
Epigenetics in Precision Nutrition



Abstract
:1. Introduction
2. DNA Methylation in Obesity, T2D, and CVD
2.1. DNA Methylation and Obesity
2.2. DNA Methylation and T2D
2.3. DNA Methylation and CVD
3. Nutrition and DNA Methylation
4. Diet and Lifestyle Interventions on DNA Methylation
5. Other Epigenetic Events
6. Summary and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qi, L. Gene–diet interaction and weight loss. Curr. Opin. Lipidol. 2014, 25, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitipaldi, H.; McCarthy, M.I.; Florez, J.C.; Franks, P.W. A Global Overview of Precision Medicine in Type 2 Diabetes. Diabetes 2018, 67, 1911–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, J.; Florez, J.C. Precision medicine in diabetes: An opportunity for clinical translation. Ann. N. Y. Acad. Sci. 2018, 1411, 140–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, C.J.; Horgan, S.; Neary, R.; Glezeva, N.; Tea, I.; Corrigan, N.; Mcdonald, K.; Ledwidge, M.; Baugh, J. Epigenetic therapy for the treatment of hypertension-induced cardiac hypertrophy and fibrosis. J. Cardiovasc. Pharmacol. Ther. 2016, 21, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Ciechomska, M.; Roszkowski, L.; Maslinski, W. DNA Methylation as a Future Therapeutic and Diagnostic Target in Rheumatoid Arthritis. Cells 2019, 8, 953. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lay, F.; Han, H.; Jones, P.A. Targeting DNA methylation for epigenetic therapy. Trends Pharmacol. Sci. 2010, 31, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef]
- World Obesity Federation. Prevalence of Obesity. n.d. Available online: https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity (accessed on 8 February 2022).
- Dick, K.J.; Nelson, C.; Tsaprouni, L.; Sandling, J.; Aïssi, D.; Wahl, S.; Meduri, E.; Morange, P.-E.; Gagnon, F.; Grallert, H.; et al. DNA methylation and body-mass index: A genome-wide analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Demerath, E.W.; Guan, W.; Grove, M.L.; Aslibekyan, S.; Mendelson, M.; Zhou, Y.-H.; Hedman, Å.K.; Sandling, J.K.; Li, L.-A.; Irvin, M.R.; et al. Epigenome-wide association study (EWAS) of BMI, BMI change and waist circumference in African American adults identifies multiple replicated loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef] [Green Version]
- Aslibekyan, S.; Demerath, E.W.; Mendelson, M.; Zhi, D.; Guan, W.; Liang, L.; Sha, J.; Pankow, J.; Liu, C.; Irvin, M.R.; et al. Epigenome-wide study identifies novel methylation loci associated with body mass index and waist circumference. Obesity 2015, 23, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Mendelson, M.; Marioni, R.; Joehanes, R.; Liu, C.; Hedman, Å.K.; Aslibekyan, S.; Demerath, E.W.; Guan, W.; Zhi, D.; Yao, C.; et al. Association of Body Mass Index with DNA Methylation and Gene Expression in Blood Cells and Relations to Cardiometabolic Disease: A Mendelian Randomization Approach. PLoS Med. 2017, 14, e1002215. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.-C.; Ried, J.S.; Zhang, W.; Yang, Y.; et al. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2017, 541, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Melton, P.; Burton, M.; Beilin, L.; Clarke-Harris, R.; Cook, E.; Godfrey, K.; Burdge, G.; Mori, T.; Anderson, D.; et al. Adiposity associated DNA methylation signatures in adolescents are related to leptin and perinatal factors. Epigenetics 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kassam, I.; Lau, S.H.; Kooner, J.S.; Wilson, R.; Peters, A.; Winkelmann, J.; Chambers, J.C.; Chow, V.T.; Khor, C.C.; et al. Impact of BMI and waist circumference on epigenome-wide DNA methylation and identification of epigenetic biomarkers in blood: An EWAS in multi-ethnic Asian individuals. Clin. Epigenetics 2021, 13, 195. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the primary prevention of cardiovascular disease: A report of the american college of cardiology/American heart association task force on clinical practice guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef]
- American Diabetes Association. 8. Obesity Management for the Treatment of Type 2 Diabetes: Standards of Medical Care in Diabetes—2019. Diabetes Care 2018, 42, S81–S89. [Google Scholar] [CrossRef] [Green Version]
- Zimmet, P.; Alberti, K.G.; Magliano, D.J.; Bennett, P.H. Diabetes mellitus statistics on prevalence and mortality: Facts and fallacies. Nat. Rev. Endocrinol. 2016, 12, 616–622. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes–Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Cameron, N.A.; Petito, L.C.; McCabe, M.; Allen, N.B.; O’Brien, M.J.; Carnethon, M.R.; Khan, S.S. Quantifying the Sex-Race/Ethnicity-Specific Burden of Obesity on Incident Diabetes Mellitus in the United States, 2001 to 2016: MESA and NHANES. J. Am. Heart Assoc. 2021, 10, e018799. [Google Scholar] [CrossRef]
- Hu, Y.; Ding, M.; Sampson, L.; Willett, W.C.; Manson, J.E.; Wang, M.; Rosner, B.; Hu, F.B.; Sun, Q. Intake of whole grain foods and risk of type 2 diabetes: Results from three prospective cohort studies. BMJ 2020, 370, m2206. [Google Scholar] [CrossRef]
- Qian, F.; Liu, G.; Hu, F.B.; Bhupathiraju, S.N.; Sun, Q. Association Between Plant-Based Dietary Patterns and Risk of Type 2 Diabetes: A Systematic Review and Meta-analysis. JAMA Intern. Med. 2019, 179, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Avery, L.; Flynn, D.; Van Wersch, A.; Sniehotta, F.F.; Trenell, M.I. Changing physical activity behavior in type 2 diabetes: A systematic review and meta-analysis of behavioral interventions. Diabetes Care 2012, 35, 2681–2689. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.; Majewski, C.; Xie, M.; Yan, P.; Guo, Y.; Bao, W.; Rong, Y.; Jackson, C.L.; Hu, F.B.; Liu, L. Sleep Duration and Risk of Type 2 Diabetes: A Meta-analysis of Prospective Studies. Diabetes Care 2015, 38, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhou, T.; Li, X.; Ma, H.; Liang, Z.; Fonseca, V.A.; Heianza, Y.; Qi, L. Baseline Vitamin D Status, Sleep Patterns, and the Risk of Incident Type 2 Diabetes in Data From the UK Biobank Study. Diabetes Care 2020, 43, 2776–2784. [Google Scholar] [CrossRef]
- Raciti, G.A.; Longo, M.; Parrillo, L.; Ciccarelli, M.; Mirra, P.; Ungaro, P.; Formisano, P.; Miele, C.; Beguinot, F. Understanding type 2 diabetes: From genetics to epigenetics. Acta Diabetol. 2015, 52, 821–827. [Google Scholar] [CrossRef]
- Florath, I.; Butterbach, K.; Heiss, J.A.; Bewerunge-Hudler, M.; Zhang, Y.; Schöttker, B.; Brenner, H. Type 2 diabetes and leucocyte DNA methylation: An epigenome-wide association study in over 1500 older adults. Diabetologia 2016, 59, 130–138. [Google Scholar] [CrossRef]
- Soriano-Tárraga, C.; Jiménez-Conde, J.; Giralt-Steinhauer, E.; Mola-Caminal, M.; Vivanco-Hidalgo, R.M.; Ois, A.; Rodríguez-Campello, A.; Cuadrado-Godia, E.; Sayols-Baixeras, S.; Elosua, R.; et al. Epigenome-wide association study identifies TXNIP gene associated with type 2 diabetes mellitus and sustained hyperglycemia. Hum. Mol. Genet. 2016, 25, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Cardona, A.; Day, F.R.; Perry, J.R.B.; Loh, M.; Chu, A.Y.; Lehne, B.; Paul, D.S.; Lotta, L.A.; Stewart, I.D.; Kerrison, N.D.; et al. Epigenome-Wide Association Study of Incident Type 2 Diabetes in a British Population: EPIC-Norfolk Study. Diabetes 2019, 68, 2315–2326. [Google Scholar] [CrossRef] [Green Version]
- Meeks, K.A.C.; Henneman, P.; Venema, A.; Addo, J.; Bahendeka, S.; Burr, T.; Danquah, I.; Galbete, C.; Mannens, M.M.A.M.; Mockenhaupt, F.P.; et al. Epigenome-wide association study in whole blood on type 2 diabetes among sub-Saharan African individuals: Findings from the RODAM study. Int. J. Epidemiol. 2019, 48, 58–70. [Google Scholar] [CrossRef]
- Al Muftah, W.A.; Al-Shafai, M.; Zaghlool, S.B.; Visconti, A.; Tsai, P.; Kumar, P.; Spector, T.; Bell, J.; Falchi, M.; Suhre, K. Epigenetic associations of type 2 diabetes and BMI in an Arab population. Clin. Epigenetics 2016, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, H.; Kos, M.Z.; Neary, J.; Dyer, T.D.; Kent, J.W., Jr.; Göring, H.H.H.; Cole, S.A.; Comuzzie, A.G.; Almasy, L.; Mahaney, M.C.; et al. Novel epigenetic determinants of type 2 diabetes in Mexican-American families. Hum. Mol. Genet. 2015, 24, 5330–5344. [Google Scholar] [CrossRef]
- Dayeh, T.; Volkov, P.; Salö, S.; Hall, E.; Nilsson, E.; Olsson, A.H.; Kirkpatrick, C.; Wollheim, C.B.; Eliasson, L.; Rönn, T.; et al. Genome-Wide DNA Methylation Analysis of Human Pancreatic Islets from Type 2 Diabetic and Non-Diabetic Donors Identifies Candidate Genes That Influence Insulin Secretion. PLoS Genet. 2014, 10, e1004160. [Google Scholar] [CrossRef]
- Juvinao-Quintero, D.L.; Marioni, R.E.; Ochoa-Rosales, C.; Russ, T.C.; Deary, I.J.; van Meurs, J.B.J.; Voortman, T.; Hivert, M.-F.; Sharp, G.C.; Relton, C.L.; et al. DNA methylation of blood cells is associated with prevalent type 2 diabetes in a meta-analysis of four European cohorts. Clin. Epigenetics 2021, 13, 40. [Google Scholar] [CrossRef]
- Liang, F.; Quan, Y.; Wu, A.; Chen, Y.; Xu, R.; Zhu, Y.; Xiong, J. Insulin-resistance and depression cohort data mining to identify nutraceutical related DNA methylation biomarker for type 2 diabetes. Genes Dis. 2020, 8, 669–676. [Google Scholar] [CrossRef]
- Nilsson, E.; Vavakova, M.; Perfilyev, A.; Säll, J.; Jansson, P.-A.; Poulsen, P.; Esguerra, J.L.S.; Eliasson, L.; Vaag, A.; Göransson, O.; et al. Differential DNA Methylation and Expression of miRNAs in Adipose Tissue From Twin Pairs Discordant for Type 2 Diabetes. Diabetes 2021, 70, 2402–2418. [Google Scholar] [CrossRef]
- Dayeh, T.; Tuomi, T.; Almgren, P.; Perfilyev, A.; Jansson, P.-A.; De Mello, V.D.; Pihlajamäki, J.; Vaag, A.; Groop, L.; Nilsson, E.; et al. DNA methylation of loci within ABCG1 and PHOSPHO1 in blood DNA is associated with future type 2 diabetes risk. Epigenetics 2016, 11, 482–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, J.C.; Loh, M.; Lehne, B.; Drong, A.; Kriebel, J.; Motta, V.; Wahl, S.; Elliott, H.; Rota, F.; Scott, W.R.; et al. Epigenome-wide association of DNA methylation markers in peripheral blood from Indian Asians and Europeans with incident type 2 diabetes: A nested case-control study. Lancet Diabetes Endocrinol. 2015, 3, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Fraszczyk, E.; Spijkerman, A.M.W.; Zhang, Y.; Brandmaier, S.; Day, F.R.; Zhou, L.; Wackers, P.; Dollé, M.E.T.; Bloks, V.W.; Gào, X. Epigenome-wide association study of incident type 2 diabetes: A meta-analysis of five prospective European cohorts. Diabetologia 2022, 1–14. [Google Scholar] [CrossRef]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update: A Report From the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- Heart Disease Facts. Available online: https://www.cdc.gov/heartdisease/facts.htm (accessed on 23 February 2022).
- Costantino, S.; Libby, P.; Kishore, R.; Tardif, J.-C.; El-Osta, A.; Paneni, F. Epigenetics and precision medicine in cardiovascular patients: From basic concepts to the clinical arena. Eur. Heart J. 2017, 39, 4150–4158. [Google Scholar] [CrossRef] [PubMed]
- Westerman, K.E.; Ordovás, J.M. DNA methylation and incident cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sanlés, A.; Sayols-Baixeras, S.; Subirana, I.; Sentí, M.; Pérez-Fernández, S.; Moura, M.D.C.; Esteller, M.; Marrugat, J.; Elosua, R. DNA methylation biomarkers of myocardial infarction and cardiovascular disease. Clin. Epigenetics 2021, 13, 86. [Google Scholar] [CrossRef]
- Rask-Andersen, M.; Martinsson, D.; Ahsan, M.; Enroth, S.; Ek, W.E.; Gyllensten, U.; Johansson, Å. Epigenome-wide association study reveals differential DNA methylation in individuals with a history of myocardial infarction. Hum. Mol. Genet. 2016, 25, 4739–4748. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhu, X.; Yu, K.; Jiang, H.; Zhang, Y.; Deng, S.; Cheng, L.; Liu, X.; Zhong, J.; Zhang, X.; et al. Genome-Wide Analysis of DNA Methylation and Acute Coronary Syndrome. Circ. Res. 2017, 120, 1754–1767. [Google Scholar] [CrossRef]
- Nakatochi, M.; Ichihara, S.; Yamamoto, K.; Naruse, K.; Yokota, S.; Asano, H.; Matsubara, T.; Yokota, M. Epigenome-wide association of myocardial infarction with DNA methylation sites at loci related to cardiovascular disease. Clin. Epigenetics 2017, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portilla-Fernández, E.; Hwang, S.-J.; Wilson, R.; Maddock, J.; Hill, W.D.; Teumer, A.; Mishra, P.P.; Brody, J.A.; Joehanes, R.; Ligthart, S.; et al. Meta-analysis of epigenome-wide association studies of carotid intima-media thickness. Eur. J. Epidemiol. 2021, 36, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Agha, G.; Mendelson, M.M.; Ward-Caviness, C.K.; Joehanes, R.; Huan, T.; Gondalia, R.; Salfati, E.; Brody, J.A.; Fiorito, G.; Bressler, J.; et al. Blood Leukocyte DNA Methylation Predicts Risk of Future Myocardial Infarction and Coronary Heart Disease. Circulation 2019, 140, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Yang, S.; Sun, D.; Yu, C.; Guo, Y.; Lin, Y.; Millwood, I.Y.; Walters, R.G.; Yang, L.; Chen, Y.; et al. Epigenome-wide analysis of DNA methylation and coronary heart disease: A nested case-control study. eLife 2021, 10, e68671. [Google Scholar] [CrossRef]
- Aslibekyan, S.; Agha, G.; Colicino, E.; Do, A.N.; Lahti, J.; Ligthart, S.; Marioni, R.; Marzi, C.; Mendelson, M.; Tanaka, T.; et al. Association of Methylation Signals With Incident Coronary Heart Disease in an Epigenome-Wide Assessment of Circulating Tumor Necrosis Factor α. JAMA Cardiol. 2018, 3, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Pang, H.; Chen, B.; Wu, C.; Wang, Y.; Hou, L.; Wang, S.; Sun, D.; Zheng, X. Genome-wide analysis of DNA methylation and risk of cardiovascular disease in a Chinese population. BMC Cardiovasc. Disord. 2021, 21, 240. [Google Scholar] [CrossRef] [PubMed]
- Westerman, K.; Sebastiani, P.; Jacques, P.; Liu, S.; DeMeo, D.; Ordovás, J.M. DNA methylation modules associate with incident cardiovascular disease and cumulative risk factor exposure. Clin. Epigenetics 2019, 11, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Sanlés, A.; Baixeras, S.S.; Subirana, I.; Degano, I.R.; Elosua, R. Association between DNA methylation and coronary heart disease or other atherosclerotic events: A systematic review. Atherosclerosis 2017, 263, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, O.S.; Sant, K.E.; Dolinoy, D.C. Nutrition and epigenetics: An interplay of dietary methyl donors, one-carbon metabolism and DNA methylation. J. Nutr. Biochem. 2012, 23, 853–859. [Google Scholar] [CrossRef] [Green Version]
- Barouki, R.; Gluckman, P.D.; Grandjean, P.; Hanson, M.; Heindel, J.J. Developmental origins of non-communicable disease: Implications for research and public health. Environ. Health 2012, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [Green Version]
- Chao, W.; D’Amore, P.A. IGF2: Epigenetic regulation and role in development and disease. Cytokine Growth Factor Rev. 2008, 19, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.; Lumey, L.H.; Talens, R.P.; Kremer, D.; Putter, H.; Stein, A.; Slagboom, P.; Heijmans, B.T. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef]
- Tobi, E.; Goeman, J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.; Stok, A.P.; Thijssen, P.E.; Müller, F.; et al. DNA methylation signatures link prenatal famine exposure to growth and metabolism. Nat. Commun. 2014, 5, 5592. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Slagboom, P.E.; van Zwet, E.W.; Lumey, L.H.; Heijmans, B.T.; et al. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci. Adv. 2018, 4, eaao4364. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Song, J.; Li, C.; Li, Y.; Shen, L.; Dong, B.; Zou, Z.; Ma, J. DNA methylation of the INSR gene as a mediator of the association between prenatal exposure to famine and adulthood waist circumference. Sci. Rep. 2020, 10, 12212. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, C.; Wang, Z.; Zhang, R.; Shen, Y.; Miles, T.; Wei, J.; Zou, Z. Early-life exposure to severe famine is associated with higher methylation level in the IGF2 gene and higher total cholesterol in late adulthood: The Genomic Research of the Chinese Famine (GRECF) study. Clin. Epigenetics 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Finer, S.; Iqbal, M.S.; Lowe, R.; Ogunkolade, B.; Pervin, S.; Mathews, C.; Smart, M.; Alam, D.S.; Hitman, G.A. Data from: Is famine exposure during developmental life in rural Bangladesh associated with a metabolic and epigenetic signature in young adulthood? A historical cohort study. BMJ Open 2016, 6, e011768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterland, R.A.; Kellermayer, R.; Laritsky, E.; Rayco-Solon, P.; Harris, R.A.; Travisano, M.; Zhang, W.; Torskaya, M.S.; Zhang, J.; Shen, L.; et al. Season of Conception in Rural Gambia Affects DNA Methylation at Putative Human Metastable Epialleles. PLoS Genet. 2010, 6, e1001252. [Google Scholar] [CrossRef] [Green Version]
- Hoyo, C.; Murtha, A.P.; Schildkraut, J.M.; Jirtle, R.L.; Demark-Wahnefried, W.; Forman, M.R.; Iversen, E.S.; Kurtzberg, J.; Overcash, F.; Huang, Z.; et al. Methylation variation at IGF2 differentially methylated regions and maternal folic acid use before and during pregnancy. Epigenetics 2011, 6, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, K.M.; Sheppard, A.; Gluckman, P.D.; Lillycrop, K.A.; Burdge, G.C.; McLean, C.; Rodford, J.; Slater-Jefferies, J.L.; Garratt, E.; Crozier, S.R.; et al. Epigenetic Gene Promoter Methylation at Birth Is Associated With Child’s Later Adiposity. Diabetes 2011, 60, 1528–1534. [Google Scholar] [CrossRef] [Green Version]
- Amarasekera, M.; Martino, D.; Ashley, S.; Harb, H.; Kesper, D.; Strickland, D.; Saffery, R.; Prescott, S.L. Genome-wide DNA methylation profiling identifies a folate-sensitive region of differential methylation upstream of ZFP57 -imprinting regulator in humans. FASEB J. 2014, 28, 4068–4076. [Google Scholar] [CrossRef] [Green Version]
- Boeke, C.E.; Baccarelli, A.; Kleinman, K.P.; Burris, H.H.; Litonjua, A.A.; Rifas-Shiman, S.L.; Tarantini, L.; Gillman, M. Gestational intake of methyl donors and global LINE-1 DNA methylation in maternal and cord blood: Prospective results from a folate-replete population. Epigenetics 2012, 7, 253–260. [Google Scholar] [CrossRef] [Green Version]
- McKay, J.A.; Groom, A.; Potter, C.; Coneyworth, L.J.; Ford, D.; Mathers, J.C.; Relton, C. Genetic and Non-Genetic Influences during Pregnancy on Infant Global and Site Specific DNA Methylation: Role for Folate Gene Variants and Vitamin B12. PLoS ONE 2012, 7, e33290. [Google Scholar] [CrossRef]
- Azzi, S.; Sas, T.C.J.; Koudou, Y.; Le Bouc, Y.; Souberbielle, J.-C.; Dargent-Molina, P.; Netchine, I.; Charles, M.A. Degree of methylation ofZAC1(PLAGL1) is associated with prenatal and post-natal growth in healthy infants of the EDEN mother child cohort. Epigenetics 2014, 9, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.L.; Jima, D.; Sharp, G.C.; McCullough, L.E.; Park, S.S.; Gowdy, K.; Skaar, D.; Cowley, M.; Maguire, R.L.; Fuemmeler, B.; et al. Maternal pre-pregnancy obesity, offspring cord blood DNA methylation, and offspring cardiometabolic health in early childhood: An epigenome-wide association study. Epigenetics 2019, 14, 325–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margerison-Zilko, C.E.; Shrimali, B.P.; Eskenazi, B.; Lahiff, M.; Lindquist, A.R.; Abrams, B.F. Trimester of Maternal Gestational Weight Gain and Offspring Body Weight at Birth and Age Five. Matern. Child Health J. 2012, 16, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Oken, E.; Levitan, E.B.; Gillman, M.W. Maternal smoking during pregnancy and child overweight: Systematic review and meta-analysis. Int. J. Obes. 2008, 32, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valsesia, A.; Wang, Q.-P.; Gheldof, N.; Carayol, J.; Ruffieux, H.; Clark, T.; Shenton, V.; Oyston, L.; Lefebvre, G.; Metairon, S.; et al. Genome-wide gene-based analyses of weight loss interventions identify a potential role for NKX6.3 in metabolism. Nat. Commun. 2019, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- Canouil, M.; Khamis, A.; Keikkala, E.; Hummel, S.; Lobbens, S.; Bonnefond, A.; Delahaye, F.; Tzala, E.; Mustaniemi, S.; Vääräsmäki, M.; et al. Epigenome-Wide Association Study Reveals Methylation Loci Associated With Offspring Gestational Diabetes Mellitus Exposure and Maternal Methylome. Diabetes Care 2021, 44, 1992–1999. [Google Scholar] [CrossRef]
- Tobi, E.W.; Juvinao-Quintero, D.L.; Ronkainen, J.; Ott, R.; Alfano, R.; Canouil, M.; Geurtsen, M.L.; Khamis, A.; Küpers, L.K.; Lim, I.Y.; et al. Maternal Glycemic Dysregulation During Pregnancy and Neonatal Blood DNA Methylation: Meta-analyses of Epigenome-Wide Association Studies. Diabetes Care 2022, 45, 614–623. [Google Scholar] [CrossRef]
- Nogues, P.; Dos Santos, E.; Jammes, H.; Berveiller, P.; Arnould, L.; Vialard, F.; Dieudonné, M.-N. Maternal obesity influences expression and DNA methylation of the adiponectin and leptin systems in human third-trimester placenta. Clin. Epigenetics 2019, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, L.; Thibault, S.; Guay, S.-P.; Santure, M.; Monpetit, A.; St-Pierre, J.; Perron, P.; Brisson, D. Leptin Gene Epigenetic Adaptation to Impaired Glucose Metabolism During Pregnancy. Diabetes Care 2010, 33, 2436–2441. [Google Scholar] [CrossRef] [Green Version]
- Joubert, B.R.; Håberg, S.E.; Nilsen, R.M.; Wang, X.; Vollset, S.E.; Murphy, S.K.; Huang, Z.; Hoyo, C.; Midttun, Ø.; Cupul-Uicab, L.A.; et al. 450K Epigenome-Wide Scan Identifies Differential DNA Methylation in Newborns Related to Maternal Smoking during Pregnancy. Environ. Health Perspect. 2012, 120, 1425–1431. [Google Scholar] [CrossRef] [Green Version]
- Richmond, R.C.; Simpkin, A.J.; Woodward, G.; Gaunt, T.R.; Lyttleton, O.; McArdle, W.L.; Ring, S.M.; Smith, A.D.; Timpson, N.J.; Tilling, K.; et al. Prenatal exposure to maternal smoking and offspring DNA methylation across the lifecourse: Findings from the Avon Longitudinal Study of Parents and Children (ALSPAC). Hum. Mol. Genet. 2015, 24, 2201–2217. [Google Scholar] [CrossRef] [Green Version]
- Novakovic, B.; Ryan, J.; Pereira, N.; Boughton, B.; Craig, J.M.; Saffery, R. Postnatal stability, tissue, and time specific effects of AHRR methylation change in response to maternal smoking in pregnancy. Epigenetics 2013, 9, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küpers, L.K.; Xu, X.; Jankipersadsing, S.A.; Vaez, A.; Gemert, S.L.B.-V.; Scholtens, S.; Nolte, I.M.; Richmond, R.C.; Relton, C.L.; Felix, J.F.; et al. DNA methylation mediates the effect of maternal smoking during pregnancy on birthweight of the offspring. Int. J. Epidemiology 2015, 44, 1224–1237. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.C.; Kelsey, K.T.; Zheng, S.; Houseman, E.A.; Marsit, C.; Wrensch, M.R.; Wiemels, J.L.; Nelson, H.; Karagas, M.R.; Kushi, L.; et al. Breast Cancer DNA Methylation Profiles Are Associated with Tumor Size and Alcohol and Folate Intake. PLoS Genet. 2010, 6, e1001043. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Marioni, R.E.; Hedman, Å.K.; Pfeiffer, L.; Tsai, P.-C.; Reynolds, L.M.; Just, A.C.; Duan, Q.; Boer, C.G.; Tanaka, T.; et al. A DNA methylation biomarker of alcohol consumption. Mol. Psychiatry 2018, 23, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Karabegović, I.; Portilla-Fernandez, E.; Li, Y.; Ma, J.; Maas, S.C.E.; Sun, D.; Hu, E.A.; Kühnel, B.; Zhang, Y.; Ambatipudi, S.; et al. Epigenome-wide association meta-analysis of DNA methylation with coffee and tea consumption. Nat. Commun. 2021, 12, 2830. [Google Scholar] [CrossRef] [PubMed]
- Shelnutt, K.; Kauwell, G.P.; Gregory, J.; Maneval, D.R.; Quinlivan, E.; Theriaque, D.W.; Henderson, G.N.; Bailey, L.B. Methylenetetrahydrofolate reductase 677C→T polymorphism affects DNA methylation in response to controlled folate intake in young women. J. Nutr. Biochem. 2004, 15, 554–560. [Google Scholar] [CrossRef]
- Ma, J.; Rebholz, C.M.; Braun, K.V.; Reynolds, L.M.; Aslibekyan, S.; Xia, R.; Biligowda, N.G.; Huan, T.; Liu, C.; Mendelson, M.M.; et al. Whole Blood DNA Methylation Signatures of Diet Are Associated With Cardiovascular Disease Risk Factors and All-Cause Mortality. Circ. Genom. Precis. Med. 2020, 13, e002766. [Google Scholar] [CrossRef]
- Van Dijk, S.J.; EpiSCOPE, M.O.; Molloy, P.; Varinli, H.; Morrison, J.; Muhlhausler, B.S. Epigenetics and human obesity. Int. J. Obes. 2015, 39, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Ling, C.; Groop, L. Epigenetics: A Molecular Link Between Environmental Factors and Type 2 Diabetes. Diabetes 2009, 58, 2718–2725. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Meir, A.Y.; Bernhart, S.H.; Gepner, Y.; Shelef, I.; Schwarzfuchs, D.; Tsaban, G.; Zelicha, H.; Hopp, L.; Müller, L.; et al. DNA methylation signature in blood mirrors successful weight-loss during lifestyle interventions: The CENTRAL trial. Genome Med. 2020, 12, 97. [Google Scholar] [CrossRef]
- Rönn, T.; Volkov, P.; Davegårdh, C.; Dayeh, T.; Hall, E.; Olsson, A.H.; Nilsson, E.; Tornberg, A.; Nitert, M.D.; Eriksson, K.-F.; et al. A Six Months Exercise Intervention Influences the Genome-wide DNA Methylation Pattern in Human Adipose Tissue. PLoS Genet. 2013, 9, e1003572. [Google Scholar] [CrossRef] [PubMed]
- Moleres, A.; Campión, J.; Milagro, F.I.; Marcos, A.; Campoy, C.; Garagorri, J.M.; Gómez-Martínez, S.; Martínez, J.A.; Azcona-Sanjulián, M.C.; Martí, A.; et al. Differential DNA methylation patterns between high and low responders to a weight loss intervention in overweight or obese adolescents: The EVASYON study. FASEB J. 2013, 27, 2504–2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitert, M.D.; Dayeh, T.; Volkov, P.; Elgzyri, T.; Hall, E.; Nilsson, E.; Yang, B.T.; Lang, S.; Parikh, H.; Wessman, Y.; et al. Impact of an Exercise Intervention on DNA Methylation in Skeletal Muscle From First-Degree Relatives of Patients with Type 2 Diabetes. Diabetes 2012, 61, 3322–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollepalli, S.; Kaye, S.; Heinonen, S.; Kaprio, J.; Rissanen, A.; A Virtanen, K.; Pietiläinen, K.; Ollikainen, M. Subcutaneous adipose tissue gene expression and DNA methylation respond to both short- and long-term weight loss. Int. J. Obes. 2018, 42, 412–423. [Google Scholar] [CrossRef]
- Gepner, Y.; Shelef, I.; Schwarzfuchs, D.; Zelicha, H.; Tene, L.; Meir, A.Y.; Tsaban, G.; Cohen, N.; Bril, N.; Rein, M.; et al. Effect of Distinct Lifestyle Interventions on Mobilization of Fat Storage Pools. Circulation 2018, 137, 1143–1157. [Google Scholar] [CrossRef]
- Meir, A.Y.; Keller, M.; Bernhart, S.H.; Rinott, E.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Schwarzfuchs, D.; Shelef, I.; Gepner, Y.; et al. Lifestyle weight-loss intervention may attenuate methylation aging: The CENTRAL MRI randomized controlled trial. Clin. Epigenetics 2021, 13, 48. [Google Scholar] [CrossRef]
- Li, X.; Shao, X.; Bazzano, L.A.; Xue, Q.; Koseva, B.S.; Grundberg, E.; Shai, I.; Bray, G.A.; Sacks, F.M.; Qi, L. Blood DNA methylation at TXNIP and glycemic changes in response to weight-loss diet interventions: The POUNDS lost trial. Int. J. Obes. 2022, 1–6. [Google Scholar] [CrossRef]
- Barres, R.; Kirchner, H.; Rasmussen, M.; Yan, J.; Kantor, F.R.; Krook, A.; Naslund, E.; Zierath, J.R. Weight loss after gastric bypass surgery in human obesity remodels promoter methylation. Cell Rep. 2013, 3, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, C.F.; Nonino, C.B.; de Oliveira, B.A.P.; Pinhel, M.A.D.S.; Mansego, M.L.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. DNA Methylation and Hydroxymethylation Levels in Relation to Two Weight Loss Strategies: Energy-Restricted Diet or Bariatric Surgery. Obes. Surg. 2015, 26, 603–611. [Google Scholar] [CrossRef]
- Ahrens, M.; Ammerpohl, O.; von Schönfels, W.; Kolarova, J.; Bens, S.; Itzel, T.; Teufel, A.; Herrmann, A.; Brosch, M.; Hinrichsen, H.; et al. DNA Methylation Analysis in Nonalcoholic Fatty Liver Disease Suggests Distinct Disease-Specific and Remodeling Signatures after Bariatric Surgery. Cell Metab. 2013, 18, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Meir, A.Y.; Keller, M.; Müller, L.; Bernhart, S.H.; Tsaban, G.; Zelicha, H.; Rinott, E.; Kaplan, A.; Gepner, Y.; Shelef, I.; et al. Effects of lifestyle interventions on epigenetic signatures of liver fat: CENTRAL randomized controlled trial. Liver Int. 2021, 41, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M.; Bray, G.A.; Carey, V.J.; Smith, S.R.; Ryan, D.H.; Anton, S.D.; McManus, K.; Champagne, C.M.; Bishop, L.M.; Laranjo, N.; et al. Comparison of Weight-Loss Diets with Different Compositions of Fat, Protein, and Carbohydrates. N. Engl. J. Med. 2009, 360, 859–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, B.; Irvin, M.R.; Sha, J.; Zhi, D.; Aslibekyan, S.; Absher, D.; Tiwari, H.K.; Kabagambe, E.K.; Ordovas, J.M.; Arnett, D.K. Epigenome-Wide Association Study of Fasting Measures of Glucose, Insulin, and HOMA-IR in the Genetics of Lipid Lowering Drugs and Diet Network Study. Diabetes 2014, 63, 801–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minn, A.H.; Hafele, C.; Shalev, A. Thioredoxin-Interacting Protein Is Stimulated by Glucose through a Carbohydrate Response Element and Induces β-Cell Apoptosis. Endocrinology 2005, 146, 2397–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkmar, M.; Dedeurwaerder, S.; A Cunha, D.; Ndlovu, M.N.; Defrance, M.; Deplus, R.; Calonne, E.; Volkmar, U.; Igoillo-Esteve, M.; Naamane, N.; et al. DNA methylation profiling identifies epigenetic dysregulation in pancreatic islets from type 2 diabetic patients. EMBO J. 2012, 31, 1405–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchi, M.; Lisi, S.; Curcio, M.; Barbuti, S.; Piaggi, P.; Ceccarini, G.; Nannipieri, M.; Anselmino, M.; Di Salvo, C.; Vitti, P.; et al. Human leptin tissue distribution, but not weight loss-dependent change in expression, is associated with methylation of its promoter. Epigenetics 2011, 6, 1198–1206. [Google Scholar] [CrossRef] [Green Version]
- Multhaup, M.L.; Seldin, M.M.; Jaffe, A.E.; Lei, X.; Kirchner, H.; Mondal, P.; Li, Y.; Rodriguez, V.; Drong, A.; Hussain, M.; et al. Mouse-human experimental epigenetic analysis unmasks dietary targets and genetic liability for diabetic phenotypes. Cell Metab. 2015, 21, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Romaine, S.P.R.; Tomaszewski, M.; Condorelli, G.; Samani, N.J. MicroRNAs in cardiovascular disease: An introduction for clinicians. Heart 2015, 101, 921–928. [Google Scholar] [CrossRef]
- Deng, J.; Guo, F. MicroRNAs and type 2 diabetes. ExRNA 2019, 1, 36. [Google Scholar] [CrossRef] [Green Version]
- Pordzik, J.; Jakubik, D.; Jarosz-Popek, J.; Wicik, Z.; Eyileten, C.; De Rosa, S.; Indolfi, C.; Siller-Matula, J.M.; Czajka, P.; Postula, M. Significance of circulating microRNAs in diabetes mellitus type 2 and platelet reactivity: Bioinformatic analysis and review. Cardiovasc. Diabetol. 2019, 18, 113. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zhang, C. MicroRNA-21 in Cardiovascular Disease. J. Cardiovasc. Transl. Res. 2010, 3, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayawardena, E.; Medzikovic, L.; Ruffenach, G.; Eghbali, M. Role of miRNA-1 and miRNA-21 in Acute Myocardial Ischemia-Reperfusion Injury and Their Potential as Therapeutic Strategy. Int. J. Mol. Sci. 2022, 23, 1512. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Cheng, Y.; Yue, J.; Yang, J.; Liu, X.; Chen, H.; Dean, D.B.; Zhang, C. MicroRNA Expression Signature and Antisense-Mediated Depletion Reveal an Essential Role of MicroRNA in Vascular Neointimal Lesion Formation. Circ. Res. 2007, 100, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C. MicroRNomics: A newly emerging approach for disease biology. Physiol. Genom. 2008, 33, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wang, B.; Zhou, Q.; Wang, Y.; Liu, X.; Liu, Z.; Zhan, Z. MicroRNA-21 prevents excessive inflammation and cardiac dysfunction after myocardial infarction through targeting KBTBD7. Cell Death Dis. 2018, 9, 769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, Y.; Vausort, M.; Goretti, E.; Nazarov, P.; Azuaje, F.; Gilson, G.; Corsten, M.F.; Schroen, B.; Lair, M.-L.; Heymans, S.; et al. Use of Circulating MicroRNAs to Diagnose Acute Myocardial Infarction. Clin. Chem. 2012, 58, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Leti, F.; DiStefano, J.K. Long Noncoding RNAs as Diagnostic and Therapeutic Targets in Type 2 Diabetes and Related Complications. Genes 2017, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Qu, J.; Gai, L.; Yuan, D.; Yuan, C. Long Non-coding RNAs in Metabolic and Inflammatory Pathways in Obesity. Curr. Pharm. Des. 2020, 26, 3317–3325. [Google Scholar] [CrossRef]
- Franco, D.; Aranega, A.; Dominguez, J.N. Non-coding RNAs and Atrial Fibrillation. Adv. Exp. Med. Biol. 2020, 1229, 311–325. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Taheri, M. The expression profile and role of non-coding RNAs in obesity. Eur. J. Pharmacol. 2020, 892, 173809. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasolo, F.; Di Gregoli, K.; Maegdefessel, L.; Johnson, J.L. Non-coding RNAs in cardiovascular cell biology and atherosclerosis. Cardiovasc. Res. 2019, 115, 1732–1756. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, C.; Prabu, P.; Mohan, V.; Balasubramanyam, M. Linking a role of lncRNAs (long non-coding RNAs) with insulin resistance, accelerated senescence, and inflammation in patients with type 2 diabetes. Hum. Genom. 2018, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Morán, I.; Akerman, I.; van de Bunt, M.; Xie, R.; Benazra, M.; Nammo, T.; Arnes, L.; Nakić, N.; García-Hurtado, J.; Rodriguez-Segui, S.A.; et al. Human β Cell Transcriptome Analysis Uncovers lncRNAs That Are Tissue-Specific, Dynamically Regulated, and Abnormally Expressed in Type 2 Diabetes. Cell Metab. 2012, 16, 435–448. [Google Scholar] [CrossRef]
- Fadista, J.; Vikman, P.; Laakso, E.O.; Mollet, I.G.; Esguerra, J.L.; Taneera, J.; Storm, P.; Osmark, P.; Ladenvall, C.; Prasad, R.B.; et al. Global genomic and transcriptomic analysis of human pancreatic islets reveals novel genes influencing glucose metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 13924–13929. [Google Scholar] [CrossRef] [Green Version]
- Akerman, I.; Tu, Z.; Beucher, A.; Rolando, D.M.; Sauty-Colace, C.; Benazra, M.; Nakic, N.; Yang, J.; Wang, H.; Pasquali, L.; et al. Human Pancreatic β Cell lncRNAs Control Cell-Specific Regulatory Networks. Cell Metab. 2017, 25, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Meerson, A.; Traurig, M.; Ossowski, V.; Fleming, J.M.; Mullins, M.; Baier, L.J. Human adipose microRNA-221 is upregulated in obesity and affects fat metabolism downstream of leptin and TNF-α. Diabetologia 2013, 56, 1971–1979. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.J.; Moreno-Navarrete, J.M.; Pardo, G.; Sabater-Masdeu, M.; Hummel, M.; Ferrer, A.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. MiRNA Expression Profile of Human Subcutaneous Adipose and during Adipocyte Differentiation. PLoS ONE 2010, 5, e9022. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sinnott-Armstrong, N.; Wagschal, A.; Wark, A.R.; Camporez, J.-P.; Perry, R.J.; Ji, F.; Sohn, Y.; Oh, J.; Wu, S.; et al. A MicroRNA Linking Human Positive Selection and Metabolic Disorders. Cell 2020, 183, 684–701.e14. [Google Scholar] [CrossRef]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.J.; Mercader, J.M.; Moreno-Navarrete, J.M.; Rovira, O.; Guerra, E.; Esteve, E.; Xifra, G.; Martínez, C.; Ricart, W.; Rieusset, J.; et al. Profiling of Circulating MicroRNAs Reveals Common MicroRNAs Linked to Type 2 Diabetes That Change With Insulin Sensitization. Diabetes Care 2014, 37, 1375–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayés-Genis, A.; Lanfear, D.E.; De Ronde, M.W.; Lupón, J.; Leenders, J.J.; Liu, Z.; Zuithoff, N.P.; Eijkemans, M.J.; Zamora, E.; De Antonio, M.; et al. Prognostic value of circulating microRNAs on heart failure-related morbidity and mortality in two large diverse cohorts of general heart failure patients. Eur. J. Heart Fail. 2017, 20, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melman, Y.F.; Shah, R.; Danielson, K.; Xiao, J.; Simonson, B.; Barth, A.; Chakir, K.; Lewis, G.D.; Lavender, Z.; Truong, Q.A.; et al. Circulating MicroRNA-30d Is Associated With Response to Cardiac Resynchronization Therapy in Heart Failure and Regulates Cardiomyocyte Apoptosis. Circulation 2015, 131, 2202–2216. [Google Scholar] [CrossRef] [PubMed]
- Van Boven, N.; Kardys, I.; Van Vark, L.C.; Akkerhuis, K.M.; De Ronde, M.W.J.; Khan, M.A.F.; Merkus, D.; Liu, Z.; Voors, A.A.; Asselbergs, F.; et al. Serially measured circulating microRNAs and adverse clinical outcomes in patients with acute heart failure. Eur. J. Heart Fail. 2018, 20, 89–96. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandra, Y.; Devanna, P.; Limana, F.; Straino, S.; Di Carlo, A.; Brambilla, P.G.; Rubino, M.; Carena, M.C.; Spazzafumo, L.; De Simone, M.; et al. Circulating microRNAs are new and sensitive biomarkers of myocardial infarction. Eur. Heart J. 2010, 31, 2765–2773. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zhang, Y.; Wang, N.; Pan, Z.; Gao, X.; Zhang, F.; Zhang, Y.; Shan, H.; Luo, X.; Bai, Y.; et al. MicroRNA-328 Contributes to Adverse Electrical Remodeling in Atrial Fibrillation. Circulation 2010, 122, 2378–2387. [Google Scholar] [CrossRef]
- Cannataro, R.; Perri, M.; Gallelli, L.; Caroleo, M.C.; De Sarro, G.; Cione, E. Ketogenic Diet Acts on Body Remodeling and MicroRNAs Expression Profile. MicroRNA 2019, 8, 116–126. [Google Scholar] [CrossRef]
- Wang, M.; Xue, Q.; Li, X.; Krohn, K.; Ziesche, S.; Ceglarek, U.; Blüher, M.; Keller, M.; Meir, A.Y.; Heianza, Y.; et al. Circulating Levels of microRNA-122 and Hepatic Fat Change in Response to Weight-Loss Interventions: CENTRAL Trial. J. Clin. Endocrinol. Metab. 2022, dgac023. [Google Scholar] [CrossRef]
- Assmann, T.S.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez, J.A. Circulating adiposity-related microRNAs as predictors of the response to a low-fat diet in subjects with obesity. J. Cell. Mol. Med. 2020, 24, 2956–2967. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Zhou, B.; Ross, S.A.; Zempleni, J. Nutrition, microRNAs, and Human Health. Adv. Nutr. 2017, 8, 105–112. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Disease | CpG Site | Gene | Gene Summary * |
|---|---|---|---|
| Obesity | cg00574958 †, cg09737197 †, cg17058475 †, cg01082498 † | CPT1A | CPT1A (Carnitine Palmitoyltransferase 1A) is a Protein Coding gene. Diseases associated with CPT1A include Carnitine Palmitoyltransferase I Deficiency and Carnitine Palmitoyltransferase Ii Deficiency, Infantile. Among its related pathways are Import of palmitoyl-CoA into the mitochondrial matrix and Regulation of lipid metabolism by Peroxisome proliferator-activated receptor alpha. Gene Ontology (GO) annotations related to this gene include identical protein binding and carnitine O-palmitoyltransferase activity. |
| cg06500161 | ABCG1 | ABCG1 (ATP Binding Cassette Subfamily G Member 1) is a Protein Coding gene. Diseases associated with ABCG1 include Tangier Disease and Sitosterolemia. Among its related pathways are Nuclear Receptors in Lipid Metabolism and Toxicity and Lipoprotein metabolism. GO annotations related to this gene include protein homodimerization activity and GTP binding. | |
| cg11024682 † | SREBF1 | SREBF1 (Sterol Regulatory Element Binding Transcription Factor 1) is a Protein Coding gene. Diseases associated with SREBF1 include Ifap Syndrome 2 and Mucoepithelial Dysplasia, Hereditary. Among its related pathways are ID signaling pathway and SREBF and miR33 in cholesterol and lipid homeostasis. GO annotations related to this gene include DNA-binding transcription factor activity and chromatin binding. | |
| cg22891070, cg27146050, cg16672562 | HIF3A | HIF3A (Hypoxia Inducible Factor 3 Subunit Alpha) is a Protein Coding gene. Diseases associated with HIF3A include Hypoxia. Among its related pathways are Hypoxic and oxygen homeostasis regulation of HIF-1-alpha and CDK-mediated phosphorylation and removal of Cdc6. GO annotations related to this gene include DNA-binding transcription factor activity and transcription coactivator activity. | |
| T2D | cg19693031 | TXNIP | TXNIP (Thioredoxin Interacting Protein) is a Protein Coding gene. Diseases associated with TXNIP include Leukostasis and Hyperglycemia. Among its related pathways are Nucleotide-binding domain, leucine rich repeat containing receptor signaling pathways and Innate Immune System. GO annotations related to this gene include ubiquitin protein ligase binding and enzyme inhibitor activity. |
| cg06500161 | ABCG1 | ABCG1 (ATP Binding Cassette Subfamily G Member 1) is a Protein Coding gene. Diseases associated with ABCG1 include Tangier Disease and Sitosterolemia. Among its related pathways are Nuclear Receptors in Lipid Metabolism and Toxicity and Lipoprotein metabolism. GO annotations related to this gene include protein homodimerization activity and GTP binding. | |
| cg11024682 | SREBF1 | SREBF1 (Sterol Regulatory Element Binding Transcription Factor 1) is a Protein Coding gene. Diseases associated with SREBF1 include Ifap Syndrome 2 and Mucoepithelial Dysplasia, Hereditary. Among its related pathways are ID signaling pathway and SREBF and miR33 in cholesterol and lipid homeostasis. GO annotations related to this gene include DNA-binding transcription factor activity and chromatin binding. | |
| cg00574958 † | CPT1A | CPT1A (Carnitine Palmitoyltransferase 1A) is a Protein Coding gene. Diseases associated with CPT1A include Carnitine Palmitoyltransferase I Deficiency and Carnitine Palmitoyltransferase Ii Deficiency, Infantile. Among its related pathways are Import of palmitoyl-CoA into the mitochondrial matrix and Regulation of lipid metabolism by Peroxisome proliferator-activated receptor alpha. GO annotations related to this gene include identical protein binding and carnitine O-palmitoyltransferase activity. | |
| CVD | cg12766383 | UBR4 | UBR4 (Ubiquitin Protein Ligase E3 Component N-Recognin 4) is a Protein Coding gene. Diseases associated with UBR4 include Retinoblastoma and Episodic Ataxia. Among its related pathways are Innate Immune System and Class I MHC mediated antigen processing and presentation. GO annotations related to this gene include binding and ubiquitin-protein transferase activity. |
| cg05820312 | TRAPPC9 | TRAPPC9 (Trafficking Protein Particle Complex Subunit 9) is a Protein Coding gene. Diseases associated with TRAPPC9 include Intellectual Disability-Obesity-Brain Malformations-Facial Dysmorphism Syndrome and Autosomal Recessive Non-Syndromic Intellectual Disability. Among its related pathways are Transport to the Golgi and subsequent modification and Vesicle-mediated transport. | |
| cg26470101 † | DLX2 | DLX2 (Distal-Less Homeobox 2) is a Protein Coding gene. Diseases associated with DLX2 include Axenfeld-Rieger Syndrome and Split-Hand/Foot Malformation 5. Among its related pathways are Sudden Infant Death Syndrome. Susceptibility Pathways and Preimplantation Embryo. GO annotations related to this gene include DNA-binding transcription factor activity and chromatin binding. | |
| cg07289306 † | MIR138−1 | MIR138-1 (MicroRNA 138-1) is an RNA Gene, and is affiliated with the miRNA class. Diseases associated with MIR138-1 include Oral Squamous Cell Carcinoma and Thyroid Cancer, Nonmedullary, 1. |
| Intervention | Population | Tissue | Methylation Sites/Method | Main Findings |
|---|---|---|---|---|
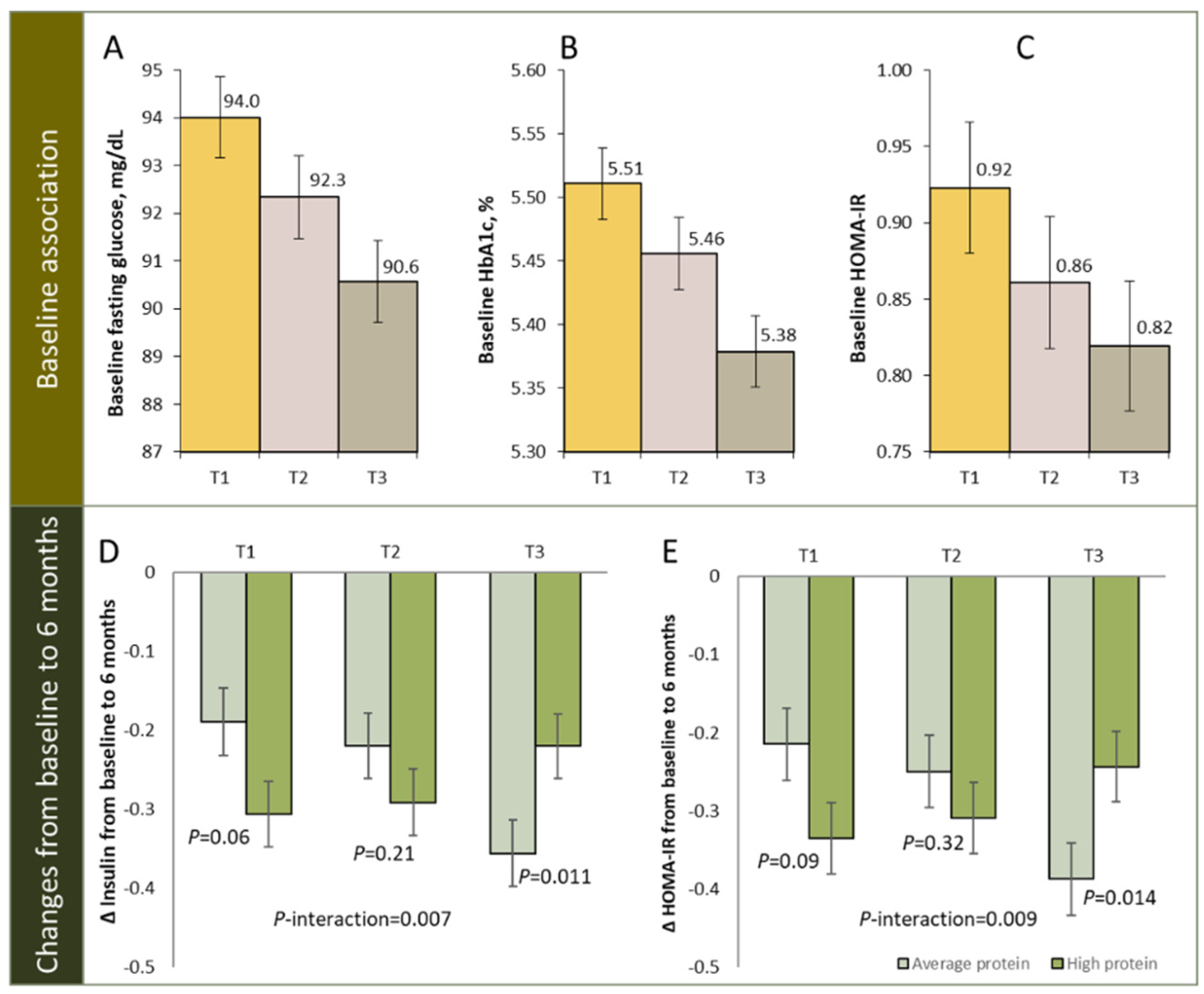
| 2-year diet interventions (4 energy-reduced diets with varying macronutrient compositions) [99] | 639 overweight/obese participants with available DNA methylation at baseline | Blood | Genome-wide/high-resolution methylC-capture sequencing | Among average-protein group, higher regional DNA methylation levels at TXNIP (average methylation level over CpG sites within 500 bp of cg19693031) was associated with greater reduction in insulin and HOMA-IR at 6 months |
| 18 months lifestyle intervention including diet and physical activity (Mediterranean low-carb (MED/LC) vs. low-fat (LF) vs. MED/LC + physical activity vs. LF + physical activity) [92] | A total of 120 sedentary adults with abdominal obesity or dyslipidemia (110 male and 10 female) | Blood | Genome-wide/Illumina HumanMethylation850 BeadChip | Differences were observed in 8 differentially methylated regions (DMRs) around 9 genes between 10 responders (mean weight change −16%) and 10 non-responders (+2.4%), including LRRC27, CRIPSP2, SLFN12, LINC00539, AURKC, RNF39, RP11-283I3.2, SLC6A12, NTSR1. Among all the 120 participants, 47 CpG sites (15 CpG sites correlated negatively and 32 positively) showed significant correlation with weight loss after intervention. |
| 18 months lifestyle intervention including diet and physical activity (Mediterranean low-carb (MED/LC) vs. low-fat (LF) vs. MED/LC + physical activity vs. LF + physical activity) [98] | A total of 120 sedentary adults with abdominal obesity or dyslipidemia (110 male and 10 female) | Blood | Genome-wide/Illumina HumanMethylation850 BeadChip | Lifestyle intervention attenuated DNA methylation age (mAge) of the men, particularly among the older participants. No difference between the mAging between the intervention groups. The 18-month changes in mAge among 39 weight-loss successors was significantly lower than that of the 81 weight-loss failures. Similarly, participants with healthy liver fat % (intrahepatic fat <5%, n = 75) at the end of the intervention had significantly lower mAge, compared to those participants with fatty liver (n = 45, intrahepatic fat >5% at the end of the intervention). |
| 6 months exercise intervention [93] | A total of 23 healthy overweight men | Adipose tissue | Genome-wide/Illumina HumanMethylation450 BeadChip | Global DNA methylation changed after exercise intervention. A total of 17,975 individual CpG sites in 7663 unique genes showed altered levels of DNA methylation. Also, 18 obesity and 21 T2D candidate genes had CpG sites with differences in adipose tissue DNA methylation, including TCF7L2 (6 CpG sites) and KCNQ1 (10 CpG sites). |
| 10 weeks multidisciplinary intensive lifestyle intervention [94] | First methylation array was conducted among 24 obese or overweight adolescents (12 high responders and 12 low responders); second, a validation analyses was performed in 107 adolescents | Blood | Genome-wide/Illumina Infinium HumanMethylation27 BeadChip27k, validation using Sequenom EpiTyper MassARRAY followed by MALDI-TOF mass spectrometry | Comparing the baseline differences between high responders and low responders revealed 97 CpG sites with >5% changes in DNA methylation, validation analysis showed 5 regions with differential methylation levels, including AQP9, DUSP22, NIPK3, TNNT1, and TNNI3. |
| 6 months caloric restriction intervention [80] | Overweight/obese postmenopausal women (7 high responders and 7 low responders) | Subcutaneous adipose tissue | Genome-wide/Human CpG-island 8.1 K array and 6800 additional CpG island loci, validation by Sequenom EpiTyper MassARRAY | At baseline, significant DNA methylation differences at 35 loci were found between the high and low responders before intervention. After intervention, 3 regions showed differential methylation. Some of the identified regions contains genes known to be related to weight control and insulin secretion, or in known imprinted genomic regions. |
| 6 months exercise intervention [95] | 15 men with (F+) and 13 men without (F-) a first-degree family history of T2D | Skeletal muscle | Genome-wide/Infinium HumanMethylation450 BeadChip, validation by Sequenom EpiTyper MassARRAY | A total of 134 individuals genes changed in DNA methylation level after intervention. The identified genes include those in the retinol metabolism and calcium signaling pathways and with known functions in muscle and T2D, such as MEF2A, RUNX1, NDUFC2, and THADA. |
| 1-year weight loss intervention (weight loss diet and exercise) [96] | 19 healthy obese participants | Subcutaneous adipose tissue | Genome-wide/Infinium HumanMethylation450 BeadChip | No genome-wide significant differentially methylated CpG sites were observed (baseline vs. 5 months, 5 months vs. 12 months, or baseline vs. 12 months) |
| RYGB [100] | Obese women with RYGB surgery (n = 8) and nonobese (normal weight) glucose-tolerant age-matched women (n = 9); obese men before and after RYGB surgery (n = 6) | Skeletal muscle | PGC1α and PDK4 promoter region and 14 other genes by bisulfite sequencing among women; Genome-wide methylation analysis for men by bisulfite sequencing | RYGB surgery decreased PGC-1α promoter methylation and conversely increased PDK4 promoter methylation. Among the 14 metabolic genes analyzed, promoter methylation of 11 genes was normalized to levels in normal weight individuals. In men, 409 DMRs were found pre- and post-RYGB. |
| Energy restriction diet and bariatric surgery [101] | Obese women with energy restriction diet (n = 22); obese women underwent a hypocaloric dietary treatment followed by RYGB (n = 14); normal weight women (n = 9) | Blood | Methylation at LINE-1, IL-6, and SERPINE-1 accessed by Methylation-Sensitive High-Resolution Melting (MS-HRM) | LINE-1 and SERPINE-1 methylation levels did not change after weight loss (energy restriction and RYBG), IL-6 methylation increased after energy restriction and decreased in the bariatric surgery group. Baseline SERPINE-1 methylation levels were significantly lower in the high responders to the RYBG. |
| Bariatric surgery [102] | 45 obese patients with NAFLD; 23 participants underwent bariatric surgery | Liver | Genome-wide/Illumina HumanMethylation450 BeadChip | Post-bariatric and NAFLD-specific methylation signatures were clearly distinct. The gene encoding protein-tyrosine phosphatase epsilon (PTPRE) showed a differential methylation before and after bariatric surgery. |
| MicroRNA ID | Changes in Expression | Associated Disease/Conditions | Ref. |
|---|---|---|---|
| miR-221 | ↑ | obesity/subcutaneous adipose tissue | [129,130] |
| miR-128-1 | ↑ | thrifty phenotype (high glucose, insulin resistance, energy storage, ets.) | [131] |
| miR-103/107 | ↑ | obesity, alcoholic liver disease (ALD), non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH) | [132] |
| miR-140-5p, miR-142-3p, miR-222 | ↑ | T2D | [133] |
| miR-423-5p, miR-125b, miR-192, miR-195, miR-130b, miR-532-5p, miR-126 | ↓ | ||
| miR-21 | ↑ | proliferative vascular disease, cardiac hypertrophy and heart failure, and ischemic heart disease. | [113,114,117] |
| miR-1254 | ↑ | chronic heart failure | [134] |
| miR-30d | ↓ | heart failure | [135] |
| miR-1306 | ↑ | acute heart failure | [136] |
| miR-1, miR-133a, miR-133b, miR-499-5p | ↑ | myocardial infarction | [137] |
| miR-122, miR-375 | ↓ | ||
| miR-223, miR-328, miR-664 | ↑ | atrial fibrillation | [138] |
| miR-101, miR-320, miR-499 | ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Qi, L. Epigenetics in Precision Nutrition. J. Pers. Med. 2022, 12, 533. https://doi.org/10.3390/jpm12040533
Li X, Qi L. Epigenetics in Precision Nutrition. Journal of Personalized Medicine. 2022; 12(4):533. https://doi.org/10.3390/jpm12040533
Chicago/Turabian StyleLi, Xiang, and Lu Qi. 2022. "Epigenetics in Precision Nutrition" Journal of Personalized Medicine 12, no. 4: 533. https://doi.org/10.3390/jpm12040533
APA StyleLi, X., & Qi, L. (2022). Epigenetics in Precision Nutrition. Journal of Personalized Medicine, 12(4), 533. https://doi.org/10.3390/jpm12040533