
C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Databases
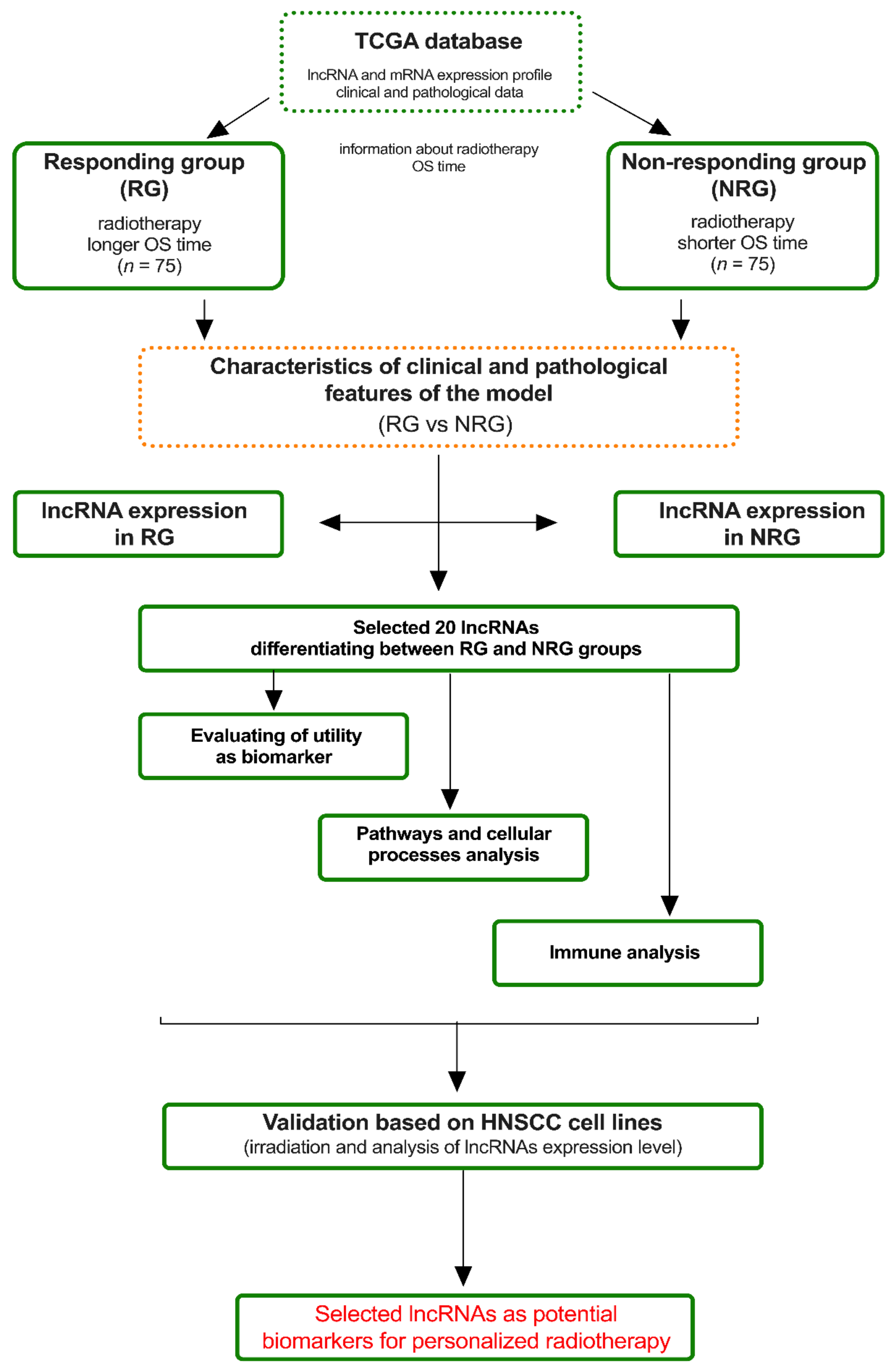
2.2. Obtaining the Model and Its Characterization
2.3. Pathways and Cellular Processes Analysis
2.4. Immune Analysis
2.5. Irradiation of HNSCC Cell Lines
2.6. lncRNA Expression Analysis
2.7. Statistical Methods
3. Results
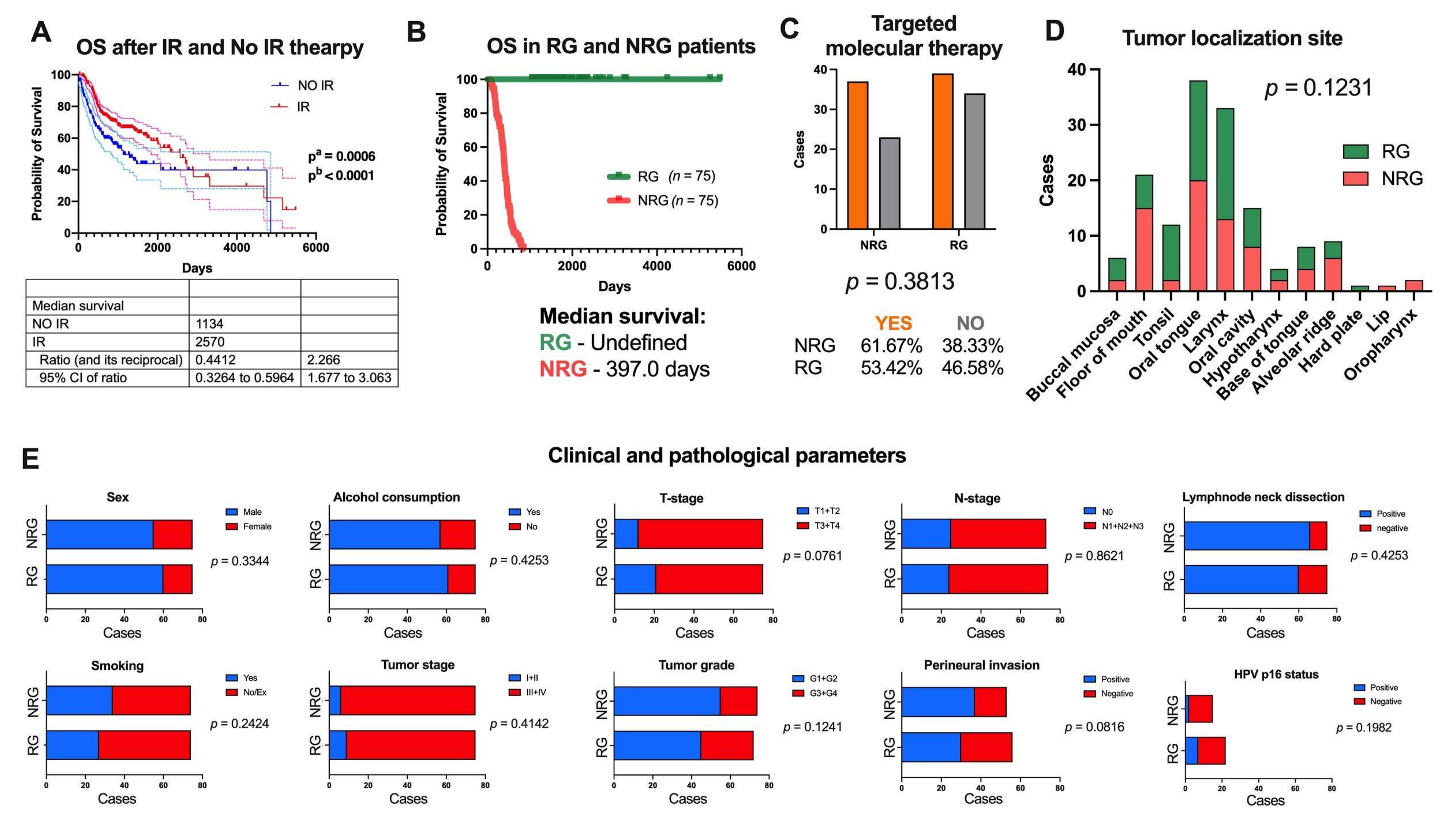
3.1. Obtaining an In Silico Experimental Model Based on TCGA Data
3.2. lncRNA Expression Profile Differs between RG and NRG Patients
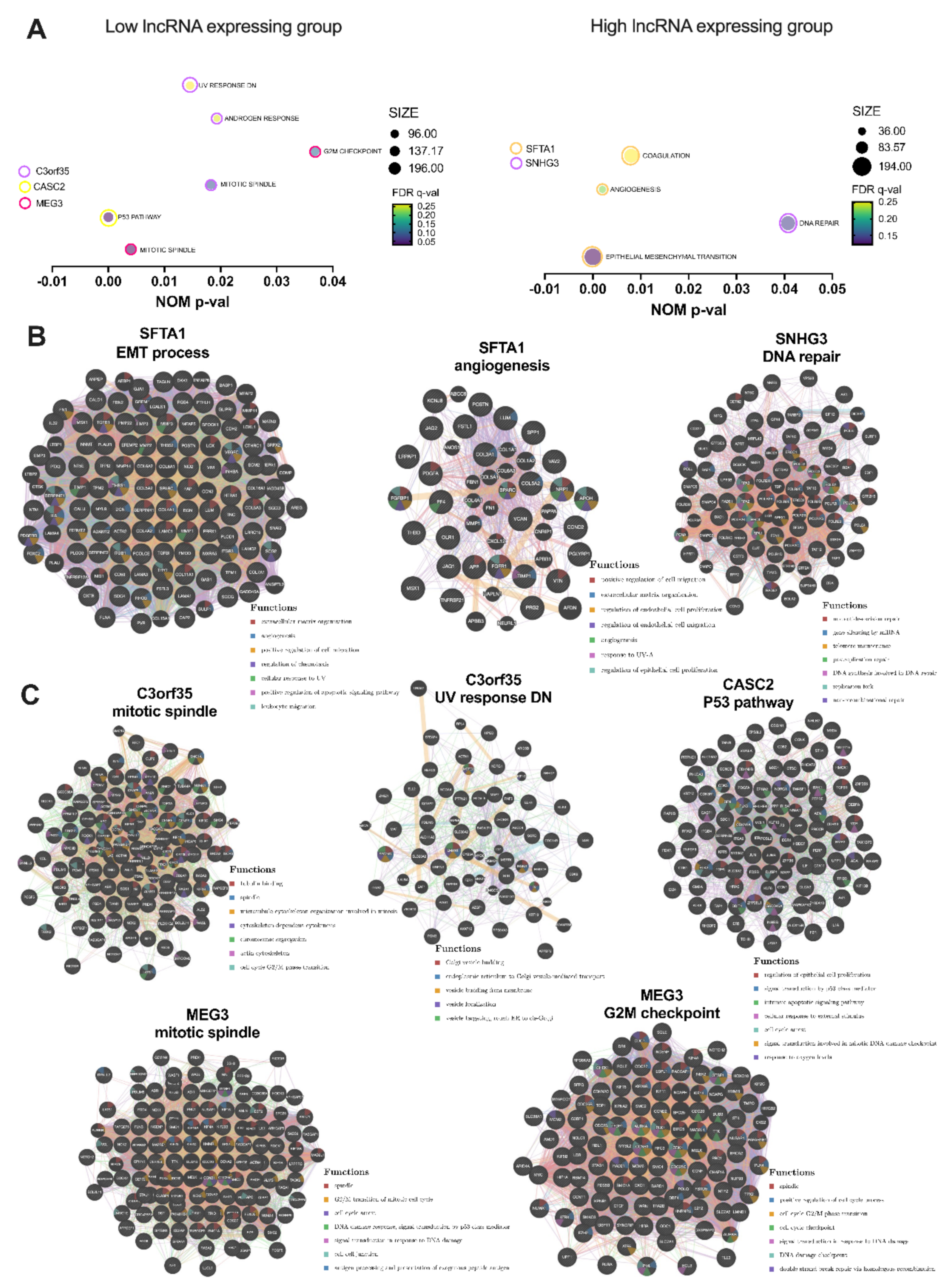
3.3. Selected lncRNAs Are Connected with Pathways and Cellular Processes Important for Response to Irradiation
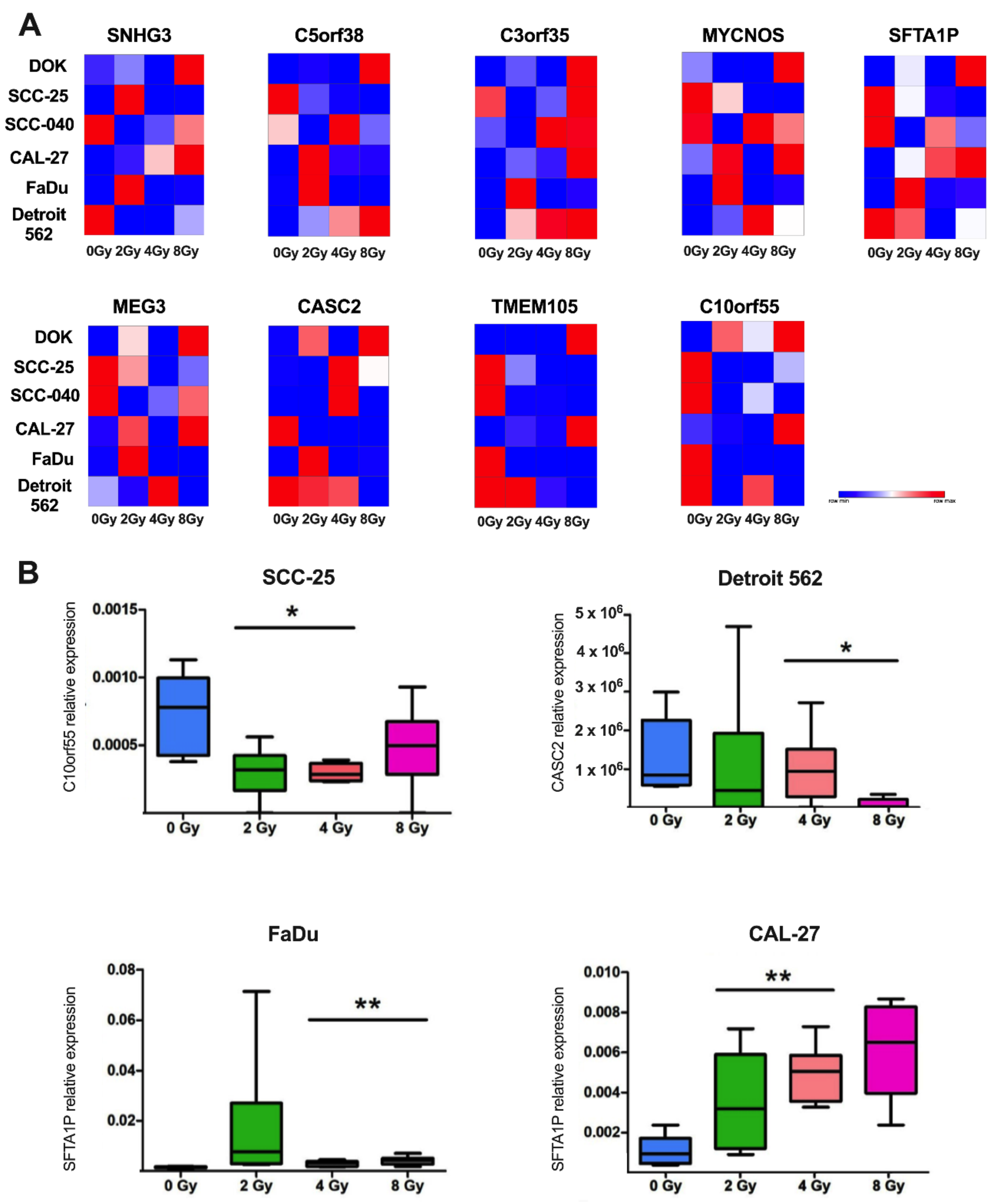
3.4. Expression Level of lncRNAs Selected Based on TCGA Data WasChanged after Ionization Radiation in Cell Line Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mandal, R.; Şenbabaoğlu, Y.; Desrichard, A.; Havel, J.J.; Dalin, M.G.; Riaz, N.; Lee, K.W.; Ganly, I.; Hakimi, A.A.; Chan, T.A.; et al. The head and neck cancer immune landscape and its immunotherapeutic implications. JCI Insight 2016, 1, e89829. [Google Scholar] [CrossRef] [PubMed]
- Conway, D.I.; Hashibe, M.; Boffetta, P.; Wunsch-Filho, V.; Muscat, J.; La Vecchia, C.; Winn, D.M. Enhancing epidemiologic research on head and neck cancer: INHANCE—The international head and neck cancer epidemiology consortium. Oral Oncol. 2009, 45, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Liu, J.; Zhang, L.; Dong, S.; Zhang, J.; Liu, Y.; Zhou, H.; Dong, W. Identification of Potential Biomarkers and Survival Analysis for Head and Neck Squamous Cell Carcinoma Using Bioinformatics Strategy: A Study Based on TCGA and GEO Datasets. BioMed Res. Int. 2019, 2019, 7376034. [Google Scholar] [CrossRef]
- Jethwa, A.R.; Khariwala, S.S. Tobacco-related carcinogenesis in head and neck cancer. Cancer Metastasis Rev. 2017, 36, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Vokes, E.E.; Agrawal, N.; Seiwert, T.Y. HPV-Associated Head and Neck Cancer. J. Natl. Cancer Inst. 2015, 107, djv344. [Google Scholar] [CrossRef]
- Specenier, P.; Vermorken, J.B. Optimizing treatments for recurrent or metastatic head and neck squamous cell carcinoma. Expert Rev. Anticancer Ther. 2018, 18, 901–915. [Google Scholar] [CrossRef]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Cramer, J.D.; Burtness, B.; Le, Q.T.; Ferris, R.L. The changing therapeutic landscape of head and neck cancer. Nat. Rev. Clin. Oncol. 2019, 16, 669–683. [Google Scholar] [CrossRef]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef] [PubMed]
- O’Rorke, M.A.; Ellison, M.V.; Murray, L.J.; Moran, M.; James, J.; Anderson, L.A. Human papillomavirus related head and neck cancer survival: A systematic review and meta-analysis. Oral Oncol. 2012, 48, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef]
- Zhou, C.; Parsons, J.L. The radiobiology of HPV-positive and HPV-negative head and neck squamous cell carcinoma. Expert Rev. Mol. Med. 2020, 22, e3. [Google Scholar] [CrossRef]
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Baskar, R.; Dai, J.; Wenlong, N.; Yeo, R.; Yeoh, K.W. Biological response of cancer cells to radiation treatment. Front. Mol. Biosci. 2014, 1, 24. [Google Scholar] [CrossRef]
- Feller, G.; Khammissa, R.A.G.; Nemutandani, M.S.; Feller, L. Biological consequences of cancer radiotherapy in the context of oral squamous cell carcinoma. Head Face Med. 2021, 17, 35. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Qian, H. Biological effects of radiation on cancer cells. Mil. Med. Res. 2018, 5, 20. [Google Scholar] [CrossRef]
- Han, P.B.; Ji, X.J.; Zhang, M.; Gao, L.Y. Upregulation of lncRNA LINC00473 promotes radioresistance of HNSCC cells through activating Wnt/β-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7305–7313. [Google Scholar] [CrossRef]
- Chen, L.L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef]
- Kozłowska, J.; Kolenda, T.; Poter, P.; Sobocińska, J.; Guglas, K.; Stasiak, M.; Bliźniak, R.; Teresiak, A.; Lamperska, K. Long Intergenic Non-Coding RNAs in HNSCC: From “Junk DNA” to Important Prognostic Factor. Cancers 2021, 13, 2949. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Smith, K.N.; Miller, S.C.; Varani, G.; Calabrese, J.M.; Magnuson, T. Multimodal Long Noncoding RNA Interaction Networks: Control Panels for Cell Fate Specification. Genetics 2019, 213, 1093–1110. [Google Scholar] [CrossRef]
- Wang, W.; Min, L.; Qiu, X.; Wu, X.; Liu, C.; Ma, J.; Zhang, D.; Zhu, L. Biological Function of Long Non-coding RNA (LncRNA) Xist. Front. Cell Dev. Biol. 2021, 9, 645647. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2018, 48, 812–830.e14. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The Gene MANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Lindell Jonsson, E.; Erngren, I.; Engskog, M.; Haglöf, J.; Arvidsson, T.; Hedeland, M.; Petterson, C.; Laurell, G.; Nestor, M. Exploring Radiation Response in Two Head and Neck Squamous Carcinoma Cell Lines Through Metabolic Profiling. Front. Oncol. 2019, 9, 825. [Google Scholar] [CrossRef]
- Guglas, K.; Kolenda, T.; Stasiak, M.; Kopczyńska, M.; Teresiak, A.; Ibbs, M.; Bliźniak, R.; Lamperska, K. YRNAs: New Insights and Potential Novel Approach in Head and Neck Squamous Cell Carcinoma. Cells 2020, 9, 1281. [Google Scholar] [CrossRef]
- Chen, G.; Sun, J.; Xie, M.; Yu, S.; Tang, Q.; Chen, L. PLAU Promotes Cell Proliferation and Epithelial-Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Front. Genet. 2021, 12, 651882. [Google Scholar] [CrossRef]
- Shi, Y.; Ren, J.; Zhuang, Z.; Zhang, W.; Wang, Z.; Liu, Y.; Li, J.; Liang, T.; He, R.; Wang, K. Comprehensive Analysis of a ceRNA Network Identifies lncRC3orf35 Associated with Poor Prognosis in Osteosarcoma. BioMed Res. Int. 2020, 2020, 3178037. [Google Scholar] [CrossRef]
- Available online: http://www.oligoarchitect.com/AlternatePrimers.jsp (accessed on 15 May 2022).
- Tao, L.; Tian, P.; Yang, L.; Guo, X. lncRNA CASC2 Enhances 131I Sensitivity in Papillary Thyroid Cancer by Sponging miR-155. BioMed Res. Int. 2020, 2020, 7183629. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Huang, Y.; Chen, T.; Wang, W.; Yang, S.; Ye, Z.; Xi, X. LncRNA MEG3 inhibits the progression of prostate cancer by modulating miR-9-5p/QKI-5 axis. J. Cell Mol. Med. 2019, 23, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ou, Z.; Lei, Y.; Chen, L.; Su, Q.; Zhang, K. LncRNA MYCNOS facilitates proliferation and invasion in hepatocellular carcinoma by regulating miR-340. Hum. Cell 2020, 33, 148–158. [Google Scholar] [CrossRef]
- Du, D.; Shen, X.; Zhang, Y.; Yin, L.; Pu, Y.; Liang, G. Expression of long non-coding RNA SFTA1P and its function in non-small cell lung cancer. Pathol. Res. Pract. 2020, 216, 153049. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Pan, M.; Li, Y.; Lu, T.; Wang, Z.; Liu, C.; Hu, G. RNA N6-methyladenosine reader IGF2BP2 promotes lymphatic metastasis and epithelial-mesenchymal transition of head and neck squamous carcinoma cells via stabilizing slug mRNA in an m6A-dependent manner. J. Exp. Clin. Cancer Res. 2022, 41, 6. [Google Scholar] [CrossRef]
- Guglas, K.; Kolenda, T.; Teresiak, A.; Kopczyńska, M.; Łasińska, I.; Mackiewicz, J.; Mackiewicz, A.; Lamperska, K. lncRNA Expression after Irradiation and Chemoexposure of HNSCC Cell Lines. Noncoding RNA 2018, 4, 33. [Google Scholar] [CrossRef]
- Lei, F.; Lei, T.; Huang, Y.; Yang, M.; Liao, M.; Huang, W. Radio-Susceptibility of Nasopharyngeal Carcinoma: Focus on Epstein- Barr Virus, MicroRNAs, Long Non-Coding RNAs and Circular RNAs. Curr. Mol. Pharmacol. 2020, 13, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Wu, X.; Li, Y. Identification and validation of potential long non-coding RNA biomarkers in predicting survival of patients with head and neck squamous cell carcinoma. Oncol. Lett. 2019, 17, 5642–5652. [Google Scholar] [CrossRef]
- Walker, C.J.; Mrózek, K.; Ozer, H.G.; Nicolet, D.; Kohlschmidt, J.; Papaioannou, D.; Genutis, L.K.; Bill, M.; Powell, B.L.; Uy, G.L.; et al. Gene expression signature predicts relapse in adult patients with cytogenetically normal acute myeloid leukemia. Blood Adv. 2021, 5, 1474–1482. [Google Scholar] [CrossRef]
- Zhang, D.; Yin, H.; Bauer, T.L.; Rogers, M.P.; Velotta, J.B.; Morgan, C.T.; Du, W.; Xu, P.; Qian, X. Development of a novel miR-3648-related gene signature as a prognostic biomarker in esophageal adenocarcinoma. Ann. Transl. Med. 2021, 9, 1702. [Google Scholar] [CrossRef]
- O’Leary, V.B.; Ovsepian, S.V.; Carrascosa, L.G.; Buske, F.A.; Radulovic, V.; Niyazi, M.; Moertl, S.; Trau, M.; Atkinson, M.J.; Anastasov, N. PARTICLE, a Triplex-Forming Long ncRNA, Regulates Locus-Specific Methylation in Response to Low-Dose Irradiation. Cell Rep. 2015, 11, 474–485. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/gene/255082 (accessed on 15 June 2022).
- Zhang, C.; Cao, W.; Wang, J.; Liu, J.; Liu, J.; Wu, H.; Li, S.; Zhang, C. A prognostic long noncoding RNA-associated competing endogenous RNA network in head and neck squamous cell carcinoma. PeerJ 2020, 8, e9701. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Yu, Y.; Qi, Y.; Wu, K.; Liu, D.; Yang, Y.; Zhang, C.; Zhao, S. Long Non-coding RNA CASC2 Enhances the Antitumor Activity of Cisplatin Through Suppressing the Akt Pathway by Inhibition of miR-181a in Esophageal Squamous Cell Carcinoma Cells. Front. Oncol. 2019, 9, 350. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, A.; Takahashi, H.; Patel, A.A.; Osman, A.A.; Myers, J.N. Targeting the DNA Damage Response in OSCC with TP53 Mutations. J. Dent. Res. 2018, 97, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Nathan, N.; Berdah, L.; Delestrain, C.; Sileo, C.; Clement, A. Interstitial lung diseases in children. Presse Med. 2020, 49, 103909. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Finch-Edmondson, M.; Leong, K.W.; Zhang, X.; Lin, Q.X.X.; Lee, Y.; Ng, W.T.; Guo, H.; Wan, Y.; Sudol, M.; et al. LncRNA SFTA1P mediates positive feedback regulation of the Hippo-YAP/TAZ signaling pathway in non-small cell lung cancer. Cell Death Discov. 2021, 7, 369. [Google Scholar] [CrossRef]
- Jiang, W.; Song, Y.; Zhong, Z.; Gao, J.; Meng, X. Ferroptosis-Related Long Non-Coding RNA Signature Contributes to the Prediction of Prognosis Outcomes in Head and Neck Squamous Cell Carcinomas. Front. Genet. 2021, 12, 785839. [Google Scholar] [CrossRef]
- Li, L.; Yin, J.Y.; He, F.Z.; Huang, M.S.; Zhu, T.; Gao, Y.F.; Chen, Y.X.; Zhou, D.B.; Chen, X.; Sun, L.Q.; et al. Long noncoding RNA SFTA1P promoted apoptosis and increased cisplatin chemosensitivity via regulating the hnRNP-U-GADD45A axis in lung squamous cell carcinoma. Oncotarget 2017, 8, 97476–97489. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, M.; Zhou, C.; Wang, W.; Yang, H.; Ye, W. The role of epithelial-mesenchymal transition in regulating radioresistance. Crit. Rev. Oncol. Hematol. 2020, 150, 102961. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Dou, C.; Liu, H.; Xu, B.; Hu, B.; Kuang, L.; Yao, J.; Zhao, Y.; Yu, S.; Li, Y.; et al. Comprehensive analysis: Necroptosis-related lncRNAs can effectively predict the prognosis of glioma patients. Front. Oncol. 2022, 12, 929233. [Google Scholar] [CrossRef] [PubMed]
- Zimta, A.A.; Tigu, A.B.; Braicu, C.; Stefan, C.; Ionescu, C.; Berindan-Neagoe, I. An Emerging Class of Long Non-coding RNA With Oncogenic Role Arises From the snoRNA Host Genes. Front. Oncol. 2020, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhong, Z.; Shao, Y.; Yi, Y. Prognostic Value of MEG3 and Its Correlation with Immune Infiltrates in Gliomas. Front. Genet. 2021, 12, 679097. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Hao, C.; Lu, Y.; Duan, X.; Liang, R.; Gao, G.; Zhang, T. MEG3 modulates TIGIT expression and CD4 + T cell activation through absorbing miR-23a. Mol. Cell Biochem. 2019, 454, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, Y.; Nakatani, K.; Nakagawara, A. De novo evolved gene product NCYM in the pathogenesis and clinical outcome of human neuroblastomas and other cancers. Jpn. J. Clin. Oncol. 2020, 50, 839–846. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA | Forward Primer (5′-3′) | Right Primer (5′-3′) | Ref: |
|---|---|---|---|
| C10orf55 | ATTCGGGAGGAGGCTTCATCA | TGAGAACTAGATACGAACAGGGT | [29] |
| C3orf35 | AAGAGGTTATTGTGCGCCCG | ATTAGCCCGCCTTCCTCTGT | [30] |
| C5orf38 | CTGCTGCCTGTTACTAAT | CAATGACGAGTGTTAAGTT | [31] |
| CASC2 | GGCTCACAAAGCCTAGGTTA | CCTTGGATATTTCCAAGAGC | [32] |
| MEG3 | CTGCCCATCTACACCTCACG | CTCTCCGCCGTCTGCGCTAGGGGCT | [33] |
| MYCNOS | TCCGACAGCTCAAACACAGAC | CCAGCTTTGCAGCCTTCTC | [34] |
| SFTA1P | CATTCCAGGTGGGCTTTCA | TCCCTTGTTTGGCTTACTCG | [35] |
| SNHG3 | AATCCAGTCTCATTCAGTAA | GCGTCCTAATCATTCAATC | [31] |
| TMEM105 | TGGCAGCAGGGATAACAG | TGAGCAACAGAGCAAGACT | [31] |
| GAPDH | CCACTCCTCCACCTTTGACG | CCACCACCCTGTTGCTGTAG | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paszkowska, A.; Kolenda, T.; Guglas, K.; Kozłowska-Masłoń, J.; Podralska, M.; Teresiak, A.; Bliźniak, R.; Dzikiewicz-Krawczyk, A.; Lamperska, K. C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers. J. Pers. Med. 2022, 12, 1696. https://doi.org/10.3390/jpm12101696
Paszkowska A, Kolenda T, Guglas K, Kozłowska-Masłoń J, Podralska M, Teresiak A, Bliźniak R, Dzikiewicz-Krawczyk A, Lamperska K. C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers. Journal of Personalized Medicine. 2022; 12(10):1696. https://doi.org/10.3390/jpm12101696
Chicago/Turabian StylePaszkowska, Anna, Tomasz Kolenda, Kacper Guglas, Joanna Kozłowska-Masłoń, Marta Podralska, Anna Teresiak, Renata Bliźniak, Agnieszka Dzikiewicz-Krawczyk, and Katarzyna Lamperska. 2022. "C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers" Journal of Personalized Medicine 12, no. 10: 1696. https://doi.org/10.3390/jpm12101696
APA StylePaszkowska, A., Kolenda, T., Guglas, K., Kozłowska-Masłoń, J., Podralska, M., Teresiak, A., Bliźniak, R., Dzikiewicz-Krawczyk, A., & Lamperska, K. (2022). C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers. Journal of Personalized Medicine, 12(10), 1696. https://doi.org/10.3390/jpm12101696