
Establishment of Adenomyosis Organoids as a Preclinical Model to Study Infertility

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Derivation and Culture of Organoids from Human Endometrial Tissue Samples
2.3. Differentiation of Endometrial Organoids to Secretory and Gestational Phases
2.4. Immunohistochemical (IHC) and Immunofluorescence (IF) Characterization
2.5. Gene Expression Analysis
2.6. ELISA
2.7. Chromosomal Stability
2.8. Statistical Analysis
3. Results
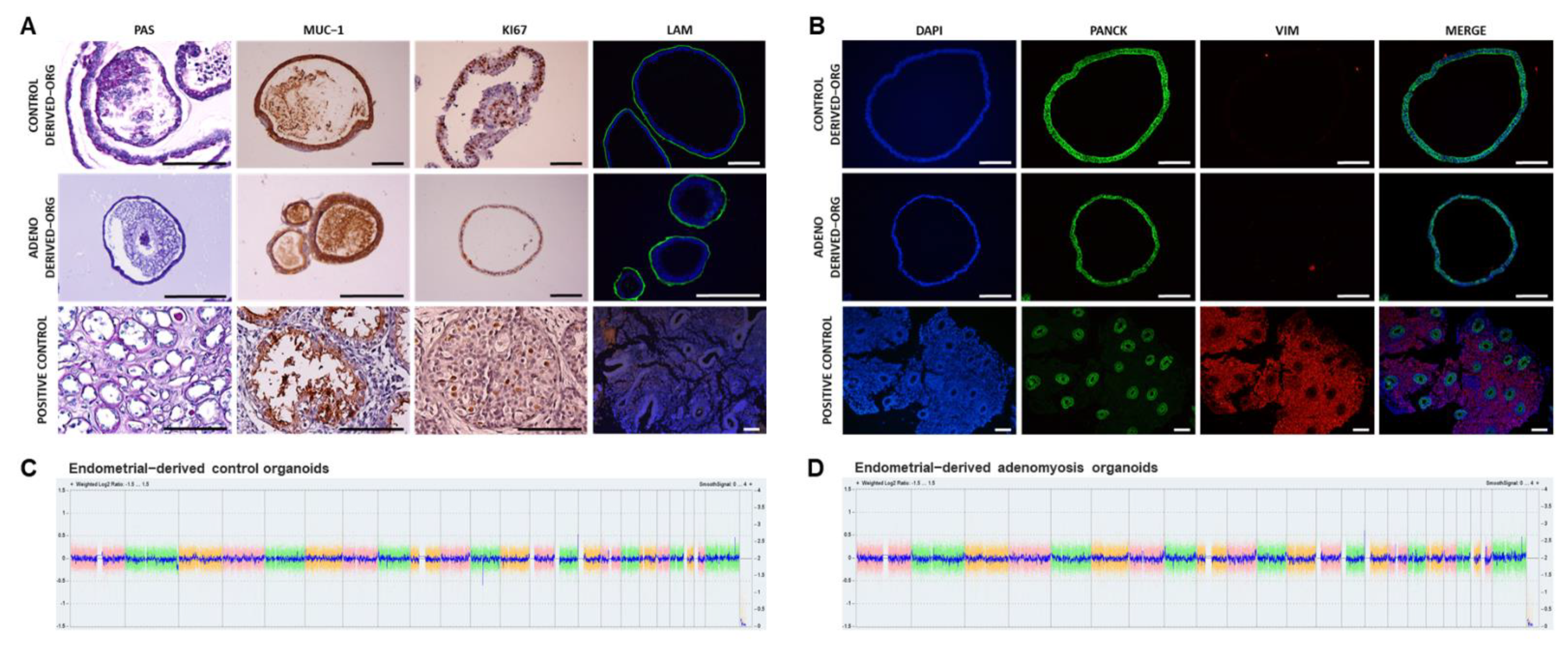
3.1. Human Endometrial Organoids Can Be Derived from Adenomyosis Patients and Recapitulate Endometrial Gland Biology In Vivo
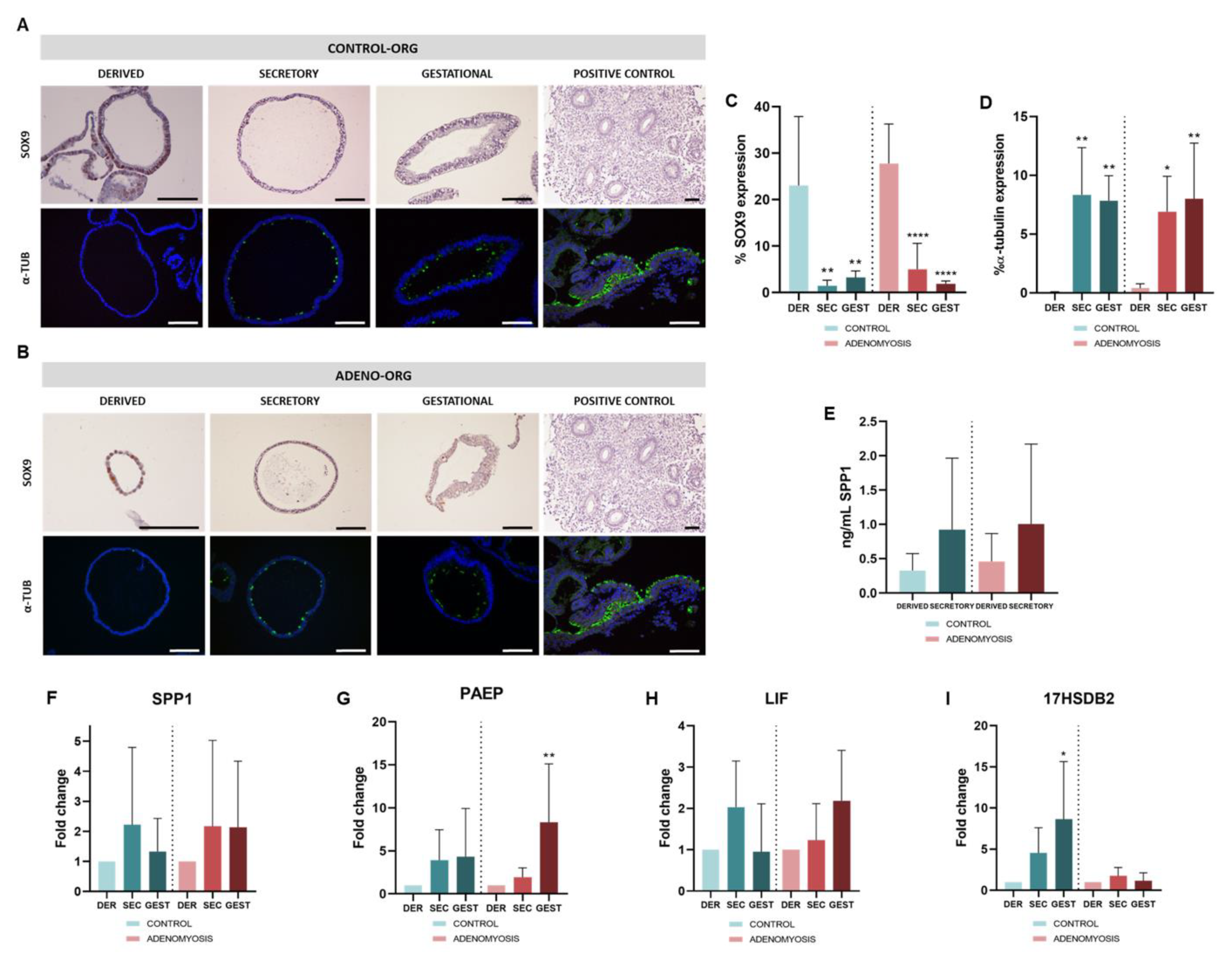
3.2. Differentiation to Secretory and Gestational Phases of Human Adenomyosis-Derived Organoids in Response to Hormonal Treatments
3.3. Human Adenomyosis-Derived Organoids Maintain Chromosomal Stability after Differentiation
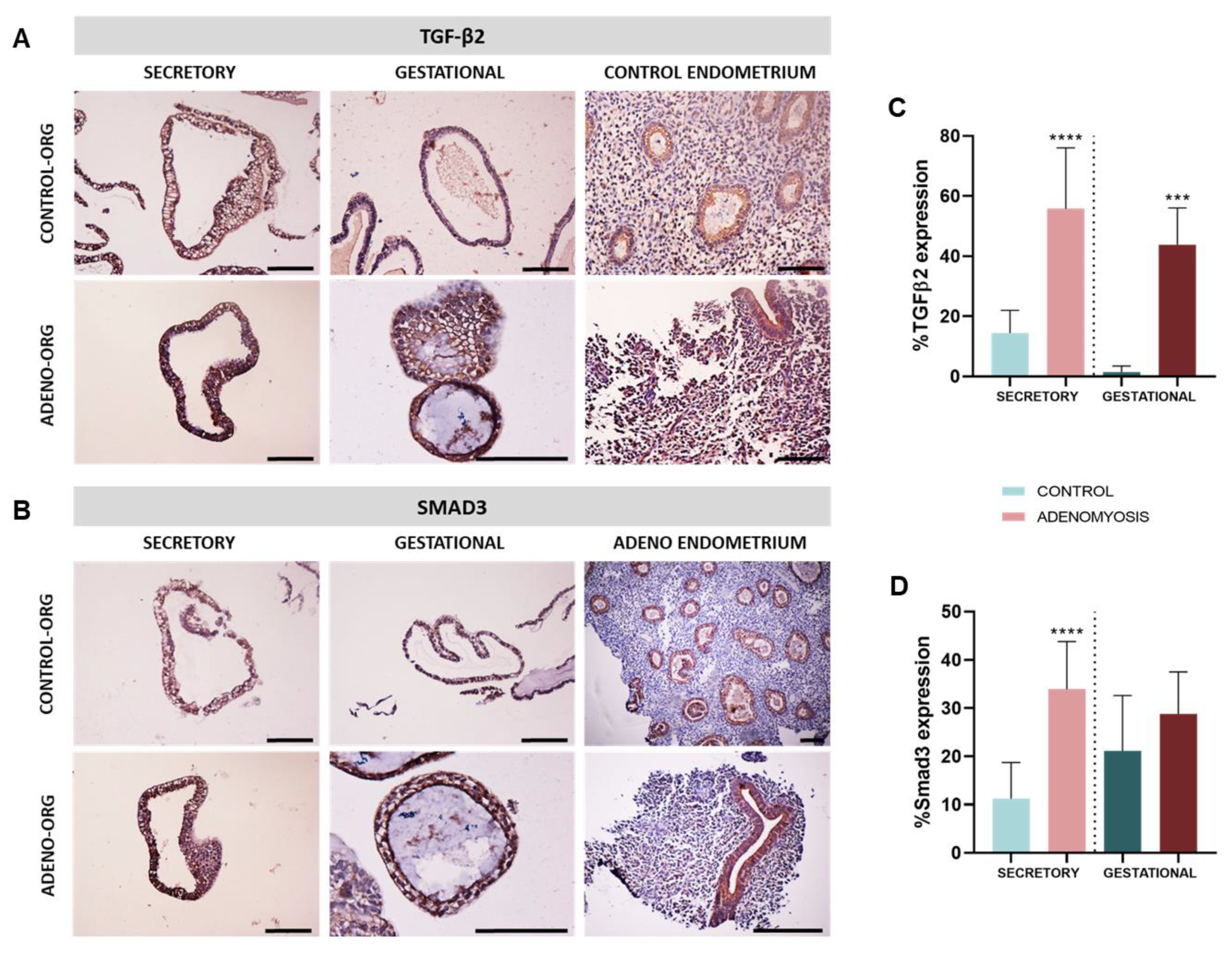
3.4. Human Adenomyosis Secretory and Gestational Organoids Recapitulate Disease-Specific Traits
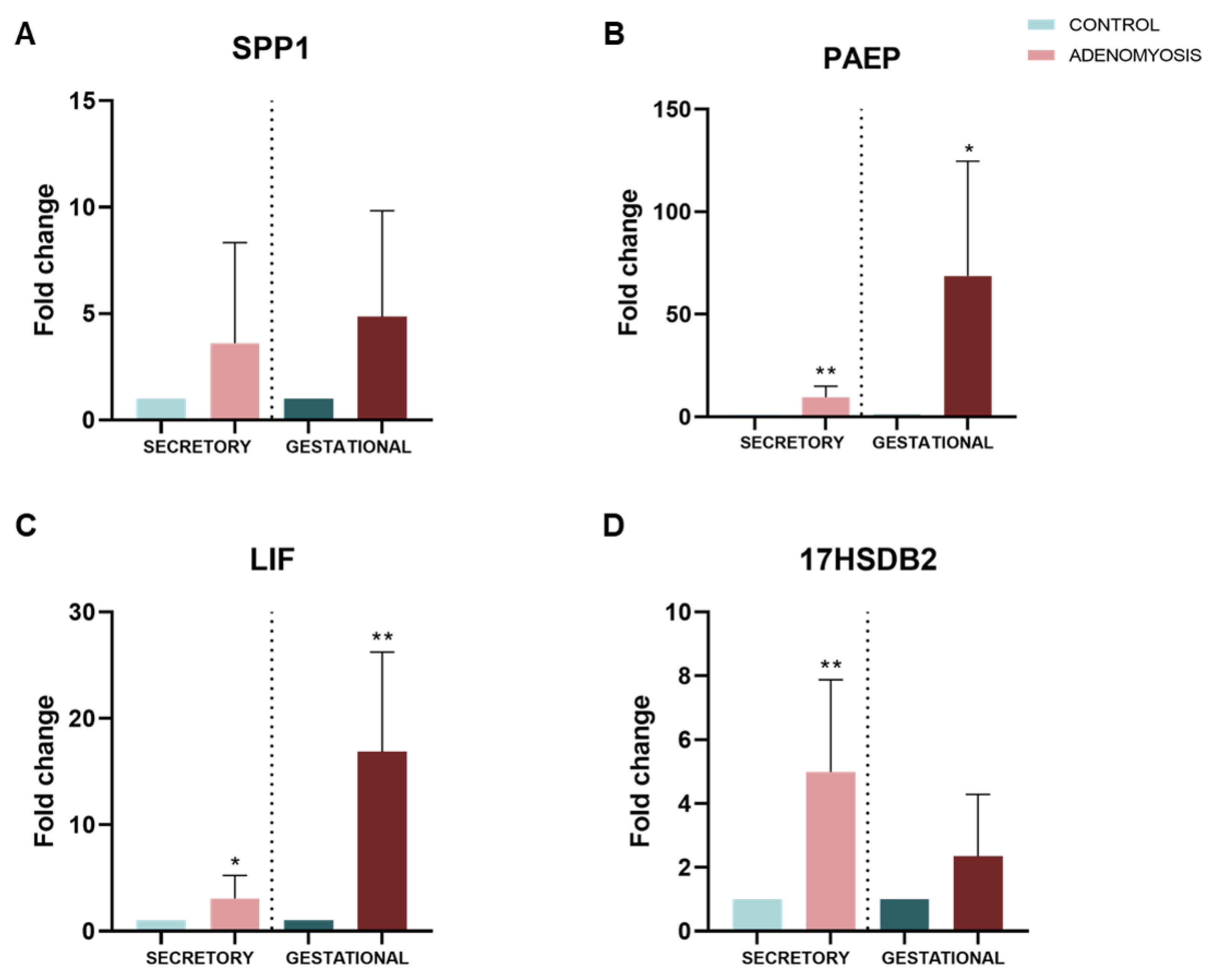
3.5. Dysregulation of Secretory and Gestational Biomarkers in Human Adenomyosis Organoids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munro, M.G. Uterine polyps, adenomyosis, leiomyomas, and endometrial receptivity. Fertil. Steril. 2019, 111, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Vercellini, P.; Bonfanti, I.; Berlanda, N. Adenomyosis and infertility: Is there a causal link? Expert Rev. Endocrinol. Metab. 2019, 14, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Stratopoulou, C.A.; Dolmans, M.-M. Uterine Adenomyosis: From Disease Pathogenesis to a New Medical Approach Using GnRH Antagonists. Int. J. Environ. Res. Public Health 2021, 18, 9941. [Google Scholar] [CrossRef]
- Stratopoulou, C.A.; Donnez, J.; Dolmans, M.-M. Origin and Pathogenic Mechanisms of Uterine Adenomyosis: What Is Known So Far. Reprod. Sci. 2021, 28, 2087–2097. [Google Scholar] [CrossRef]
- García-Solares, J.; Donnez, J.; Donnez, O.; Dolmans, M.-M. Pathogenesis of uterine adenomyosis: Invagination or metaplasia? Fertil. Steril. 2018, 109, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Leyendecker, G.; Wildt, L. A new concept of endometriosis and adenomyosis: Tissue injury and repair (TIAR). Horm. Mol. Biol. Clin. Investig. 2011, 5, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Li, H.-Y.; Huang, C.-H.; Twu, N.-F.; Yen, M.-S.; Wang, P.-H.; Chou, T.-Y.; Liu, Y.-N.; Chao, K.-C.; Yang, M.-H. Oestrogen-induced epithelial-mesenchymal transition of endometrial epithelial cells contributes to the development of adenomyosis. J. Pathol. 2010, 222, 261–270. [Google Scholar] [CrossRef]
- Batt, R.E.; Yeh, J. Müllerianosis. Reprod. Sci. 2013, 20, 1030–1037. [Google Scholar] [CrossRef]
- Signorile, P.G.; Baldi, F.; Bussani, R.; D’Armiento, M.; De Falco, M.; Baldi, A. Ectopic endometrium in human foetuses is a common event and sustains the theory of müllerianosis in the pathogenesis of endometriosis, a disease that predisposes to cancer. J. Exp. Clin. Cancer Res. 2009, 28, 49. [Google Scholar] [CrossRef] [Green Version]
- Gargett, C.E.; Schwab, K.E.; Brosens, J.J.; Puttemans, P.; Benagiano, G.; Brosens, I. Potential role of endometrial stem/progenitor cells in the pathogenesis of early-onset endometriosis. Mol. Hum. Reprod. 2014, 20, 591–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargett, C.E.; Schwab, K.E.; Deane, J.A. Endometrial stem/progenitor cells: The first 10 years. Hum. Reprod. Update 2015, 22, 137–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, J.A. Metastatic or Embolic Endometriosis, due to the Menstrual Dissemination of Endometrial Tissue into the Venous Circulation. Am. J. Pathol. 1927, 3, 93–110.43. [Google Scholar]
- Morassutto, C.; Monasta, L.; Ricci, G.; Barbone, F.; Ronfani, L. Incidence and Estimated Prevalence of Endometriosis and Adenomyosis in Northeast Italy: A Data Linkage Study. PLoS ONE 2016, 11, e0154227. [Google Scholar] [CrossRef] [Green Version]
- Yu, O.; Schulze-Rath, R.; Grafton, J.; Hansen, K.; Scholes, D.; Reed, S.D. Adenomyosis incidence, prevalence and treatment: United States population-based study 2006–2015. Am. J. Obstet. Gynecol. 2020, 223, 94.e1–94.e10. [Google Scholar] [CrossRef]
- Chapron, C.; Vannuccini, S.; Santulli, P.; Abrão, M.S.; Carmona, F.; Fraser, I.S.; Gordts, S.; Guo, S.-W.; Just, P.-A.; Noël, J.-C.; et al. Diagnosing adenomyosis: An integrated clinical and imaging approach. Hum. Reprod. Update 2020, 26, 392–411. [Google Scholar] [CrossRef]
- Tellum, T.; Nygaard, S.; Lieng, M. Noninvasive Diagnosis of Adenomyosis: A Structured Review and Meta-analysis of Diagnostic Accuracy in Imaging. J. Minim. Invasive Gynecol. 2020, 27, 408–418.e3. [Google Scholar] [CrossRef]
- Upson, K.; Missmer, S.A. Epidemiology of Adenomyosis. Semin. Reprod. Med. 2020, 38, 089–107. [Google Scholar] [CrossRef]
- Peric, H.; Fraser, I.S. The symptomatology of adenomyosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 547–555. [Google Scholar] [CrossRef]
- Puente, J.M.; Fabris, A.; Patel, J.; Patel, A.; Cerrillo, M.; Requena, A.; Garcia-Velasco, J.A. Adenomyosis in infertile women: Prevalence and the role of 3D ultrasound as a marker of severity of the disease. Reprod. Biol. Endocrinol. 2016, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Vercellini, P.; Consonni, D.; Dridi, D.; Bracco, B.; Frattaruolo, M.P.; Somigliana, E. Uterine adenomyosis and in vitro fertilization outcome: A systematic review and meta-analysis. Hum. Reprod. 2014, 29, 964–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, H.; Kishi, H.; Kitade, M.; Asai-Sato, M.; Tanaka, A.; Murakami, T.; Minegishi, T.; Sugino, N. Clinical outcomes of infertility treatment for women with adenomyosis in Japan. Reprod. Med. Biol. 2017, 16, 276–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younes, G.; Tulandi, T. Effects of adenomyosis on in vitro fertilization treatment outcomes: A meta-analysis. Fertil. Steril. 2017, 108, 483–490.e3. [Google Scholar] [CrossRef] [Green Version]
- Carrarelli, P.; Yen, C.-F.; Funghi, L.; Arcuri, F.; Tosti, C.; Bifulco, G.; Luddi, A.; Lee, C.-L.; Petraglia, F. Expression of Inflammatory and Neurogenic Mediators in Adenomyosis. Reprod. Sci. 2017, 24, 369–375. [Google Scholar] [CrossRef]
- Benagiano, G.; Brosens, I.; Habiba, M. Structural and molecular features of the endomyometrium in endometriosis and adenomyosis. Hum. Reprod. Update 2014, 20, 386–402. [Google Scholar] [CrossRef]
- Ota, H.; Tanaka, T. Stromal vascularization in the endometrium during adenomyosis. Microsc. Res. Tech. 2003, 60, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Vannuccini, S.; Petraglia, F.; Giudice, L.C. Adenomyosis: Mechanisms and Pathogenesis. Semin. Reprod. Med. 2020, 38, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, M.; Santulli, P.; Jeljeli, M.; Vannuccini, S.; Marcellin, L.; Doridot, L.; Petraglia, F.; Batteux, F.; Chapron, C. Immunological changes associated with adenomyosis: A systematic review. Hum. Reprod. Update 2021, 27, 108–129. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Wu, M.Y.; Chen, C.D.; Chen, M.J.; Yang, Y.S.; Ho, H.N. Altered apoptosis and proliferation in endometrial stromal cells of women with adenomyosis. Hum. Reprod. 2007, 22, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Peng, G.Q.; Ban, D.Y.; Zhang, C.; Zhang, X.Q.; Li, Y.P. High-Expression of Neuropilin 1 Correlates to Estrogen-Induced Epithelial-Mesenchymal Transition of Endometrial Cells in Adenomyosis. Reprod. Sci. 2020, 27, 395–403. [Google Scholar] [CrossRef]
- Moustakas, A.; Heldin, C.-H. Signaling networks guiding epithelial?mesenchymal transitions during embryogenesis and cancer progression. Cancer Sci. 2007, 98, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-Y.; Ku, B.J.; Kim, T.H.; Il Ahn, J.; Ahn, J.Y.; Yang, W.S.; Lim, J.M.; Taketo, M.M.; Shin, J.-H.; Jeong, J.-W. β-catenin activates TGF-β-induced epithelial–mesenchymal transition in adenomyosis. Exp. Mol. Med. 2020, 52, 1754–1765. [Google Scholar] [CrossRef]
- Liu, X.; Shen, M.; Qi, Q.; Zhang, H.; Guo, S.-W. Corroborating evidence for platelet-induced epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the development of adenomyosis. Hum. Reprod. 2016, 31, 734–749. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Li, D.; Yuan, M.; Li, Q.; Zhang, L.; Wang, G. Different macrophages equally induce EMT in endometria of adenomyosis and normal. Reproduction 2017, 154, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Letamendia, A.; Labbé, E.; Attisano, L. Transcriptional regulation by Smads: Crosstalk between the TGF-beta and Wnt pathways. J. Bone Joint Surg. Am. 2001, 83 (Suppl. 1), S31–S39. [Google Scholar] [CrossRef] [PubMed]
- Kriseman, M.; Monsivais, D.; Agno, J.; Masand, R.P.; Creighton, C.J.; Matzuk, M.M. Uterine double-conditional inactivation of Smad2 and Smad3 in mice causes endometrial dysregulation, infertility, and uterine cancer. Proc. Natl. Acad. Sci. 2019, 116, 3873–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Li, S.; Deng, J.; Yang, S.; Xiang, Z.; Guo, H.; Xi, H.; Sang, M.; Zhang, W. Aspirin inhibits endometrial fibrosis by suppressing the TGF-β1-Smad2/Smad3 pathway in intrauterine adhesions. Int. J. Mol. Med. 2020, 45, 1351–1360. [Google Scholar] [CrossRef]
- Li, J.; Dong, X.; Yang, P.; Yang, S.; Hu, D.; Zhang, H.; Sui, C. Activation of Uterine Smad3 Pathway Is Crucial for Embryo Implantation. Curr. Med. Sci. 2019, 39, 997–1002. [Google Scholar] [CrossRef]
- Lessey, B.A. Adhesion molecules and implantation. J. Reprod. Immunol. 2002, 55, 101–112. [Google Scholar] [CrossRef]
- White, F.J.; Burghardt, R.C.; Hu, J.; Joyce, M.M.; Spencer, T.E.; Johnson, G.A. Secreted phosphoprotein 1 (osteopontin) is expressed by stromal macrophages in cyclic and pregnant endometrium of mice, but is induced by estrogen in luminal epithelium during conceptus attachment for implantation. Reproduction 2006, 132, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, U.; Kuhn, C.; Mylonas, I.; Schulze, S.; Friese, K.; Mayr, D.; Speer, R.; Briese, V.; Richter, D.-U.; Haase, M.; et al. Development and characterization of monoclonal antibodies for the immunohistochemical detection of glycodelin A in decidual, endometrial and gynaecological tumour tissues. Histopathology 2006, 48, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Stavreus-Evers, A.; Mandelin, E.; Koistinen, R.; Aghajnova, L.; Hovatta, O.; Seppälä, M. Glycodelin is present in pinopodes of receptive-phase human endometrium and is associated with down-regulation of progesterone receptor B. Fertil. Steril. 2006, 85, 1803–1811. [Google Scholar] [CrossRef] [PubMed]
- Focarelli, R.; Luddi, A.; De Leo, V.; Capaldo, A.; Stendardi, A.; Pavone, V.; Benincasa, L.; Belmonte, G.; Petraglia, F.; Piomboni, P. Dysregulation of GdA Expression in Endometrium of Women With Endometriosis: Implication for Endometrial Receptivity. Reprod. Sci. 2018, 25, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Tee, M.K.; Vigne, J.-L.; Yu, J.; Taylor, R.N. Natural and recombinant human glycodelin activate a proapoptotic gene cascade in monocyte cells. J. Leukoc. Biol. 2008, 83, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-L.; Lam, K.K.W.; Vijayan, M.; Koistinen, H.; Seppala, M.; Ng, E.H.Y.; Yeung, W.S.B.; Chiu, P.C.N. The Pleiotropic Effect of Glycodelin-A in Early Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Prašnikar, E.; Kunej, T.; Knez, J.; Repnik, K.; Potočnik, U.; Kovačič, B. Determining the molecular background of endometrial receptivity in adenomyosis. Biomolecules 2020, 10, 1311. [Google Scholar] [CrossRef]
- Yoo, I.; Chae, S.; Han, J.; Lee, S.; Kim, H.J.; Ka, H. Leukemia inhibitory factor and its receptor: Expression and regulation in the porcine endometrium throughout the estrous cycle and pregnancy. Asian-Australasian J. Anim. Sci. 2019, 32, 192. [Google Scholar] [CrossRef]
- Bulun, S.E.; Cheng, Y.H.; Yin, P.; Imir, G.; Utsunomiya, H.; Attar, E.; Innes, J.; Julie Kim, J. Progesterone resistance in endometriosis: Link to failure to metabolize estradiol. Mol. Cell. Endocrinol. 2006, 248, 94–103. [Google Scholar] [CrossRef]
- Drolet, R.; Simard, M.; Plante, J.; Laberge, P.; Tremblay, Y. Human type 2 17beta-hydroxysteroid dehydrogenase mRNA and protein distribution in placental villi at mid and term pregnancy. Reprod. Biol. Endocrinol. 2007, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Kitawaki, J.; Koshiba, H.; Ishihara, H.; Kusuki, I.; Tsukamoto, K.; Honjo, H. Progesterone induction of 17β-hydroxysteroid dehydrogenase type 2 during the secretory phase occurs in the endometrium of estrogen-dependent benign diseases but not in normal endometrium. J. Clin. Endocrinol. Metab. 2000, 85, 3292–3296. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Jiang, R.; Cheng, X.; Zhang, Q.; Hu, Y.; Zhang, H.; Cao, Y.; Zhang, M.; Wang, J.; Ding, L.; et al. Decreased expression of NR4A nuclear receptors in adenomyosis impairs endometrial decidualization. Mol. Hum. Reprod. 2016, 22, 655–668. [Google Scholar] [CrossRef] [Green Version]
- Kissler, S.; Hamscho, N.; Zangos, S.; Wiegratz, I.; Schlichter, S.; Menzel, C.; Doebert, N.; Gruenwald, F.; Vogl, T.; Gaetje, R.; et al. Uterotubal transport disorder in adenomyosis and endometriosis—a cause for infertility. BJOG An Int. J. Obstet. Gynaecol. 2006, 113, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.P.; Kayisili, U.; Taylor, H.S. HOXA10 expression is decreased in endometrium of women with adenomyosis. Fertil. Steril. 2011, 95, 1133–1136. [Google Scholar] [CrossRef]
- Campo, S.; Campo, V.; Benagiano, G. Infertility and Adenomyosis. Obstet. Gynecol. Int. 2012, 2012, 786132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudoma, I. The evaluation of pinopode formation in patients with adenomyosis. Fertil. Steril. 2002, 77, S27. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [Green Version]
- Maru, Y.; Tanaka, N.; Itami, M.; Hippo, Y. Efficient use of patient-derived organoids as a preclinical model for gynecologic tumors. Gynecol. Oncol. 2019, 154, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Deane, J.A.; Cousins, F.L.; Gargett, C.E. Endometrial organoids: In vitro models for endometrial research and personalized medicine. Biol. Reprod. 2017, 97, 781–783. [Google Scholar] [CrossRef]
- Heidari-Khoei, H.; Esfandiari, F.; Hajari, M.A.; Ghorbaninejad, Z.; Piryaei, A.; Baharvand, H. Organoid technology in female reproductive biomedicine. Reprod. Biol. Endocrinol. 2020, 18, 64. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Hughes, J.; Cindrova-Davies, T.; Gomez, M.J.; Farrell, L.; Hollinshead, M.; Marsh, S.G.E.; Brosens, J.J.; Critchley, H.O.; et al. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nat. Cell Biol. 2017, 19, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Boretto, M.; Cox, B.; Noben, M.; Hendriks, N.; Fassbender, A.; Roose, H.; Amant, F.; Timmerman, D.; Tomassetti, C.; Vanhie, A.; et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development 2017, 144, 1775–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boretto, M.; Maenhoudt, N.; Luo, X.; Hennes, A.; Boeckx, B.; Bui, B.; Heremans, R.; Perneel, L.; Kobayashi, H.; Van Zundert, I.; et al. Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening. Nat. Cell Biol. 2019, 21, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Francés-Herrero, E.; Juárez-Barber, E.; Campo, H.; López-Martínez, S.; de Miguel-Gómez, L.; Faus, A.; Pellicer, A.; Ferrero, H.; Cervelló, I. Improved models of human endometrial organoids based on hydrogels from decellularized endometrium. J. Pers. Med. 2021, 11, 504. [Google Scholar] [CrossRef]
- Alzamil, L.; Nikolakopoulou, K.; Turco, M.Y. Organoid systems to study the human female reproductive tract and pregnancy. Cell Death Differ. 2021, 28, 35–51. [Google Scholar] [CrossRef]
- Ojosnegros, S.; Seriola, A.; Godeau, A.L.; Veiga, A. Embryo implantation in the laboratory: An update on current techniques. Hum. Reprod. Update 2021, 27, 501–530. [Google Scholar] [CrossRef]
- Nikolakopoulou, K.; Turco, M.Y. Investigation of infertility using endometrial organoids. Reproduction 2021, 161, R113–R127. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Drury, J.; Da Silva, L.; Al-Lamee, H.; Earp, A.; Valentijn, A.J.; Edirisinghe, D.P.; Murray, P.A.; Fazleabas, A.T.; Gargett, C.E. Abnormally located SSEA1+/SOX9+ endometrial epithelial cells with a basalis-like phenotype in the eutopic functionalis layer may play a role in the pathogenesis of endometriosis. Hum. Reprod. 2019, 34, 56–68. [Google Scholar] [CrossRef]
- Bishop, R.C.; Boretto, M.; Rutkowski, M.R.; Vankelecom, H.; Derré, I. Murine Endometrial Organoids to Model Chlamydia Infection. Front. Cell. Infect. Microbiol. 2020, 10, 416. [Google Scholar] [CrossRef]
- Dolat, L.; Valdivia, R.H. An endometrial organoid model of interactions between Chlamydia and epithelial and immune cells. J. Cell Sci. 2021, 134, jcs252403. [Google Scholar] [CrossRef]
- Yui, S.; Nakamura, T.; Sato, T.; Nemoto, Y.; Mizutani, T.; Zheng, X.; Ichinose, S.; Nagaishi, T.; Okamoto, R.; Tsuchiya, K.; et al. Functional engraftment of colon epithelium expanded in vitro from a single adult Lgr5+ stem cell. Nat. Med. 2012, 18, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Huch, M.; Gehart, H.; van Boxtel, R.; Hamer, K.; Blokzijl, F.; Verstegen, M.M.A.; Ellis, E.; van Wenum, M.; Fuchs, S.A.; de Ligt, J.; et al. Long-Term Culture of Genome-Stable Bipotent Stem Cells from Adult Human Liver. Cell 2015, 160, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Dossena, M.; Piras, R.; Cherubini, A.; Barilani, M.; Dugnani, E.; Salanitro, F.; Moreth, T.; Pampaloni, F.; Piemonti, L.; Lazzari, L. Standardized GMP-compliant scalable production of human pancreas organoids. Stem Cell Res. Ther. 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Jagannathan, N. Cytokeratin: A review on current concepts. Int. J. Orofac. Biol. 2018, 2, 6. [Google Scholar] [CrossRef]
- White, F.J.; Ross, J.W.; Joyce, M.M.; Geisert, R.D.; Burghardt, R.C.; Johnson, G.A. Steroid Regulation of Cell Specific Secreted Phosphoprotein 1 (Osteopontin) Expression in the Pregnant Porcine Uterus1. Biol. Reprod. 2005, 73, 1294–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, L.; Li, R.; Hartley, L.; Nandurkar, M.H.; Koentgen, F.; Glenn Begley, C. Infertility in female mice lacking the receptor for interleukin 11 is due to a defective uterine response to implantation. Nat. Med. 1998, 4, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Gamperl, M.; Burkard, T.R.; Kunihs, V.; Kaindl, U.; Junttila, S.; Fiala, C.; Schmidt, K.; Mendjan, S.; Knöfler, M.; et al. Estrogen Signaling Drives Ciliogenesis in Human Endometrial Organoids. Endocrinology 2019, 160, 2282–2297. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, C.; Chen, Y.; Zhao, J. The involvement of osteopontin and β3 integrin in implantation and endometrial receptivity in an early mouse pregnancy model. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 171–176. [Google Scholar] [CrossRef]
- Bell, S.C.; Drife, J.O. 4 Secretory proteins of the endometrium—potential markers for endometrial dysfunction. Baillieres. Clin. Obstet. Gynaecol. 1989, 3, 271–291. [Google Scholar] [CrossRef]
- Soni, C.; Karande, A.A. Glycodelin A suppresses the cytolytic activity of CD8+ T lymphocytes. Mol. Immunol. 2010, 47, 2458–2466. [Google Scholar] [CrossRef]
- Serafini, P.C.; Silva, I.D.C.G.; Smith, G.D.; Motta, E.L.A.; Rocha, A.M.; Baracat, E.C. Endometrial claudin-4 and leukemia inhibitory factor are associated with assisted reproduction outcome. Reprod. Biol. Endocrinol. 2009, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ruijter-Villani, M.; Deelen, C.; Stout, T.A.E. Expression of leukaemia inhibitory factor at the conceptus–maternal interface during preimplantation development and in the endometrium during the oestrous cycle in the mare. Reprod. Fertil. Dev. 2016, 28, 1642. [Google Scholar] [CrossRef] [PubMed]
- Hasdemir, P.; Farasat, M.; Aydin, C.; Ozyurt, B.; Guvenal, T.; Pekindil, G. The Role of Adenomyosis in the Pathogenesis of Preeclampsia. Geburtshilfe Frauenheilkd. 2016, 76, 882–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirgianakis, K.; Kalaitzopoulos, D.R.; Schwartz, A.S.K.; Spaanderman, M.; Kramer, B.W.; Mueller, M.D.; Mueller, M. Fertility, pregnancy and neonatal outcomes of patients with adenomyosis: A systematic review and meta-analysis. Reprod. Biomed. Online 2021, 42, 185–206. [Google Scholar] [CrossRef]
- Hashimoto, A.; Iriyama, T.; Sayama, S.; Nakayama, T.; Komatsu, A.; Miyauchi, A.; Nishii, O.; Nagamatsu, T.; Osuga, Y.; Fujii, T. Adenomyosis and adverse perinatal outcomes: Increased risk of second trimester miscarriage, preeclampsia, and placental malposition. J. Matern. Neonatal Med. 2018, 31, 364–369. [Google Scholar] [CrossRef]
- Xu, X.-H.; Jia, Y.; Zhou, X.; Xie, D.; Huang, X.; Jia, L.; Zhou, Q.; Zheng, Q.; Zhou, X.; Wang, K.; et al. Downregulation of lysyoxidase and lysyl oxidase-like protein 2 suppressed the migration and invasion of trophoblasts by activating the TGF-β/collagen pathway in preeclampsia. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef]
- Brkić, J.; Dunk, C.; Shan, Y.; O’Brien, J.A.; Lye, P.; Qayyum, S.; Yang, P.; Matthews, S.G.; Lye, S.J.; Peng, C. Differential Role of Smad2 and Smad3 in the Acquisition of an Endovascular Trophoblast-Like Phenotype and Preeclampsia. Front. Endocrinol. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Matheus Bremm, J.; Michels, M.; Duarte Rengel, B.; Gomes, F.G.; Fraga, L.R.; Sanseverino, M.T.V. Genetic and in silico analysis show a role of SMAD3 on recurrent pregnancy loss. Hum. Fertil. 2021. [Google Scholar] [CrossRef]
- Lappas, M. Identification of SMAD3 as a Novel Mediator of Inflammation in Human Myometrium In Vitro. Mediators Inflamm. 2018, 2018, 3140420. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Yang, X.; Sun, X.; Peng, C.; Li, X.; Wang, M.; Zhou, Y. Expression of integrin beta3 and osteopontin in endometrium of patients with adenomyosis. Zhonghua Fu Chan Ke Za Zhi 2009, 44, 354–358. [Google Scholar]
- Batorfi, J.; Fulop, V.; Kim, J.H.; Genest, D.R.; Doszpod, J.; Mok, S.C.; Berkowitz, R.S. Osteopontin is down-regulated in hydatidiform mole. Gynecol. Oncol. 2003, 89, 134–139. [Google Scholar] [CrossRef]
- Xiao, Y.; Sun, X.; Yang, X.; Zhang, J.; Xue, Q.; Cai, B.; Zhou, Y. Leukemia inhibitory factor is dysregulated in the endometrium and uterine flushing fluid of patients with adenomyosis during implantation window. Fertil. Steril. 2010, 94, 85–89. [Google Scholar] [CrossRef]
- Yen, C.F.; Liao, S.K.; Huang, S.J.; Tabak, S.; Arcuri, F.; Lee, C.L.; Arici, A.; Petraglia, F.; Wang, H.S.; Kayisli, U.A. Decreased Endometrial Expression of Leukemia Inhibitory Factor Receptor Disrupts the STAT3 Signaling in Adenomyosis during the Implantation Window. Reprod. Sci. 2017, 24, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Rosario, G.X.; Stewart, C.L. The Multifaceted Actions of Leukaemia Inhibitory Factor in Mediating Uterine Receptivity and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitawaki, J.; Kado, N.; Ishihara, H.; Koshiba, H.; Kitaoka, Y.; Honjo, H. Endometriosis: The pathophysiology as an estrogen-dependent disease. J. Steroid Biochem. Mol. Biol. 2002, 83, 149–155. [Google Scholar] [CrossRef]
- Kitawaki, J. Adenomyosis: The pathophysiology of an oestrogen-dependent disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 493–502. [Google Scholar] [CrossRef]
- Rantakari, P.; Strauss, L.; Kiviranta, R.; Lagerbohm, H.; Paviala, J.; Holopainen, I.; Vainio, S.; Pakarinen, P.; Poutanen, M. Placenta defects and embryonic lethality resulting from disruption of mouse hydroxysteroid (17-β) dehydrogenase 2 gene. Mol. Endocrinol. 2008, 22, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Abbas, Y.; Brunel, L.G.; Hollinshead, M.S.; Fernando, R.C.; Gardner, L.; Duncan, I.; Moffett, A.; Best, S.; Turco, M.Y.; Burton, G.J.; et al. Generation of a three-dimensional collagen scaffold-based model of the human endometrium. Interface Focus 2020, 10, 20190079. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Gordillo, V.; Kassis, T.; Lampejo, A.; Choi, G.; Gamboa, M.E.; Gnecco, J.S.; Brown, A.; Breault, D.T.; Carrier, R.; Griffith, L.G. Fully synthetic matrices for in vitro culture of primary human intestinal enteroids and endometrial organoids. Biomaterials 2020, 254, 120125. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, T.M.; Makwana, K.; Taylor, D.M.; Molè, M.A.; Fishwick, K.J.; Tryfonos, M.; Odendaal, J.; Hawkes, A.; Zernicka-Goetz, M.; Hartshorne, G.M.; et al. Modelling the impact of decidual senescence on embryo implantation in human endometrial assembloids. Elife 2021, 10, e69603. [Google Scholar] [CrossRef]
- Campo, H.; Murphy, A.; Yildiz, S.; Woodruff, T.; Cervelló, I.; Kim, J.J. Microphysiological Modeling of the Human Endometrium. Tissue Eng. Part A 2020, 26, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.-M.; Fellah, L. What if deep endometriotic nodules and uterine adenomyosis were actually two forms of the same disease? Fertil. Steril. 2019, 111, 454–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyendecker, G.; Bilgicyildirim, A.; Inacker, M.; Stalf, T.; Huppert, P.; Mall, G.; Böttcher, B.; Wildt, L. Adenomyosis and endometriosis. Re-visiting their association and further insights into the mechanisms of auto-traumatisation. An MRI study. Arch. Gynecol. Obstet. 2015, 291, 917–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapron, C.; Tosti, C.; Marcellin, L.; Bourdon, M.; Lafay-Pillet, M.-C.; Millischer, A.-E.; Streuli, I.; Borghese, B.; Petraglia, F.; Santulli, P. Relationship between the magnetic resonance imaging appearance of adenomyosis and endometriosis phenotypes. Hum. Reprod. 2017, 32, 1393–1401. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Company | Product Number | Concentration in ExM |
|---|---|---|---|
| Advance DMEM/F12 | Life Technologies | 12634010 | 1X |
| N2 supplement | Life Technologies | 17502048 | 1X |
| B27 supplement minus vitamin A | Life Technologies | 12587010 | 1X |
| Primocin | Invivogen | ant-pm-1 | 100 μg/ml |
| N-Acetyl-L-cysteine | Sigma | A9165 | 1.25 mM |
| L-glutamine | Sigma | G7513 | 2 mM |
| Recombinant human EGF | Peprotech | AF-100-15 | 50 ng/ml |
| Recombinant human Noggin | Peprotech | 120-10c | 100 ng/ml |
| Recombinant human Rspondin-1 | Peprotech | 120-38 | 500 ng/ml |
| Recombinant human FGF-10 | Peprotech | 100-26 | 100 ng/ml |
| Recombinant human HGF | Peprotech | 100-39 | 50 ng/ml |
| ALK-4, -5, -7 inhibitor, A83-01 | Peprotech | 9094360 | 500 nM |
| Nicotinamide | Sigma | N0636 | 10 nM |
| Antibody | Company | Product Number | Concentration |
|---|---|---|---|
| Anti-MUC-1 | Abcam | ab109185 | 1:250 |
| Anti-SOX9 | Abcam | ab185966 | 1:100 |
| Anti-Ki67 | Dako | M7240 | 1:100 |
| Anti-TGF-β2 | Abcam | ab36495 | 1:1000 |
| Anti-Smad3 | Abcam | ab40854 | 1:500 |
| Anti-PanCK | Abcam | ab86734 | 1:100 |
| Anti-Vimentin | Abcam | ab92547 | 1:250 |
| Anti-Laminin | Abcam | ab11575 | 1:200 |
| Anti-acetylated α-tubulin | Santa Cruz Bt | 611B1 | 1:500 |
| AlexaFluor 488 goat antimouse IgG1 | Invitrogen | A21121 | 1:500 |
| AlexaFluor 555 goat antirabbit IgG | Invitrogen | A21429 | 1:500 |
| AlexaFluor 488 goat antirabbit IgG | Invitrogen | A11034 | 1:500 |
| AlexaFluor 488 goat antimouse IgG | Invitrogen | A11029 | 1:500 |
| GENE | Forward Sequence | Reverse Sequence |
|---|---|---|
| SPP1 | CGAGGTGATAGTGTGGTTTATG | GTCTGTAGCATCAGGGTACT |
| PAEP | ATGGCGACCAACAACATC | CTCTCCAAGGACCTTCTTCT |
| LIF | AACTGGCACAGCTCAATG | ATAGCTTGTCCAGGTTGTTG |
| 17HSDß2 | TGAATGTCAGCAGCATGG | GGAAAGCTCCAGTCTCATAAC |
| GAPDH | AACGTGTCAGTGGTGGACCTGA | ACCACCCTGTTGCTGTAGCCAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juárez-Barber, E.; Francés-Herrero, E.; Corachán, A.; Vidal, C.; Giles, J.; Alamá, P.; Faus, A.; Pellicer, A.; Cervelló, I.; Ferrero, H. Establishment of Adenomyosis Organoids as a Preclinical Model to Study Infertility. J. Pers. Med. 2022, 12, 219. https://doi.org/10.3390/jpm12020219
Juárez-Barber E, Francés-Herrero E, Corachán A, Vidal C, Giles J, Alamá P, Faus A, Pellicer A, Cervelló I, Ferrero H. Establishment of Adenomyosis Organoids as a Preclinical Model to Study Infertility. Journal of Personalized Medicine. 2022; 12(2):219. https://doi.org/10.3390/jpm12020219
Chicago/Turabian StyleJuárez-Barber, Elena, Emilio Francés-Herrero, Ana Corachán, Carmina Vidal, Juan Giles, Pilar Alamá, Amparo Faus, Antonio Pellicer, Irene Cervelló, and Hortensia Ferrero. 2022. "Establishment of Adenomyosis Organoids as a Preclinical Model to Study Infertility" Journal of Personalized Medicine 12, no. 2: 219. https://doi.org/10.3390/jpm12020219
APA StyleJuárez-Barber, E., Francés-Herrero, E., Corachán, A., Vidal, C., Giles, J., Alamá, P., Faus, A., Pellicer, A., Cervelló, I., & Ferrero, H. (2022). Establishment of Adenomyosis Organoids as a Preclinical Model to Study Infertility. Journal of Personalized Medicine, 12(2), 219. https://doi.org/10.3390/jpm12020219