
Diaporine Potentiates the Anticancer Effects of Oxaliplatin and Doxorubicin on Liver Cancer Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Effect of Diaporine, Oxaliplatin, Doxorubicin and Their Combination on the Number of Liver Cancer Cells
2.3. Analysis of Drug Interactions
2.4. Effect of Different Treatments on Colony Growth in Matrigel Matrix
2.5. Quantification of Apoptosis
2.6. Effects of Diaporine, Oxaliplatin, Doxorubicin, and Their Combination on Expression and Phosphorylation of Proapoptotic and Antiproliferative Proteins
2.7. Statistics
3. Results
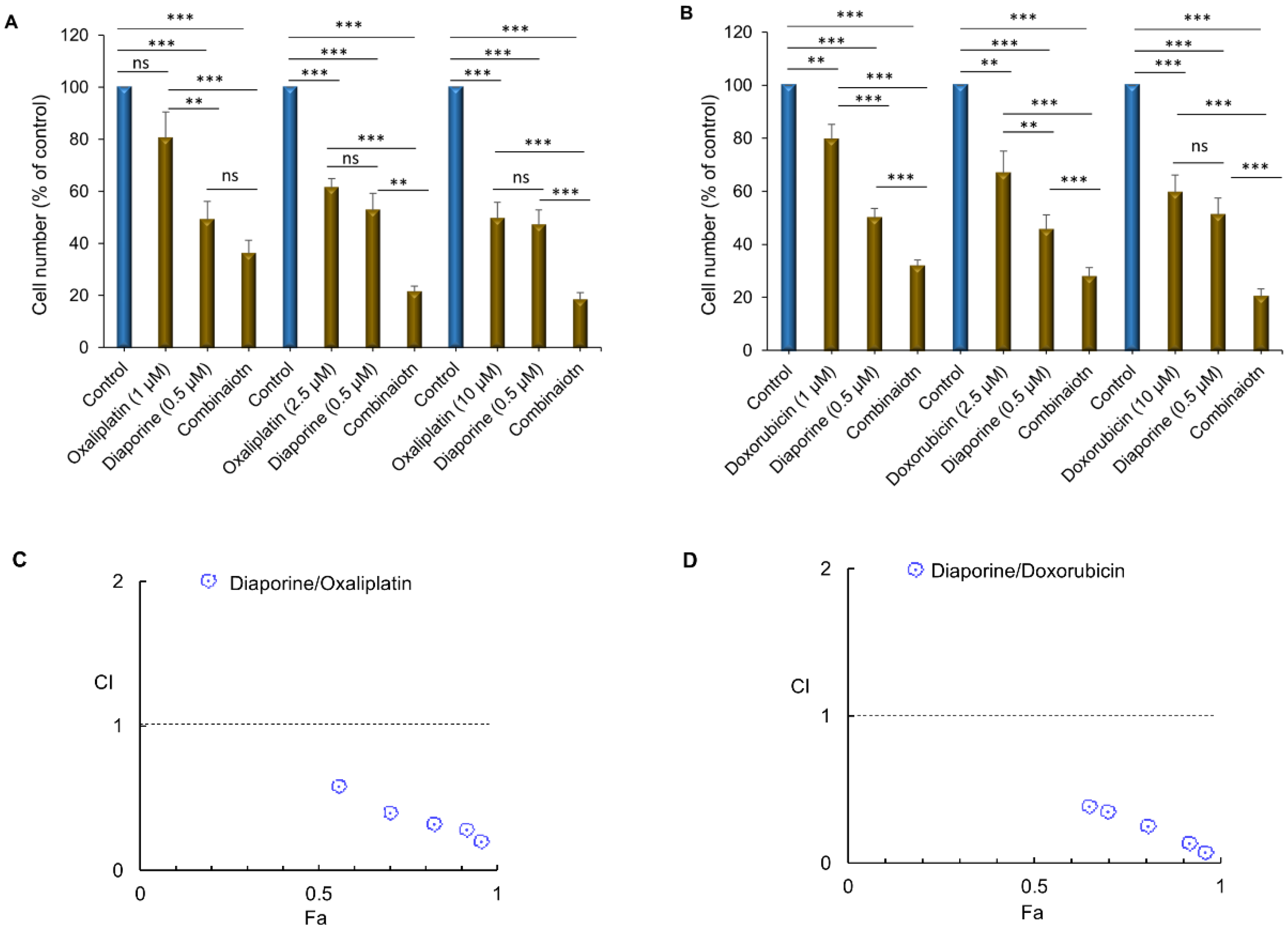
3.1. Effect of Diaporine, Oxaliplatin, and Doxorubicin on Cell Numbers
3.2. Diaporine Improved the Antitumor Activity of Oxaliplatin and Doxorubicin against HEPG2 Cells
3.3. Diaporine Combined with Doxorubicin or Oxaliplatin Promotes Apoptosis of HepG2 Hepatoma Cells
3.4. Effects of Diaporine, Oxaliplatin, and Doxorubicin Alone or in Combination on Survival, Apoptosis Pathway, and DNA Damage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, L.Q.; Lv, R.W.; Qu, X.D.; Chen, X.J.; Lu, H.S.; Wang, Y. Aloesin Suppresses Cell Growth and Metastasis in Ovarian Cancer SKOV3 Cells through the Inhibition of the MAPK Signaling Pathway. Anal. Cell. Pathol. 2017, 2017, 8158254. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Gu, J.; Lim, L.Y.; Yuan, Z.X.; Mo, J. Nanomedicine-Mediated Therapies to Target Breast Cancer Stem Cells. Front. Pharmacol. 2016, 7, 313. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Huang, G.; Chen, Z.; Zhang, Y. Nanomaterials in Targeting Cancer Stem Cells for Cancer Therapy. Front. Pharmacol. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Altekruse, S.F.; McGlynn, K.A.; Reichman, M.E. Hepatocellular carcinoma incidence, mortality, and survival trends in the United States from 1975 to 2005. J. Clin. Oncol. 2009, 27, 1485–1491. [Google Scholar] [CrossRef]
- Ikeda, M.; Morizane, C.; Ueno, M.; Okusaka, T.; Ishii, H.; Furuse, J. Chemotherapy for hepatocellular carcinoma: Current status and future perspectives. Jpn. J. Clin. Oncol. 2018, 48, 103–114. [Google Scholar] [CrossRef]
- Llovet, J.M.; Schwartz, M.; Mazzaferro, V. Resection and liver transplantation for hepatocellular carcinoma. Semin. Liver Dis. 2005, 25, 181–200. [Google Scholar] [CrossRef]
- Patra, C.R.; Mukherjee, S.; Kotcherlakota, R. Biosynthesized silver nanoparticles: A step forward for cancer theranostics? Nanomedicine 2014, 9, 1445–1448. [Google Scholar] [CrossRef]
- Mukherjee, S.; Patra, C.R. Therapeutic application of anti-angiogenic nanomaterials in cancers. Nanoscale 2016, 8, 12444–12470. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod. Rep. 2000, 17, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Zhang, Y.L.; Lin, F.C.; Hu, Y.Z.; Zhang, C.L. Natural products with antitumor activity from endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 1056–1074. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.H.; Zhang, W.Y.; Ding, G.; Saparpakorn, P.; Song, Y.C.; Hannongbua, S.; Tan, R.X. Chaetoglobins A and B, two unusual alkaloids from endophytic Chaetomium globosum culture. Chem. Commun. 2008, 45, 5978–5980. [Google Scholar] [CrossRef]
- Wu, H.C.; Ge, H.M.; Zang, L.Y.; Bei, Y.C.; Niu, Z.Y.; Wei, W.; Feng, X.J.; Dind, S.; Ng, S.W.; Shen, P.P.; et al. Diaporine, a novel endophyte-derived regulator of macrophage differentiation. Org. Biomol. Chem. 2014, 12, 6545–6548. [Google Scholar] [CrossRef]
- Song, Y.; Dou, H.; Wang, P.; Zhao, S.; Wang, T.; Gong, W.; Zhao, J.; Li, E.; Tan, R.; Hou, Y. A novel small-molecule compound diaporine A inhibits non-small cell lung cancer growth by regulating miR-99a/mTOR signaling. Cancer Biol. Ther. 2014, 15, 1423–1430. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Sebolt-Leopold, J.S.; Dudley, D.T.; Herrera, R.; Van Becelaere, K.; Wiland, A.; Gowan, R.C.; Tecle, H.; Barrett, S.D.; Bridges, A.; Przybranowski, S.; et al. Blockade of the MAP kinase pathway suppresses growth of colon tumors in vivo. Nat. Med. 1999, 5, 810–816. [Google Scholar] [CrossRef]
- Moon, D.O.; Park, S.Y.; Choi, Y.H.; Kim, N.D.; Lee, C.; Kim, G.Y. Melittin induces Bcl-2 and caspase-3-dependent apoptosis through downregulation of Akt phosphorylation in human leukemic U937 cells. Toxicon 2008, 51, 112–120. [Google Scholar] [CrossRef]
- Weng, H.Y.; Hsu, M.J.; Chen, C.C.; Chen, B.C.; Hong, C.Y.; Teng, C.M.; Pan, S.L.; Chiu, W.T.; Lin, C.H. Denbinobin induces human glioblastoma multiforme cell apoptosis through the IKKalpha-Akt-FKHR signaling cascade. Eur. J. Pharmacol. 2013, 698, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Shiragami, R.; Murata, S.; Kosugi, C.; Tezuka, T.; Yamazaki, M.; Hirano, A.; Yoshimura, Y.; Suzuki, M.; Shuto, K.; Koda, K. Enhanced antitumor activity of cerulenin combined with oxaliplatin in human colon cancer cells. Int. J. Oncol. 2013, 43, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Dale Rein, I.; Solberg Landsverk, K.; Micci, F.; Patzke, S.; Stokke, T. Replication-induced DNA damage after PARP inhibition causes G2 delay, and cell line-dependent apoptosis, necrosis and multinucleation. Cell Cycle 2015, 14, 3248–3260. [Google Scholar] [CrossRef]
- Lee, T.K.; Lau, T.C.; Ng, I.O. Doxorubicin-induced apoptosis and chemosensitivity in hepatoma cell lines. Cancer Chemother. Pharmacol. 2002, 49, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; van der Zanden, S.Y.; Wander, D.P.A.; Borràs, D.M.; Song, J.-Y.; Li, X.; van Duikeren, S.; van Gils, N.; Rutten, A.; van Herwaarden, T.; et al. Uncoupling DNA damage from chromatin damage to detoxify doxorubicin. Proc. Natl. Acad. Sci. USA 2020, 117, 15182–15192. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Franklin, R.A.; Abrams, S.L.; Chappell, W.H.; Wong, E.W.; Lehmann, B.; Terrian, D.M.; Basecke, J.; Stivala, F.; et al. Targeting the RAF/MEK/ERK, PI3K/AKT and p53 pathways in hematopoietic drug resistance. Adv. Enzym. Regul. 2007, 47, 64–103. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Yang, L.; Cui, J.; Che, S.; Liu, Y.; Han, J.; An, X.; Cao, B.; Song, Y. miR-34a/c induce caprine endometrial epithelial cell apoptosis by regulating circ-8073/CEP55 via the RAS/RAF/MEK/ERK and PI3K/AKT/mTOR pathways. J. Cell Physiol. 2020, 235, 10051–10067. [Google Scholar] [CrossRef]
- Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Kempf, C.R.; Long, J.; Laidler, P.; Mijatovic, S.; Maksimovic-Ivanic, D.; Stivala, F.; Mazzarino, M.C.; et al. Roles of the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity to therapy-implications for cancer and aging. Aging 2011, 3, 192–222. [Google Scholar] [CrossRef]
- De Luca, A.; Maiello, M.R.; D’Alessio, A.; Pergameno, M.; Normanno, N. The RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: Role in cancer pathogenesis and implications for therapeutic approaches. Expert Opin. Ther. Targets 2012, 16 (Suppl. 2), S17–S27. [Google Scholar] [CrossRef]
- Steelman, L.S.; Pohnert, S.C.; Shelton, J.G.; Franklin, R.A.; Bertrand, F.E.; McCubrey, J.A. JAK/STAT, Raf/MEK/ERK, PI3K/Akt and BCR-ABL in cell cycle progression and leukemogenesis. Leukemia 2004, 18, 189–218. [Google Scholar] [CrossRef]
- Won, J.K.; Yang, H.W.; Shin, S.Y.; Lee, J.H.; Heo, W.D.; Cho, K.H. The crossregulation between ERK and PI3K signaling pathways determines the tumoricidal efficacy of MEK inhibitor. J. Mol. Cell. Biol. 2012, 4, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Dworakowska, D.; Wlodek, E.; Leontiou, C.A.; Igreja, S.; Çakır, M.; Teng, M.; Prodromou, N.; Góth, M.I.; Grozinsky-Glasberg, S.; Gueorguiev, M.; et al. Activation of RAF/MEK/ERK and PI3K/AKT/mTOR pathways in pituitary adenomas and their effects on downstream effectors. Endocr. Relat. Cancer 2009, 16, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Farooqi, A.A.; Ismail, M. Recent progress in fungus-derived bioactive agents for targeting of signaling machinery in cancer cells. Drug Des. Dev. Ther. 2015, 9, 1797–1804. [Google Scholar]
- Deng, Q.; Yu, X.; Xiao, L.; Hu, Z.; Luo, X.; Tao, Y.; Yang, L.; Liu, X.; Chen, H.; Ding, Z.; et al. Neoalbaconol induces energy depletion and multiple cell death in cancer cells by targeting PDK1-PI3-K/Akt signaling pathway. Cell Death Dis. 2013, 4, e804. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Himaya, S.W.; Dewapriya, P.; Zhang, C.; Kim, S.K. Fumigaclavine C from a marine-derived fungus Aspergillus fumigatus induces apoptosis in MCF-7 breast cancer cells. Mar. Drugs 2013, 11, 5063–5086. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Huang, J.J.; Cheung, P.C. Extract of Pleurotus pulmonarius suppresses liver cancer development and progression through inhibition of VEGF-induced PI3K/AKT signaling pathway. PLoS ONE 2012, 7, e34406. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Yu, W.; Zhou, F.; Chen, J.; Shen, P. A novel small molecule compound diaporine inhibits breast cancer cell proliferation via promoting ROS generation. Biomed. Pharmacother. 2016, 83, 1038–1047. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug A | Dose A (µM) | Drug B | Dose-B (µM) | Effect (Fa) | CI Value | Interaction |
|---|---|---|---|---|---|---|
| Diaporine | 0.5 | Oxaliplatin | 1.0 | 0.5624 | 0.56 | Synergism |
| 2.5 | 0.7032 | 0.39 | Synergism | |||
| 10.0 | 0.8211 | 0.31 | Synergism | |||
| 50.0 | 0.9134 | 0.26 | Synergism | |||
| 100.0 | 0.9512 | 0.18 | Synergism | |||
| Diaporine | 0.5 | Doxorubicin | 1.0 | 0.6512 | 0.39 | Synergism |
| 2.5 | 0.7018 | 0.35 | Synergism | |||
| 10.0 | 0.8011 | 0.25 | Synergism | |||
| 50.0 | 0.9023 | 0.17 | Synergism | |||
| 100.0 | 0.9612 | 0.06 | Synergism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, S.; Su, R.; Wu, K.; Zhou, X.; Vadgama, J.V.; Wu, Y. Diaporine Potentiates the Anticancer Effects of Oxaliplatin and Doxorubicin on Liver Cancer Cells. J. Pers. Med. 2022, 12, 1318. https://doi.org/10.3390/jpm12081318
Tian S, Su R, Wu K, Zhou X, Vadgama JV, Wu Y. Diaporine Potentiates the Anticancer Effects of Oxaliplatin and Doxorubicin on Liver Cancer Cells. Journal of Personalized Medicine. 2022; 12(8):1318. https://doi.org/10.3390/jpm12081318
Chicago/Turabian StyleTian, Shiliu, Rui Su, Ke Wu, Xuhan Zhou, Jaydutt V. Vadgama, and Yong Wu. 2022. "Diaporine Potentiates the Anticancer Effects of Oxaliplatin and Doxorubicin on Liver Cancer Cells" Journal of Personalized Medicine 12, no. 8: 1318. https://doi.org/10.3390/jpm12081318