
Shortened Daily Photoperiod Alleviates Anxiety-like Behaviour by Antioxidant Effect and Changes Serum Fatty Acid Profile in Diabetic Rats
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Experimental Design
2.1. Experimental Animals
2.2. Behavioural Testing
2.3. Melatonin Determination
2.4. Oxidative Stress
2.5. Lipid Status and FAME Determination
2.6. Statistical Analysis
3. Results
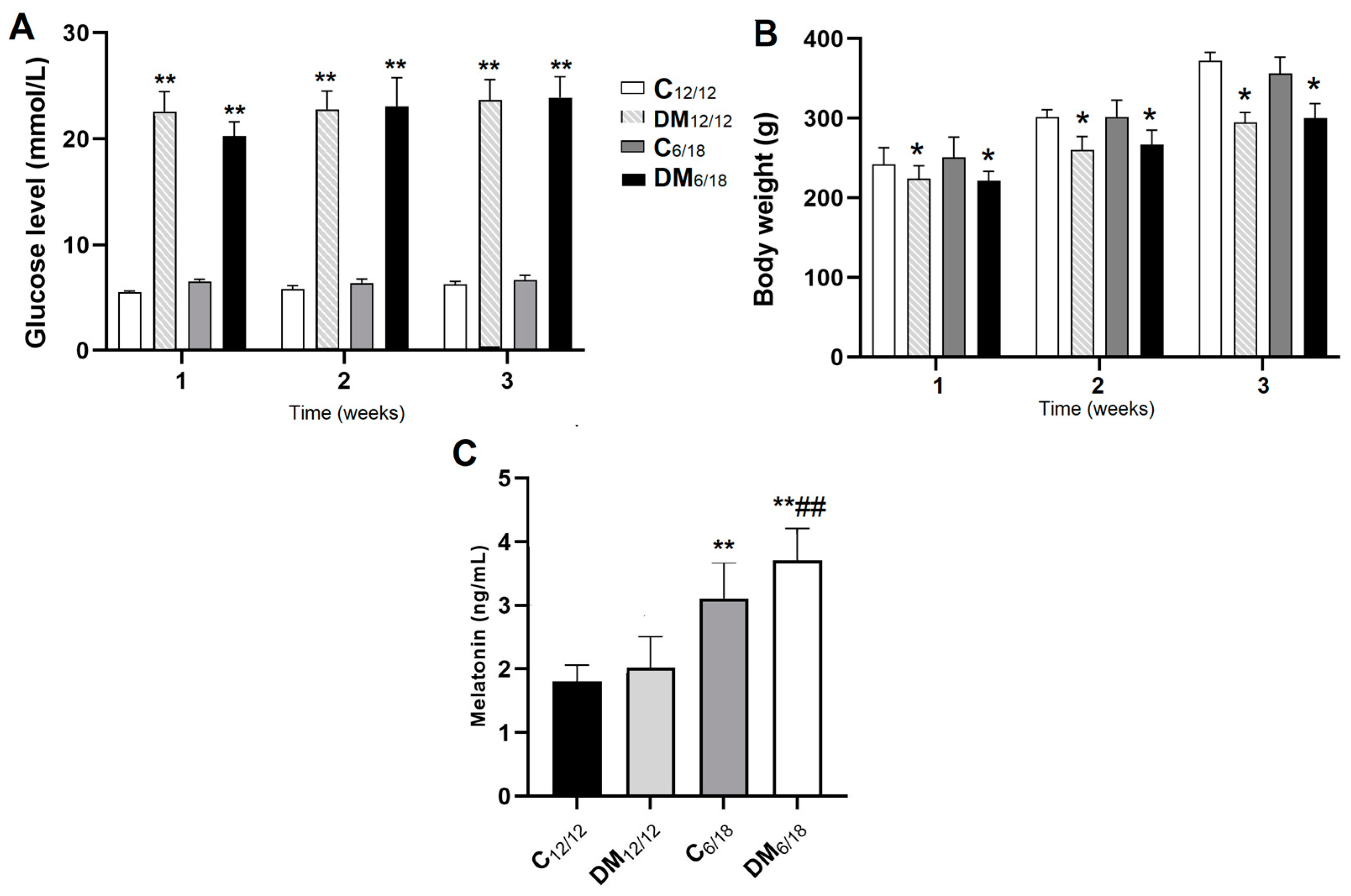
3.1. Blood Glucose and Melatonin Level
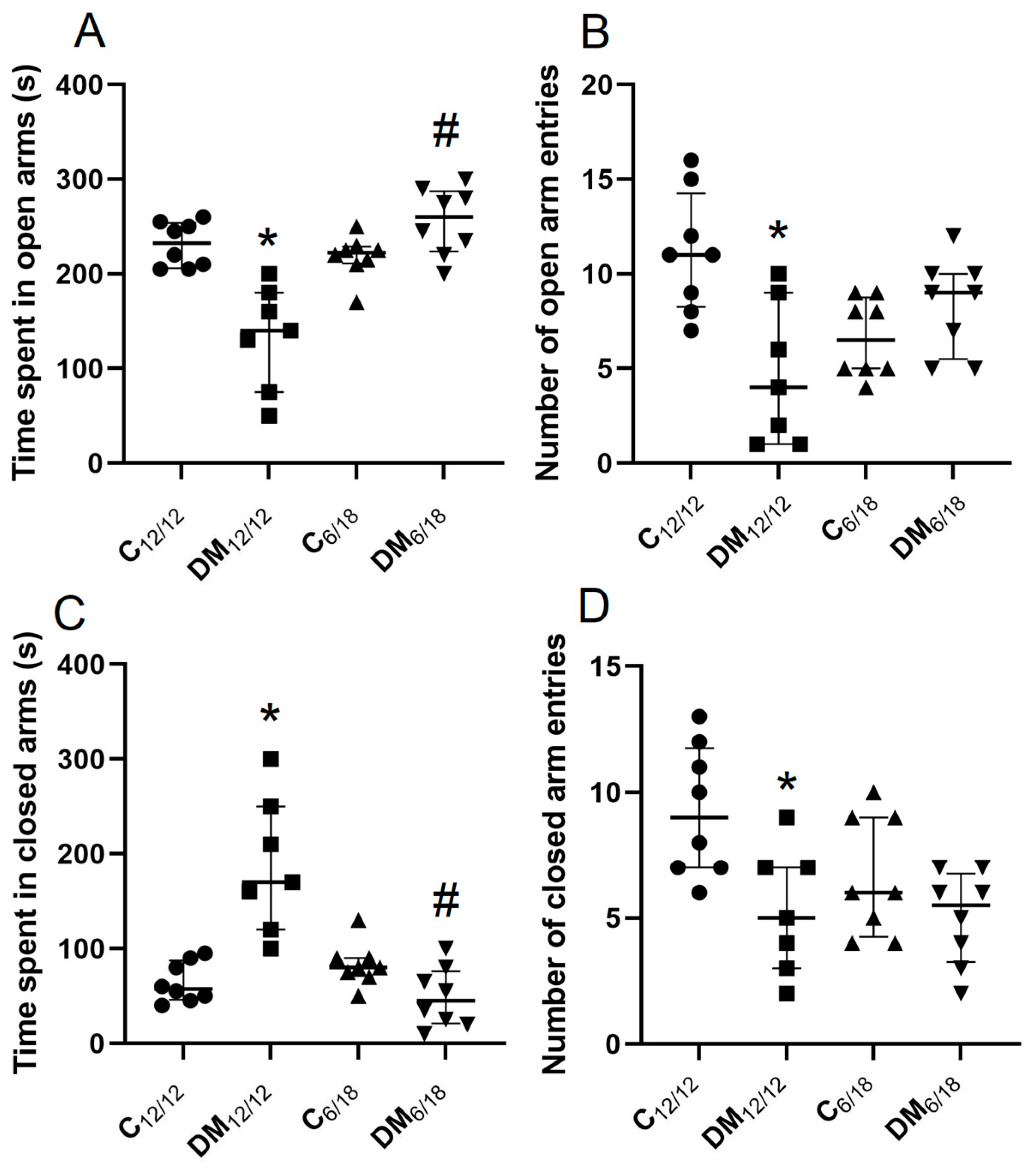
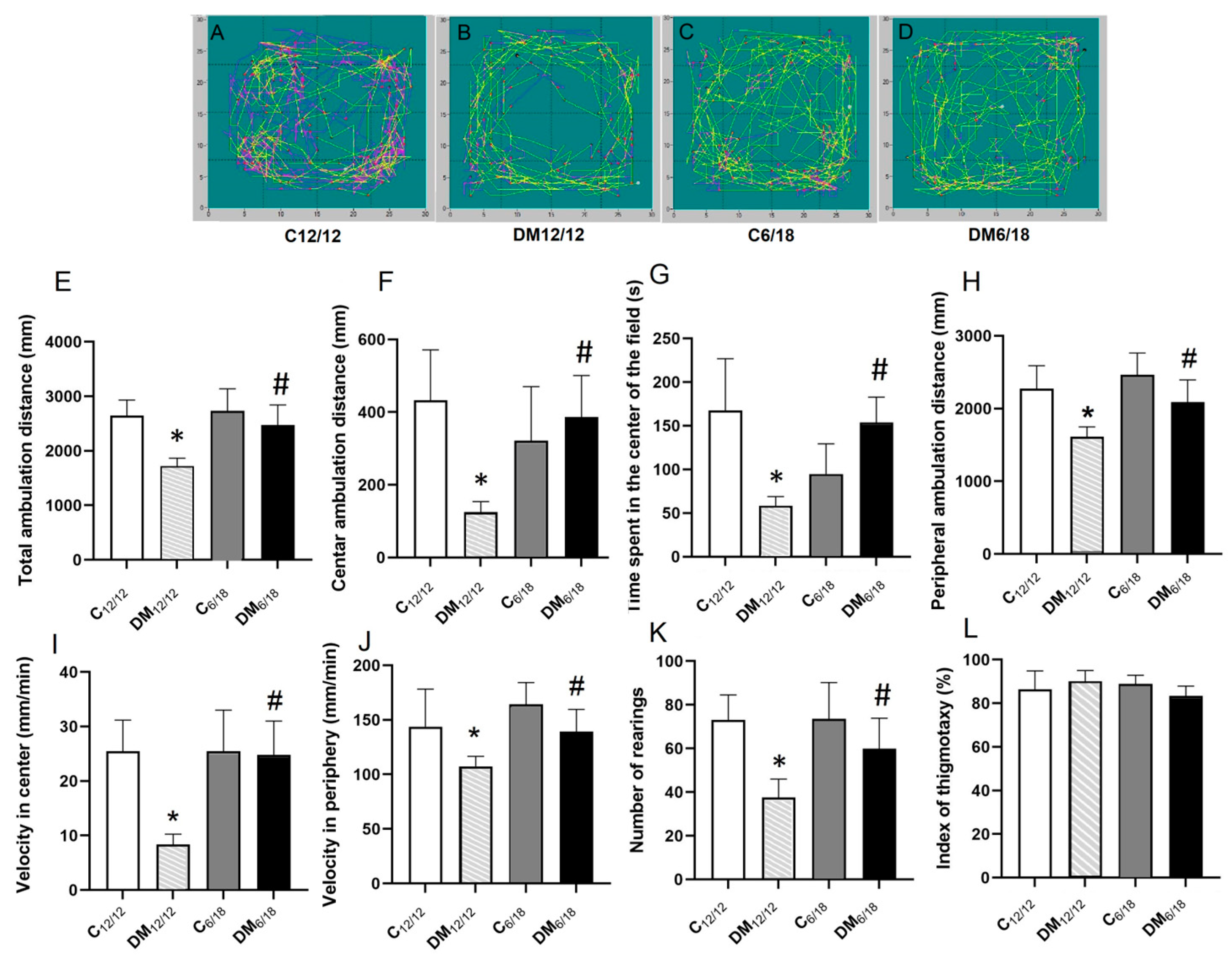
3.2. Behavioural Testing
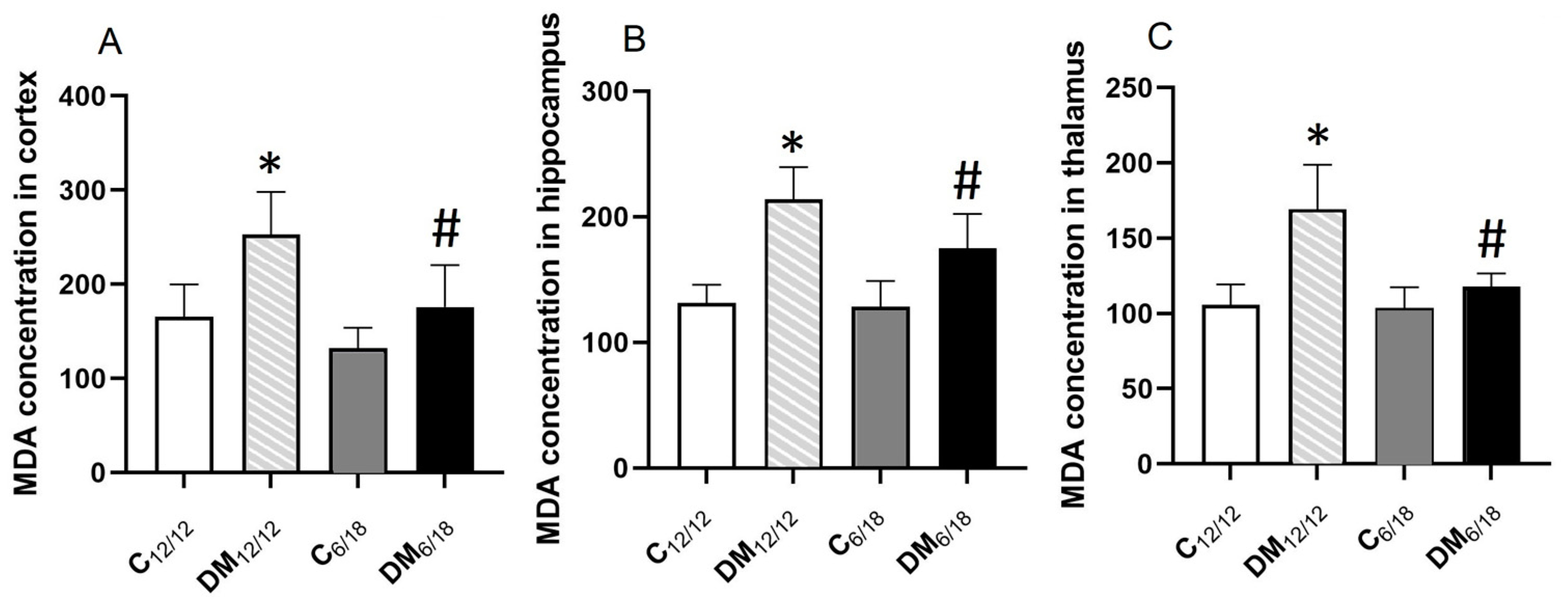
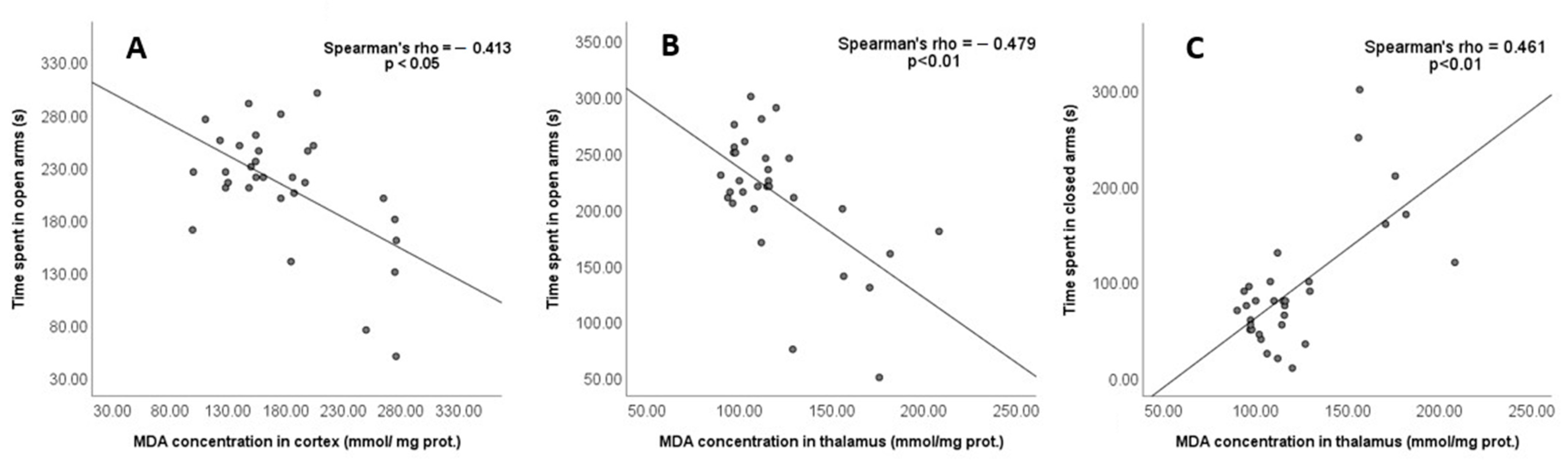
3.3. Oxidative Stress
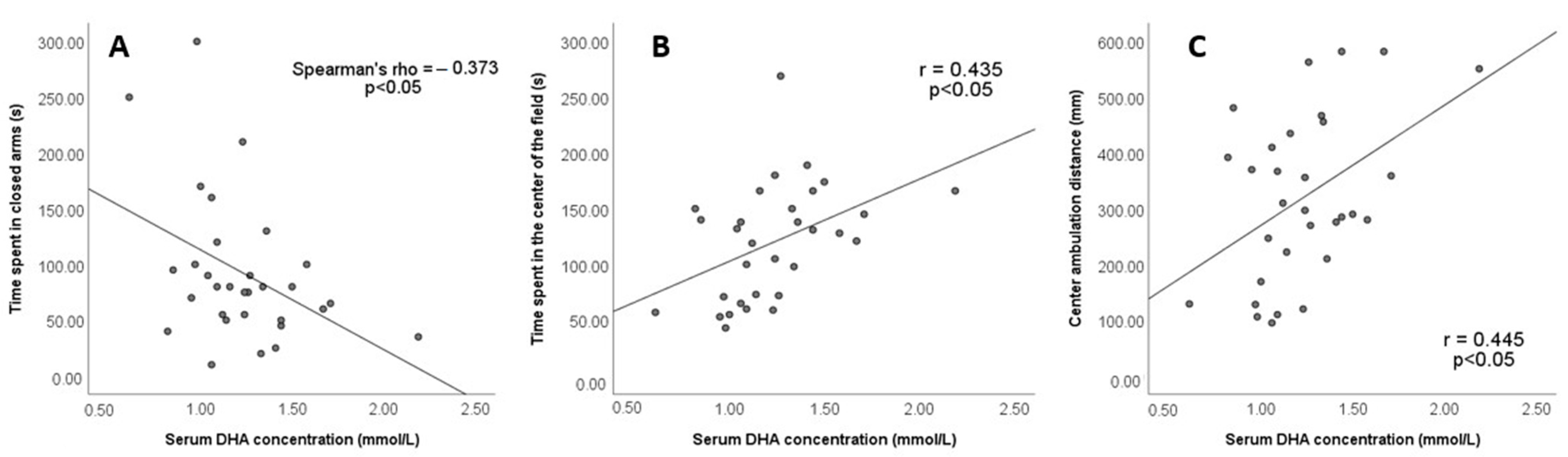
3.4. Lipid Status and FAMEs Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Indian J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef]
- OuYang, H.; Chen, B.; Abdulrahman, A.M.; Li, L.; Wu, N. Associations between Gestational Diabetes and Anxiety or Depression: A Systematic Review. J. Diabetes. Res. 2021, 27, 9959779. [Google Scholar] [CrossRef] [PubMed]
- Damanik, J.; Yunir, E. Type 2 Diabetes Mellitus and Cognitive Impairment. Acta Med. Indones. 2021, 53, 213–220. [Google Scholar] [PubMed]
- Bouayed, J.; Rammal, H.; Soulimani, R. Oxidative stress and anxiety: Relationship and cellular pathways. Oxid. Med. Cell. Longev. 2009, 2, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Fedoce, A.D.G.; Ferreira, F.; Bota, R.G.; Bonet-Costa, V.; Sun, P.Y.; Davies, K.J.A. The role of oxidative stress in anxiety disorder: Cause or consequence? Free Radic. Res. 2018, 52, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharmacol. 2019, 70, 809–824. [Google Scholar]
- Dang, R.; Wang, M.; Li, X.; Wang, H.; Liu, L.; Wu, Q.; Zhao, J.; Ji, P.; Zhong, L.; Licinio, J.; et al. Edaravone ameliorates depressive and anxiety-like behaviors via Sirt1/Nrf2/HO-1/Gpx4 pathway. J. Neuroinflamm. 2022, 19, 41. [Google Scholar] [CrossRef]
- Vastegani, S.M.; Khoshnam, S.E.; Mansouri, E.; Hajipour, S.; Ghafouri, S.; Bakhtiari, N.; Sarkaki, A.; Farbood, Y. Neuroprotective effect of anethole against rotenone induced non-motor deficits and oxidative stress in rat model of Parkinson’s disease. Behav. Brain. Res. 2023, 437, 114100. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Q.; Huang, H.; Liu, Z. The efficacy and acceptability of curcumin for the treatment of depression or depressive symptoms: A systematic review and meta-analysis. J. Affect. Disord. 2021, 282, 242–251. [Google Scholar] [CrossRef]
- Lama, A.; Pirozzi, C.; Severi, I.; Morgese, M.G.; Senzacqua, M.; Annunziata, C.; Comella, F.; Del Piano, F.; Schiavone, S.; Petrosino, S.; et al. Palmitoylethanolamide dampens neuroinflammation and anxiety-like behavior in obese mice. Brain Behav. Immun. 2022, 102, 110–123. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Andridge, R.; Malarkey, W.B.; Glaser, R. Omega-3 supplementation lowers inflammation and anxiety in medical students: A randomized controlled trial. Brain Behav. Immun. 2011, 25, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Jacka, F.N.; Pasco, J.A.; Williams, L.J.; Meyer, B.J.; Digger, R.; Berk, M. Dietary intake of fish and PUFA, and clinical depressive and anxiety disorders in women. Br. J. Nutr. 2013, 109, 2059–2066. [Google Scholar] [CrossRef]
- Cardoso, C.; Afonso, C.; Bandarra, N.M. Dietary DHA and health: Cognitive function ageing. Nutr. Res. Rev. 2016, 29, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H.; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [PubMed]
- Mason, I.C.; Qian, J.; Adler, G.K.; Scheer, F.A.J.L. Impact of circadian disruption on glucose metabolism: Implications for type 2 diabetes. Diabetologia 2020, 63, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Morris, C.J.; Scheer, F.A.J.L. Circadian misalignment increases mood vulnerability in simulated shift work. Sci. Rep. 2020, 10, 18614. [Google Scholar] [CrossRef] [PubMed]
- Ketchesin, K.D.; Becker-Krail, D.; McClung, C.A. Mood-related central and peripheral clocks. Eur. J. Neurosci. 2020, 51, 326–345. [Google Scholar] [CrossRef]
- Farhadi, N.; Gharghani, M.; Farhadi, Z. Effects of long-term light, darkness and oral administration of melatonin on serum levels of melatonin. Biomed. J. 2016, 39, 81–84. [Google Scholar] [CrossRef]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin ameliorates anxiety-like behaviors induced by sleep deprivation in mice: Role of oxidative stress, neuroinflammation, autophagy and apoptosis. Brain Res. Bull. 2021, 174, 161–172. [Google Scholar] [CrossRef]
- Oliveira-Abreu, K.; Cipolla-Neto, J.; Leal-Cardoso, J.H. Effects of Melatonin on Diabetic Neuropathy and Retinopathy. Int. J. Mol. Sci. 2021, 23, 100. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, A.; Ohta, Y.; Ohashi, K. Melatonin improves metabolic syndrome induced by high fructose intake in rats. J. Pineal. Res. 2012, 52, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Kondo, Y.; Nakatani, A.; Hayashi, K.; Naruse, A. Characterization of low dose streptozotocin-induced progressive diabetes in mice. Environ. Toxicol. Pharmacol. 2001, 9, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Sohn, S.; Lee, M.; Jung, J.; Kineman, R.D.; Park, S. Differential responses of the growth hormone axis in two rat models of streptozotocin-induced insulinopenic diabetes. J. Endocrinol. 2006, 188, 263–270. [Google Scholar] [CrossRef]
- Chen, V.; Ianuzzo, C.D. Dosage effect of streptozotocin on rat tissue enzyme activities and glycogen concentration. Can. J. Physiol. Pharmacol. 1982, 60, 1251–1256. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5.47.1–5.47.20. [Google Scholar] [CrossRef]
- Haller, J.; Aliczki, M.; Gyimesine Pelczer, K. Classical and novel approaches to the preclinical testing of anxiolytics: A critical evaluation. Neurosci. Biobehav. Rev. 2013, 37, 2318–2330. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Hrncic, D.; Mikić, J.; Rasic-Markovic, A.; Velimirović, M.; Stojković, T.; Obrenović, R.; Rankov-Petrović, B.; Šušić, V.; Djuric, D.; Petronijević, N.; et al. Anxiety-related behavior in hyperhomocysteinemia induced by methionine nutritional overload in rats: Role of the brain oxidative stress. Can. J. Physiol. Pharmacol. 2016, 94, 1074–1082. [Google Scholar] [CrossRef]
- Ramos, A.; Correia, E.C.; Izídio, G.S.; Brüske, G.R. Genetic selection of two new rat lines displaying different levels of anxiety-related behaviors. Behav. Genet. 2003, 33, 657–668. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [PubMed]
- Jocelyn, P.C. Spectrophotometric assay of thiols. Methods Enzymol. 1987, 143, 44–67. [Google Scholar] [PubMed]
- Sugiuchi, H.; Uji, Y.; Okabe, H.; Irie, T.; Uekama, K.; Kayahara, N.; Miyauchi, K. Direct measurement of high-density lipoprotein cholesterol in serum with polyethylene glycol-modified enzymes and sulfated alpha-cyclodextrin. Clin. Chem. 1995, 41, 717–723. [Google Scholar] [CrossRef]
- Wahlefeld, A.W. Determination of Serum Triglycerides Level. In Methods of Enzymatic Analysis; Bergmeyer, H., Ed.; Academic Press: New York, NY, USA, 1974; pp. 1831–1835. [Google Scholar]
- Schlechtriem, C.; Henderson, J.R.; Tocher, D.R. A critical assessment of different transmethylation procedures commonly employed in the fatty acid analysis of aquatic organisms. Limnol. Oceanogr. Methods 2008, 6, 523–531. [Google Scholar] [CrossRef]
- Zhou, X.; Gan, T.; Fang, G.; Wang, S.; Mao, Y.; Ying, C. Zeaxanthin improved diabetes-induced anxiety and depression through inhibiting inflammation in hippocampus. Metab. Brain Dis. 2018, 33, 705–711. [Google Scholar]
- Zhu, X.; Zhang, Y.M.; Zhang, M.Y.; Chen, Y.J.; Liu, Y.W. Hesperetin ameliorates diabetes-associated anxiety and depression-like behaviors in rats via activating Nrf2/ARE pathway. Metab. Brain Dis. 2021, 36, 1969–1983. [Google Scholar] [CrossRef]
- Buchberger, B.; Huppertz, H.; Krabbe, L.; Lux, B.; Mattivi, J.T.; Siafarikas, A. Symptoms of depression and anxiety in youth with type 1 diabetes: A systematic review and meta-analysis. Psychoneuroendocrinology 2016, 70, 70–84. [Google Scholar]
- Wang, Y.; Wang, S.; Xin, Y.; Zhang, J.; Wang, S.; Yang, Z.; Liu, C. Hydrogen sulfide alleviates the anxiety-like and depressive-like behaviors of type 1 diabetic mice via inhibiting inflammation and ferroptosis. Life Sci. 2021, 1, 278. [Google Scholar] [CrossRef]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, C.; Klabnik, J.J.; O’Donnell, J.M. Novel therapeutic targets in depression and anxiety: Antioxidants as a candidate treatment. Curr. Neuropharmacol. 2014, 12, 108–119. [Google Scholar] [CrossRef]
- Souza, C.G.; Moreira, J.D.; Siqueira, I.R.; Pereira, A.G.; Rieger, D.K.; Souza, D.O.; Souza, T.M.; Portela, L.V.; Perry, M.L. Highly palatable diet consumption increases protein oxidation in rat frontal cortex and anxiety-like behavior. Life Sci. 2007, 81, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Desrumaux, C.; Risold, P.Y.; Schroeder, H.; Deckert, V.; Masson, D.; Athias, A.; Laplanche, H.; Le Guern, N.; Blache, D.; Jiang, X.C.; et al. Phospholipid transfer protein (PLTP) deficiency reduces brain vitamin E content and increases anxiety in mice. FASEB J. 2005, 19, 296–297. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Capone, F.; Giorgio, M.; Pelicci, P.G.; de Kloet, E.R.; Alleva, E.; Minghetti, L.; Cirulli, F. Deletion of the life span determinant p66Shc prevents age-dependent increases in emotionality and pain sensitivity in mice. Exp. Gerontol. 2007, 42, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Mladenović, D.; Petronijević, N.; Stojković, T.; Velimirović, M.; Jevtić, G.; Hrnčić, D.; Radosavljević, T.; Rašić-Marković, A.; Maksić, N.; Djuric, D.; et al. Finasteride Has Regionally Different Effects on Brain Oxidative Stress and Acetylcholinesterase Activity in Acute Thioacetamide-Induced Hepatic Encephalopathy in Rats. PLoS ONE 2015, 10, e0134434. [Google Scholar] [CrossRef]
- Veskovic, M.; Mladenovic, D.; Jorgacevic, B.; Stevanovic, I.; de Luka, S.; Radosavljevic, T. Alpha-lipoic acid affects the oxidative stress in various brain structures in mice with methionine and choline deficiency. Exp. Biol. Med. 2015, 240, 418–425. [Google Scholar] [CrossRef]
- Shivakumar, B.R.; Anandatheerthavarada, H.K.; Ravindranath, V. Free radical scavenging systems in developing rat brain. Int. J. Dev. Neurosci. 1991, 9, 181–185. [Google Scholar] [CrossRef]
- Bennet, C.; Bettaiya, R.; Rajanna, S.; Baker, L.; Yallapragada, P.R.; Brice, J.J.; White, S.L.; Bokara, K.K. Region specific increase in the antioxidant enzymes and lipid peroxidation products in the brain of rats exposed to lead. Free Radic. Res. 2007, 41, 267–273. [Google Scholar] [CrossRef]
- Madsen, B.K.; Zetner, D.; Møller, A.M.; Rosenberg, J. Melatonin for preoperative and postoperative anxiety in adults. Cochrane Database Syst. Rev. 2020, 8, 12. [Google Scholar]
- Lv, W.J.; Liu, C.; Yu, L.Z.; Zhou, J.H.; Li, Y.; Xiong, Y.; Guo, A.; Chao, L.M.; Qu, Q.; Wei, G.W.; et al. Melatonin Alleviates Neuroinflammation and Metabolic Disorder in DSS-Induced Depression Rats. Oxid. Med. Cell. Longev. 2020, 2020, 1241894. [Google Scholar] [CrossRef]
- He, R.; Cui, M.; Lin, H.; Zhao, L.; Wang, J.; Chen, S.; Shao, Z. Melatonin resists oxidative stress-induced apoptosis in nucleus pulposus cells. Life Sci. 2018, 199, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Zakria, M.; Ahmad, N.; Al Kury, L.T.; Alattar, A.; Uddin, Z.; Siraj, S.; Ullah, S.; Alshaman, R.; Khan, M.I.; Shah, F.A. Melatonin rescues the mice brain against cisplatin-induced neurodegeneration, an insight into antioxidant and anti-inflammatory effects. Neurotoxicology 2021, 87, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saxena, G.; Bharti, S.; Kamat, P.K.; Sharma, S.; Nath, C. Melatonin alleviates memory deficits and neuronal degeneration induced by intracerebroventricular administration of streptozotocin in rats. Pharmacol. Biochem. Behav. 2010, 94, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Zhang, X.; Wenk, M.R.; Goh, E.L.; Silver, D.L. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Muscaritoli, M. The Impact of Nutrients on Mental Health and Well-Being: Insights from the Literature. Front. Nutr. 2021, 8, 656290. [Google Scholar] [CrossRef]
- Robinson, D.G.; Gallego, J.A.; John, M.; Hanna, L.A.; Zhang, J.P.; Birnbaum, M.L.; Greenberg, J.; Naraine, M.; Peters, B.D.; McNamara, R.K.; et al. A potential role for adjunctive omega-3 polyunsaturated fatty acids for depression and anxiety symptoms in recent onset psychosis: Results from a 16 week randomized placebo-controlled trial for participants concurrently treated with risperidone. Schizophr. Res. 2019, 204, 295–303. [Google Scholar] [CrossRef]
- Larrieu, T.; Layé, S. Food for Mood: Relevance of Nutritional Omega-3 Fatty Acids for Depression and Anxiety. Front. Physiol. 2018, 9, 1047. [Google Scholar] [CrossRef]
- Neto, J.; Jantsch, J.; de Oliveira, S.; Braga, M.F.; Castro, L.F.D.S.; Diniz, B.F.; Moreira, J.C.F.; Giovenardi, M.; Porawski, M.; Guedes, R.P. DHA/EPA supplementation decreases anxiety-like behaviour, but it does not ameliorate metabolic profile in obese male rats. Br. J. Nutr. 2021, 128, 964–974. [Google Scholar] [CrossRef]
- Tanprasertsuk, J.; Mohn, E.S.; Matthan, N.R.; Lichtenstein, A.H.; Barger, K.; Vishwanathan, R.; Johnson, M.A.; Poon, L.W.; Johnson, E.J. Serum Carotenoids, Tocopherols, Total n-3 Polyunsaturated Fatty Acids, and n-6/n-3 Polyunsaturated Fatty Acid Ratio Reflect Brain Concentrations in a Cohort of Centenarians. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 306–314. [Google Scholar] [CrossRef]
- Daley, C.; Patterson, A.; Sibbritt, D.; MacDonald-Wicks, L. Unsaturated fat intakes and mental health outcomes in young women from the Australian Longitudinal Study on Women’s Heath. Public. Health Nutr. 2015, 18, 546–553. [Google Scholar] [CrossRef]
- Fatemi, F.; Siassi, F.; Qorbani, M.; Sotoudeh, G. Higher dietary fat quality is associated with lower anxiety score in women: A cross-sectional study. Ann. Gen. Psychiatry 2020, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R.; Kellogg, M.D.; Kramer, F.M.; Bathalon, G.P.; Lesher, L.L. Lipid and other plasma markers are associated with anxiety, depression, and fatigue. Health Psychol. 2012, 31, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Maroni, M.J.; Capri, K.M.; Arruda, N.L.; Gelineau, R.R.; Deane, H.V.; Concepcion, H.A.; DeCourcey, H.; Monteiro De Pina, I.K.; Cushman, A.V.; Chasse, M.H.; et al. Substrain specific behavioral responses in male C57BL/6N and C57BL/6J mice to a shortened 21-hour day and high-fat diet. Chronobiol. Int. 2020, 37, 809–823. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter [mmol/L] | C12/12 | DM12/12 | C6/18 | DM6/18 |
|---|---|---|---|---|
| Total cholesterol | 1.04 ± 0.09 | 1.06 ± 0.06 | 1.08 ± 0.08 | 1.11 ± 0.10 |
| TRG | 0.65 ± 0.14 | 0.87 ± 0.17 ** | 0.66 ± 0.08 | 0.58 ± 0.12 ## |
| HDL | 0.52 ± 0.05 | 0.45 ± 0.04 | 0.46 ± 0.03 | 0.63 ± 0.09 ** ## |
| C16:0 | 23.65 ± 1.55 | 22.83 ± 3.82 | 23.96 ± 2.91 | 23.62 ± 2.28 |
| C18:0 | 12.36 ± 2.29 | 12.95 ± 1.41 | 11.76 ± 1.95 | 11.18 ± 2.05 |
| C16:1 | 0.42 ± 0.06 | 0.42 ± 0.62 | 0.54 ± 0.09 | 0.53 ± 0.12 |
| C18:1ω9 | 9.55 ± 2.74 | 8.34 ± 0.85 | 10.78 ± 1.41 | 12.16 ± 3.97 ## |
| C18:1ω7 | 2.04 ± 0.26 | 1.05 ± 0.27 ** | 1.81 ± 0.31 | 1.93 ± 0.62 ## |
| C18:2 | 28.97 ± 2.36 | 28.77 ± 4.07 | 26.57 ± 3.71 | 28.32 ± 3.07 |
| C18:3ω6 | 0.12 ± 0.04 | 0.25 ± 0.12 | 0.15 ± 0.03 | 0.24 ± 0.18 |
| C20:3 | 0.50 ± 0.08 | 0.44 ± 0.04 | 0.62 ± 0.17 | 0.66 ± 0.16 ## |
| C20:4 | 19.66 ± 2.07 | 19.87 ± 2.15 | 19.99 ± 2.73 | 20.68 ± 3.73 |
| C22:4 | 0.43 ± 0.08 | 0.42 ± 0.07 | 0.39 ± 0.05 | 0.54 ± 0.15 |
| C18:3ω3 | 0.32 ± 0.14 | 0.32 ± 0.11 | 0.28 ± 0.04 | 0.29 ± 0.08 |
| C20:5 | 0.06 ± 0.03 | 0.09 ± 0.02 | 0.08 ± 0.05 | 0.07 ± 0.03 |
| C22:5 | 0.45 ± 0.16 | 0.37 ± 0.07 | 0.35 ± 0.05 | 0.45 ± 0.13 |
| C22:6 | 1.27 ± 0.31 | 0.99 ± 0.19 | 1.18 ± 0.15 | 1.45 ± 0.38 ## |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasović, D.D.; Vesković, M.; Šutulović, N.; Hrnčić, D.; Takić, M.; Jerotić, Đ.; Matić, M.; Stanojlović, O.; Ivković, S.; Jovanović Macura, I.; et al. Shortened Daily Photoperiod Alleviates Anxiety-like Behaviour by Antioxidant Effect and Changes Serum Fatty Acid Profile in Diabetic Rats. J. Pers. Med. 2023, 13, 744. https://doi.org/10.3390/jpm13050744
Vasović DD, Vesković M, Šutulović N, Hrnčić D, Takić M, Jerotić Đ, Matić M, Stanojlović O, Ivković S, Jovanović Macura I, et al. Shortened Daily Photoperiod Alleviates Anxiety-like Behaviour by Antioxidant Effect and Changes Serum Fatty Acid Profile in Diabetic Rats. Journal of Personalized Medicine. 2023; 13(5):744. https://doi.org/10.3390/jpm13050744
Chicago/Turabian StyleVasović, Dolika D., Milena Vesković, Nikola Šutulović, Dragan Hrnčić, Marija Takić, Đurđa Jerotić, Marija Matić, Olivera Stanojlović, Sanja Ivković, Irena Jovanović Macura, and et al. 2023. "Shortened Daily Photoperiod Alleviates Anxiety-like Behaviour by Antioxidant Effect and Changes Serum Fatty Acid Profile in Diabetic Rats" Journal of Personalized Medicine 13, no. 5: 744. https://doi.org/10.3390/jpm13050744