. J. Mol. Endocrinol. 2021. [Google Scholar] [CrossRef]
- Saidak, Z.; Boudot, C.; Abdoune, R.; Petit, L.; Brazier, M.; Mentaverri, R.; Kamel, S. Extracellular Calcium Promotes the Migration of Breast Cancer Cells through the Activation of the Calcium Sensing Receptor. Exp. Cell Res. 2009, 315, 2072–2080. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Wang, H.; Canaff, L.; Hendy, G.N.; Appelman, H.; Varani, J. Calcium Sensing Receptor in Human Colon Carcinoma: Interaction with Ca(2+) and 1,25-Dihydroxyvitamin D(3). Cancer Res. 2005, 65, 493–498. [Google Scholar] [PubMed]
- Whitfield, J.F. Calcium, Calcium-Sensing Receptor and Colon Cancer. Cancer Lett. 2009, 275, 9–16. [Google Scholar] [CrossRef]
- Iamartino, L.; Elajnaf, T.; Kallay, E.; Schepelmann, M. Calcium-Sensing Receptor in Colorectal Inflammation and Cancer: Current Insights and Future Perspectives. World J. Gastroenterol. 2018, 24, 4119–4131. [Google Scholar] [CrossRef]
- Li, F.; Tian, Z. The Liver Works as a School to Educate Regulatory Immune Cells. Cell Mol. Immunol. 2013, 10, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.-S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The Calcium-Sensing Receptor Regulates the NLRP3 Inflammasome through Ca2+ and CAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Pierer, M.; Raulien, N.; Quandt, D.; Meusch, U.; Rothe, K.; Schubert, K.; Schöneberg, T.; Schaefer, M.; Krügel, U.; et al. Extracellular Ca2+ Is a Danger Signal Activating the NLRP3 Inflammasome through G Protein-Coupled Calcium Sensing Receptors. Nat. Commun. 2012, 3, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moossavi, M.; Parsamanesh, N.; Bahrami, A.; Atkin, S.L.; Sahebkar, A. Role of the NLRP3 Inflammasome in Cancer. Mol. Cancer 2018, 17, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Huang, C.-F.; Li, Y.-C.; Deng, W.-W.; Mao, L.; Wu, L.; Zhang, W.-F.; Zhang, L.; Sun, Z.-J. Blockage of the NLRP3 Inflammasome by MCC950 Improves Anti-Tumor Immune Responses in Head and Neck Squamous Cell Carcinoma. Cell Mol. Life Sci. 2018, 75, 2045–2058. [Google Scholar] [CrossRef]
- Cheng, S.X.; Lightfoot, Y.L.; Yang, T.; Zadeh, M.; Tang, L.; Sahay, B.; Wang, G.P.; Owen, J.L.; Mohamadzadeh, M. Epithelial CaSR Deficiency Alters Intestinal Integrity and Promotes Proinflammatory Immune Responses. FEBS Lett. 2014, 588, 4158–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piranavan, P.; Li, Y.; Brown, E.; Kemp, E.H.; Trivedi, N. Immune Checkpoint Inhibitor-Induced Hypoparathyroidism Associated With Calcium-Sensing Receptor-Activating Autoantibodies. J. Clin. Endocrinol. Metab. 2019, 104, 550–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupi, I.; Brancatella, A.; Cetani, F.; Latrofa, F.; Kemp, E.H.; Marcocci, C. Activating Antibodies to The Calcium-Sensing Receptor in Immunotherapy-Induced Hypoparathyroidism. J. Clin. Endocrinol. Metab. 2020, 105, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Chamberlin, M.; Kemp, E.H.; Weetman, A.P.; Khadka, B.; Brown, E.M. Immunosuppressive Therapy of Autoimmune Hypoparathyroidism in a Patient with Activating Autoantibodies against the Calcium-Sensing Receptor. Clin. Endocrinol. 2019, 90, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Aranow, C. Vitamin D and the Immune System. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagzag, J.; Hu, M.I.; Fisher, S.B.; Perrier, N.D. Hypercalcemia and Cancer: Differential Diagnosis and Treatment. CA Cancer J. Clin. 2018, 68, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Demographics | European Dataset | African Dataset | p |
|---|---|---|---|
| N | 53680 | 10777 | |
| Average Age (SD) | 48.74 (22.03) | 37.98 (21.57) | <0.001 |
| Average BMI (SD) | 27.75 (7.25) | 29.04 (8.65) | <0.001 |
| Sex (%) | <0.001 | ||
| Female | 30801 (57.4) | 6767 (62.8) | |
| Male | 22879 (42.6) | 4010 (37.2) | |
| rs1801725 Alleles (%) | <0.001 | ||
| GG | 39939 (74.4) | 9976 (92.6) | |
| GT | 12736 (23.7) | 773 (7.2) | |
| TT | 1005 (1.9) | 20 (0.2) | |
| rs1801726 Alleles (%) | <0.001 | ||
| CC | 49733 (92.7) | 7487 (69.5) | |
| CG | 3858 (7.2) | 2972 (27.6) | |
| GG | 87 (0.2) | 309 (2.9) | |
| EHR-Reported Race (%) | <0.001 | ||
| White | 52461 (97.7) | 285 (2.6) | |
| Black | 59 (0.1) | 10313 (95.7) | |
| Calcium (mg/dL) | |||
| N | 51176 | 9878 | |
| Median [IQR] | 9.30 (9.05, 9.60) | 9.40 (9.10, 9.60) | <0.001 |
| Ionized Calcium (mg/dL) | |||
| N | 13899 | 2336 | |
| Median [IQR] | 4.52 (4.32, 4.73) | 4.57 (4.35, 4.80) | <0.001 |
| Vitamin D (ng/mL) | |||
| N | 18885 | 3341 | |
| Median [IQR] | 31 (24, 39) | 23 (17, 30.50) | <0.001 |
| Parathyroid Hormone (pg/mL) | |||
| N | 6937 | 1430 | |
| Median [IQR] | 56 (36, 91) | 82 (50, 160) | <0.001 |
| rs1801725 | rs1801726 | |||
|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) | p | |
| Calcium | 1.15 (1.13–1.16) | 8.40 × 10−73* | 0.98 (0.96–1.01) | 0.16 |
| Ionized Calcium | 1.10 (1.07–1.14) | 1.43 × 10−9* | 1.03 (0.97–1.09) | 0.28 |
| Vitamin D | 0.99 (0.96–1.02) | 0.50 | 0.98 (0.94–1.03) | 0.51 |
| Parathyroid Hormone | 1.00 (0.95–1.05) | 0.98 | 1.03 (0.94–1.12) | 0.57 |
| Primary Cancer | 1.02 (0.97–1.08) | 0.40 | 1.00 (0.91–1.09) | 0.95 |
| Secondary Malignancy | 1.08 (1.01–1.15) | 0.03 | 1.00 (0.88–1.14) | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Actkins, K.V.; Beasley, H.K.; Faucon, A.B.; Davis, L.K.; Sakwe, A.M. Calcium-Sensing Receptor Polymorphisms at rs1801725 Are Associated with Increased Risk of Secondary Malignancies. J. Pers. Med. 2021, 11, 642. https://doi.org/10.3390/jpm11070642
Actkins KV, Beasley HK, Faucon AB, Davis LK, Sakwe AM. Calcium-Sensing Receptor Polymorphisms at rs1801725 Are Associated with Increased Risk of Secondary Malignancies. Journal of Personalized Medicine. 2021; 11(7):642. https://doi.org/10.3390/jpm11070642
Chicago/Turabian StyleActkins, Ky’Era V., Heather K. Beasley, Annika B. Faucon, Lea K. Davis, and Amos M. Sakwe. 2021. "Calcium-Sensing Receptor Polymorphisms at rs1801725 Are Associated with Increased Risk of Secondary Malignancies" Journal of Personalized Medicine 11, no. 7: 642. https://doi.org/10.3390/jpm11070642