
Evaluation of CNPase and TGFβ1/Smad Signalling Pathway Molecule Expression in Sinus Epithelial Tissues of Patients with Chronic Rhinosinusitis with (CRSwNP) and without Nasal Polyps (CRSsNP)
 , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Immunohistochemistry
2.3. Immunofluorescence Tissue Staining
2.4. Statistical Analysis
3. Results
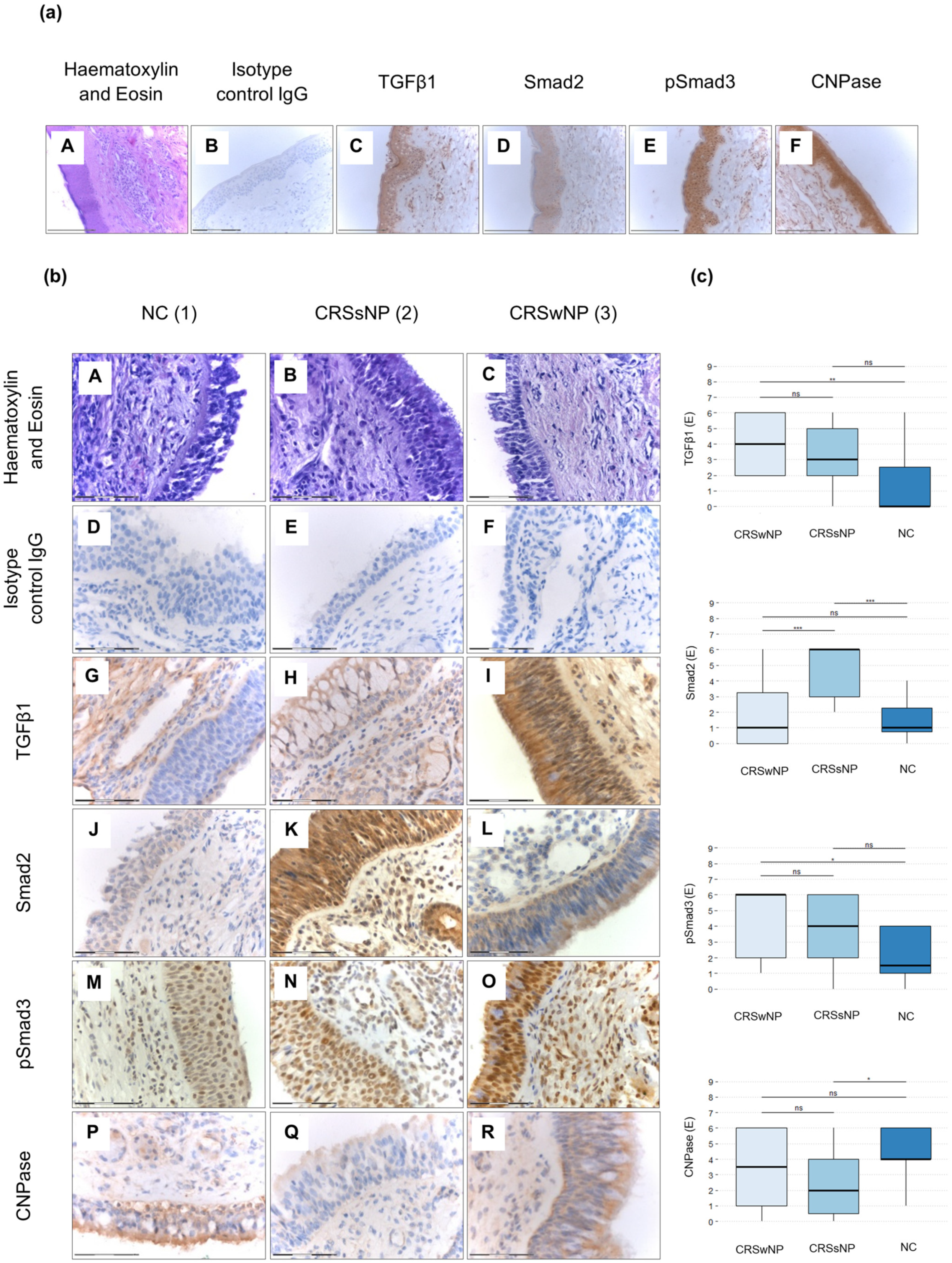
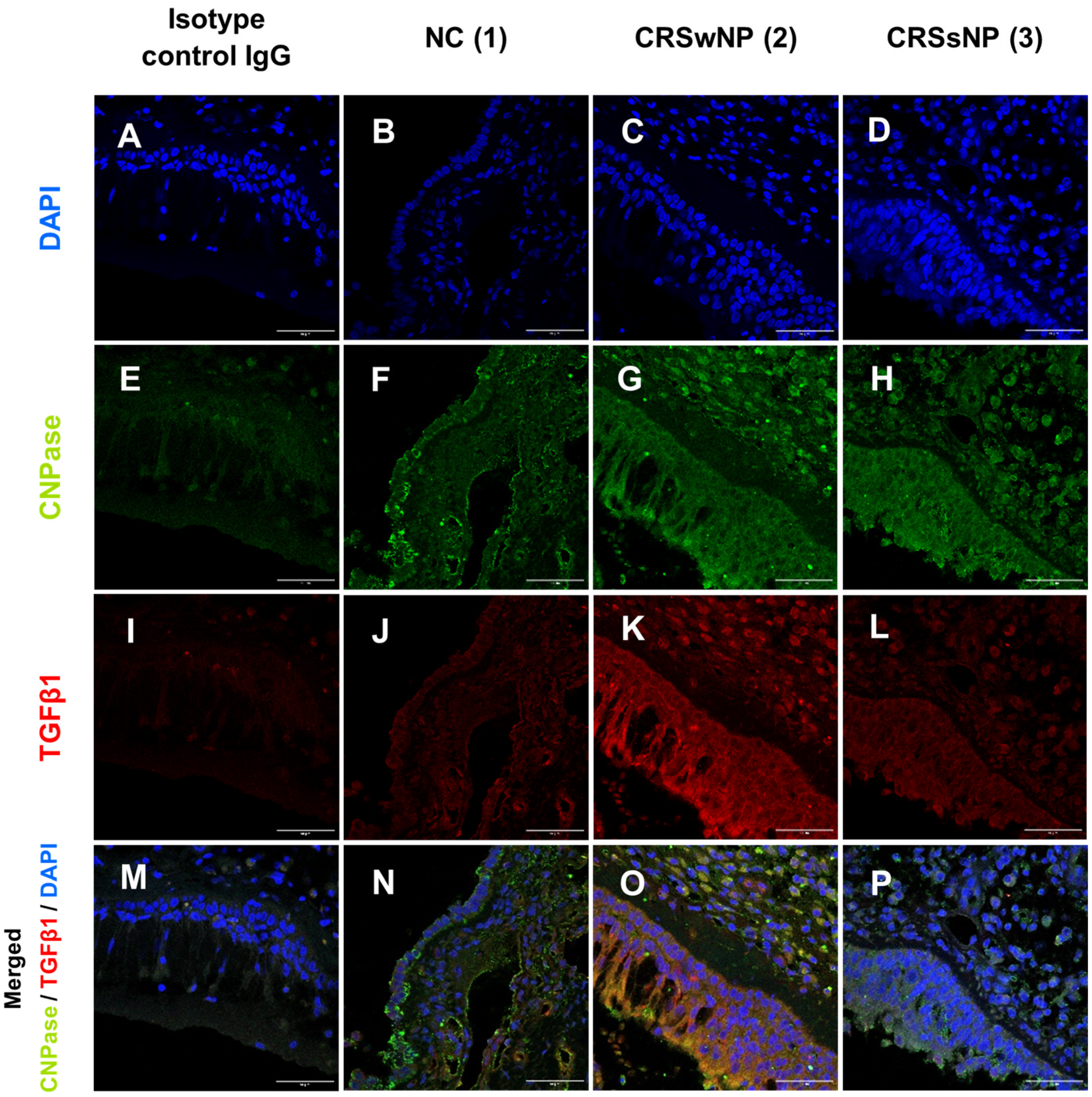
3.1. Levels of Smad2, pSmad3, TGFβ1, and CNPase Antigens in the Upper Airway Epithelium and Blood Eosinophils of CRSsNP, CRSwNP, and NC Patients
3.2. Expression Levels of Smad2, pSmad3, TGFβ1, and CNPase Correlate Positively with Each Other within Specific Patient Groups, Clinically Observed Symptoms, and Selected Questions from SNOT-20 and EPOS 2020 Questionnaires
3.2.1. Correlations within Control Patients
3.2.2. Correlations within the Group of Patients with CRSsNP
3.2.3. Correlations within the Group of Patients with CRSwNP
3.3. Age of Patients Correlated with the Subtype of CRS and CNPase Expression Level


4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, C. Chronic Rhinosinusitis with Nasal Polyps. N. Engl. J. Med. 2019, 381, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Levi, L.; Havazelet, S.; Reuven, Y.; Elmograbi, A.; Badir, S.; Shraga, Y.; Nakache, G.; Soudry, E. Patterns of recurrence in patients with CRSwNP who underwent complete FESS. Eur. Arch. Otorhinolaryngol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, C.; Masieri, S.; Begvarfaj, E.; Loperfido, A.; Baroncelli, S.; Cascone, F.; Ciofalo, A. Long-Term Perspectives on Chronic Rhinosinusitis with Nasal Polyps: Evaluating Recurrence Rates after Functional Endoscopic Sinus Surgery in the Biologics Era-A 5-Year Follow-Up Study. J. Pers. Med. 2024, 14, 297. [Google Scholar] [CrossRef]
- Calvanese, L.; Fabbris, C.; Brescia, G.; Di Pasquale Fiasca, V.M.; Deretti, A.; Finozzi, F.; Franz, L.; Frigo, A.C.; Marioni, G. Polyps’ Extension and Recurrence in Different Endotypes of Chronic Rhinosinusitis: A Series of 449 Consecutive Patients. J. Clin. Med. 2024, 13, 1125. [Google Scholar] [CrossRef]
- Nakayama, T.; Lee, I.T.; Le, W.; Tsunemi, Y.; Borchard, N.A.; Zarabanda, D.; Dholakia, S.S.; Gall, P.A.; Yang, A.; Kim, D.; et al. Inflammatory molecular endotypes of nasal polyps derived from White and Japanese populations. J. Allergy Clin. Immunol. 2022, 149, 1296–1308.e6. [Google Scholar] [CrossRef]
- Giombi, F.; Carron-Herrero, A.; Pirola, F.; Paoletti, G.; Nappi, E.; Russo, E.; De Virgilio, A.; Mercante, G.; Canonica, G.W.; Spriano, G.; et al. Prevalence of familial link in patients affected by chronic rhinosinusitis with nasal polyposis. Int. Forum Allergy Rhinol. 2022, 12, 1562–1565. [Google Scholar] [CrossRef]
- Oakley, G.M.; Curtin, K.; Orb, Q.; Schaefer, C.; Orlandi, R.R.; Alt, J.A. Familial risk of chronic rhinosinusitis with and without nasal polyposis: Genetics or environment. Int. Forum Allergy Rhinol. 2015, 5, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.X.; Mandanas, M.V.; Djeddi, S.; Fernandez-Salinas, D.; Gutierrez-Arcelus, M.; Barrett, N.A. Increased glycolysis and cellular crosstalk in eosinophilic chronic rhinosinusitis with nasal polyps. Front. Immunol. 2024, 15, 1321560. [Google Scholar] [CrossRef]
- Brar, T.; Baheti, S.; Marino, M.J.; Kita, H.; Lal, D. Genome-wide Epigenetic Study of Chronic Rhinosinusitis Tissues Reveals Dysregulated Inflammatory, Immunologic and Remodeling Pathways. Am. J. Rhinol. Allergy 2023, 37, 692–704. [Google Scholar] [CrossRef]
- Chen, J.; Chen, S.; Gong, G.; Yang, F.; Chen, J.; Wang, Y. Inhibition of IL-4/STAT6/IRF4 signaling reduces the epithelial-mesenchymal transition in eosinophilic chronic rhinosinusitis with nasal polyps. Int. Immunopharmacol. 2023, 121, 110554. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Li, X.; Lai, X.; Chen, X.; Li, Y.; Yao, Z.; Huang, Z.; Huang, J.; Chang, L.; Zhang, G. Interleukin-19 upregulates fibronectin and collagen I expression via the NF-kappaB-Smad2/3 pathway in fibroblasts of patients with chronic rhinosinusitis. Inflamm. Res. 2023, 72, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.S.; Bassiouni, A.; Ramezanpour, M.; Finnie, J.; Chegeni, N.; Colella, A.D.; Chataway, T.K.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. Proteomic analysis of nasal mucus samples of healthy patients and patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2021, 147, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Bankova, L.G.; Barrett, N.A. Epithelial cell function and remodeling in nasal polyposis. Ann. Allergy Asthma Immunol. 2020, 124, 333–341. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Cohen, N.A.; Barrett, N.A. Epithelial dysregulation in chronic rhinosinusitis with nasal polyposis (CRSwNP) and aspirin-exacerbated respiratory disease (AERD). J. Allergy Clin. Immunol. 2021, 148, 1161–1164. [Google Scholar] [CrossRef]
- Wang, D.Y.; Li, Y.; Yan, Y.; Li, C.; Shi, L. Upper airway stem cells: Understanding the nose and role for future cell therapy. Curr. Allergy Asthma Rep. 2015, 15, 490. [Google Scholar] [CrossRef] [PubMed]
- Shieh, J.M.; Tsai, Y.J.; Chi, J.C.; Wu, W.B. TGFbeta mediates collagen production in human CRSsNP nasal mucosa-derived fibroblasts through Smad2/3-dependent pathway and CTGF induction and secretion. J. Cell Physiol. 2019, 234, 10489–10499. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.H.; Shin, J.M.; Yang, H.W.; Lee, H.M.; Park, I.H. TGF-beta1-induced HSP47 regulates extracellular matrix accumulation via Smad2/3 signaling pathways in nasal fibroblasts. Sci. Rep. 2019, 9, 15563. [Google Scholar] [CrossRef]
- Batlle, E.; Massague, J. Transforming Growth Factor-beta Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef]
- Shelke, G.V.; Yin, Y.; Jang, S.C.; Lasser, C.; Wennmalm, S.; Hoffmann, H.J.; Li, L.; Gho, Y.S.; Nilsson, J.A.; Lotvall, J. Endosomal signalling via exosome surface TGFbeta-1. J. Extracell. Vesicles 2019, 8, 1650458. [Google Scholar] [CrossRef]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Pietrowska, M.; Widlak, P.; Hinck, C.S.; Gluszko, A.; Szczepanski, M.J.; Karmer, T.; Kallinger, I.; et al. TGFbeta(+) small extracellular vesicles from head and neck squamous cell carcinoma cells reprogram macrophages towards a pro-angiogenic phenotype. J. Extracell. Vesicles 2022, 11, e12294. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.J.; Heydet, D.; Veldre, T.; Ghildyal, R. Transcriptomic changes during TGF-beta-mediated differentiation of airway fibroblasts to myofibroblasts. Sci. Rep. 2019, 9, 20377. [Google Scholar] [CrossRef] [PubMed]
- Strauss, L.; Bergmann, C.; Szczepanski, M.; Gooding, W.; Johnson, J.T.; Whiteside, T.L. A unique subset of CD4+CD25highFoxp3+ T cells secreting interleukin-10 and transforming growth factor-beta1 mediates suppression in the tumor microenvironment. Clin. Cancer Res. 2007, 13, 4345–4354. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Chen, Y.; Liu, H.; Ye, M.; Chu, L.; Wang, T. CD109 identified in circulating proteomics mitigates postoperative recurrence in chronic rhinosinusitis with nasal polyps by suppressing TGF-beta1-induced epithelial-mesenchymal transition. Int. Immunopharmacol. 2024, 130, 111793. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sun, K.; Tu, Y.; Li, P.; Hao, D.; Yu, P.; Chen, A.; Wan, Y.; Shi, L. miR-200a-3p regulates epithelial-mesenchymal transition and inflammation in chronic rhinosinusitis with nasal polyps by targeting ZEB1 via ERK/p38 pathway. Int. Forum Allergy Rhinol. 2024, 14, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Lee, S.H.; Jo, H.R.; Weon, S.; Jeon, C.; Park, M.K.; Kim, T.H.; Cho, S.H. Eosinophil-derived TGFbeta1 controls the new bone formation in chronic rhinosinusitis with nasal polyps. Rhinology 2023, 61, 338–347. [Google Scholar] [CrossRef]
- Park, J.H.; Shin, J.M.; Yang, H.W.; Park, I.H. DNMTs Are Involved in TGF-beta1-Induced Epithelial-Mesenchymal Transitions in Airway Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 3003. [Google Scholar] [CrossRef]
- Yang, Y.C.; Zhang, N.; Van Crombruggen, K.; Hu, G.H.; Hong, S.L.; Bachert, C. Transforming growth factor-beta1 in inflammatory airway disease: A key for understanding inflammation and remodeling. Allergy 2012, 67, 1193–1202. [Google Scholar] [CrossRef]
- Ungefroren, H.; Groth, S.; Sebens, S.; Lehnert, H.; Gieseler, F.; Fandrich, F. Differential roles of Smad2 and Smad3 in the regulation of TGF-beta1-mediated growth inhibition and cell migration in pancreatic ductal adenocarcinoma cells: Control by Rac1. Mol. Cancer 2011, 10, 67. [Google Scholar] [CrossRef]
- Pacini, E.S.A.; Satori, N.A.; Jackson, E.K.; Godinho, R.O. Extracellular cAMP-Adenosine Pathway Signaling: A Potential Therapeutic Target in Chronic Inflammatory Airway Diseases. Front. Immunol. 2022, 13, 866097. [Google Scholar] [CrossRef]
- Jackson, E.K. The 2′,3′-cAMP-adenosine pathway. Am. J. Physiol. Renal Physiol. 2011, 301, F1160–F1167. [Google Scholar] [CrossRef]
- Jackson, E.K.; Gillespie, D.G.; Mi, Z.; Cheng, D.; Bansal, R.; Janesko-Feldman, K.; Kochanek, P.M. Role of 2′,3′-cyclic nucleotide 3′-phosphodiesterase in the renal 2′,3′-cAMP-adenosine pathway. Am. J. Physiol. Renal Physiol. 2014, 307, F14–F24. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.K.; Ren, J.; Mi, Z. Extracellular 2′,3′-cAMP is a source of adenosine. J. Biol. Chem. 2009, 284, 33097–33106. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C.; Sasaki, M.; Lankford, K.L.; Gallo, V.; Kocsis, J.D. CNPase expression in olfactory ensheathing cells. J. Biomed. Biotechnol. 2011, 2011, 608496. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Y.; Wang, Y. 2′,3′-Cyclic-nucleotide 3′-phosphodiesterase contributes to epithelial-mesenchymal transition of lens epithelial cells through the notch signalling pathway. Cell Prolif. 2019, 52, e12707. [Google Scholar] [CrossRef] [PubMed]
- Regateiro, F.S.; Howie, D.; Nolan, K.F.; Agorogiannis, E.I.; Greaves, D.R.; Cobbold, S.P.; Waldmann, H. Generation of anti-inflammatory adenosine by leukocytes is regulated by TGF-beta. Eur. J. Immunol. 2011, 41, 2955–2965. [Google Scholar] [CrossRef] [PubMed]
- Schiller, M.; Dennler, S.; Anderegg, U.; Kokot, A.; Simon, J.C.; Luger, T.A.; Mauviel, A.; Bohm, M. Increased cAMP levels modulate transforming growth factor-beta/Smad-induced expression of extracellular matrix components and other key fibroblast effector functions. J. Biol. Chem. 2010, 285, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Vasiukov, G.; Menshikh, A.; Owens, P.; Novitskaya, T.; Hurley, P.; Blackwell, T.; Feoktistov, I.; Novitskiy, S.V. Adenosine/TGFbeta axis in regulation of mammary fibroblast functions. PLoS ONE 2021, 16, e0252424. [Google Scholar] [CrossRef]
- Vasiukov, G.; Novitskaya, T.; Zijlstra, A.; Owens, P.; Ye, F.; Zhao, Z.; Moses, H.L.; Blackwell, T.; Feoktistov, I.; Novitskiy, S.V. Myeloid Cell-Derived TGFbeta Signaling Regulates ECM Deposition in Mammary Carcinoma via Adenosine-Dependent Mechanisms. Cancer Res. 2020, 80, 2628–2638. [Google Scholar] [CrossRef]
- Coste, A.; Lefaucheur, J.P.; Wang, Q.P.; Lesprit, E.; Poron, F.; Peynegre, R.; Escudier, E. Expression of the transforming growth factor beta isoforms in inflammatory cells of nasal polyps. Arch. Otolaryngol. Head. Neck Surg. 1998, 124, 1361–1366. [Google Scholar] [CrossRef]
- Bifulco, M.; Laezza, C.; Stingo, S.; Wolff, J. 2′,3′-Cyclic nucleotide 3′-phosphodiesterase: A membrane-bound, microtubule-associated protein and membrane anchor for tubulin. Proc. Natl. Acad. Sci. USA 2002, 99, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Li, Z.; Alvarez, R., Jr.; Feng, X.H.; Goldschmidt-Clermont, P.J. Microtubule binding to Smads may regulate TGF beta activity. Mol. Cell 2000, 5, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, J.F.; Merritt, M.G., Jr.; Richards, M.L. Psychometric and clinimetric validity of the 20-Item Sino-Nasal Outcome Test (SNOT-20). Otolaryngol. Head Neck Surg. 2002, 126, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Pawankar, R.; Nonaka, M. Inflammatory mechanisms and remodeling in chronic rhinosinusitis and nasal polyps. Curr. Allergy Asthma Rep. 2007, 7, 202–208. [Google Scholar] [CrossRef]
- Nnabue, O.E.; Pletcher, S.D.; Gurrola, J.G., 2nd; Goldberg, A.N.; Jordan, K.M.; Knox, S.M.; May, A.J. Septum submucosal glands exhibit aberrant morphology and reduced mucin production in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2021, 11, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liu, X.; Zhang, N.; Li, J.; Wen, Y.; Wei, Y.; Li, Z.; Lu, T.; Wen, W. Microvessel quantification by fully convolutional neural networks associated with type 2 inflammation in chronic rhinosinusitis. Ann. Allergy Asthma Immunol. 2022, 128, 697–704.e1. [Google Scholar] [CrossRef] [PubMed]
- Van Bruaene, N.; Derycke, L.; Perez-Novo, C.A.; Gevaert, P.; Holtappels, G.; De Ruyck, N.; Cuvelier, C.; Van Cauwenberge, P.; Bachert, C. TGF-beta signaling and collagen deposition in chronic rhinosinusitis. J. Allergy Clin. Immunol. 2009, 124, 253–259.e2. [Google Scholar] [CrossRef]
- Lucas, B.R.; Voegels, R.L.; do Amaral, J.B.; Bachi, A.L.L.; Pezato, R. BMP-7, MMP-9, and TGF-beta tissue remodeling proteins and their correlations with interleukins 6 and 10 in chronic rhinosinusitis. Eur. Arch. Otorhinolaryngol. 2021, 278, 4335–4343. [Google Scholar] [CrossRef]
- Hackett, T.L.; Warner, S.M.; Stefanowicz, D.; Shaheen, F.; Pechkovsky, D.V.; Murray, L.A.; Argentieri, R.; Kicic, A.; Stick, S.M.; Bai, T.R.; et al. Induction of epithelial-mesenchymal transition in primary airway epithelial cells from patients with asthma by transforming growth factor-beta1. Am. J. Respir. Crit. Care Med. 2009, 180, 122–133. [Google Scholar] [CrossRef]
- Konnecke, M.; Burmeister, M.; Pries, R.; Boscke, R.; Bruchhage, K.L.; Ungefroren, H.; Klimek, L.; Wollenberg, B. Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis: Differences Revealed Between Epithelial Cells from Nasal Polyps and Inferior Turbinates. Arch. Immunol. Ther. Exp. 2017, 65, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Carsuzaa, F.; Bequignon, E.; Bainaud, M.; Jegou, J.F.; Dufour, X.; Lecron, J.C.; Favot, L. Oncostatin M Counteracts the Fibrotic Effects of TGF-beta1 and IL-4 on Nasal-Polyp-Derived Fibroblasts: A Control of Fibrosis in Chronic Rhinosinusitis with Nasal Polyps? Int. J. Mol. Sci. 2022, 23, 6308. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, J.H.; Lee, S.A.; Lee, J.G.; Shin, J.M.; Lee, H.M. All-trans retinoic acid regulates TGF-beta1-induced extracellular matrix production via p38, JNK, and NF-kappaB-signaling pathways in nasal polyp-derived fibroblasts. Int. Forum Allergy Rhinol. 2020, 10, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Escudier, E.; Roudot-Thoraval, F.; Abd-Al Samad, I.; Peynegre, R.; Coste, A. Myofibroblast accumulation induced by transforming growth factor-beta is involved in the pathogenesis of nasal polyps. Laryngoscope 1997, 107, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Nocera, A.L.; Dillon, S.T.; Wu, D.; Libermann, T.A.; Bleier, B.S. Highly multiplexed proteomic analysis reveals significant tissue and exosomal coagulation pathway derangement in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2018, 8, 1438–1444. [Google Scholar] [CrossRef]
- Meng, X.M.; Huang, X.R.; Chung, A.C.; Qin, W.; Shao, X.; Igarashi, P.; Ju, W.; Bottinger, E.P.; Lan, H.Y. Smad2 protects against TGF-beta/Smad3-mediated renal fibrosis. J. Am. Soc. Nephrol. 2010, 21, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Yerneni, S.S.; Menshikova, E.V.; Gillespie, D.G.; Jackson, E.K.; Whiteside, T.L. Simultaneous Inhibition of Glycolysis and Oxidative Phosphorylation Triggers a Multi-Fold Increase in Secretion of Exosomes: Possible Role of 2′3′-cAMP. Sci. Rep. 2020, 10, 6948. [Google Scholar] [CrossRef]
- Nocera, A.L.; Mueller, S.K.; Workman, A.D.; Wu, D.; McDonnell, K.; Sadow, P.M.; Amiji, M.M.; Bleier, B.S. Cystatin SN is a potent upstream initiator of epithelial-derived type 2 inflammation in chronic rhinosinusitis. J. Allergy Clin. Immunol. 2022, 150, 872–881. [Google Scholar] [CrossRef]
- Shimizu, S.; Tojima, I.; Nakamura, K.; Arai, H.; Kouzaki, H.; Shimizu, T. Nasal polyp fibroblasts (NPFs)-derived exosomes are important for the release of vascular endothelial growth factor from cocultured eosinophils and NPFs. Auris Nasus Larynx 2022, 49, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Tan, K.S.; Guan, W.J.; Jiang, L.J.; Deng, J.; Gao, W.X.; Lee, Y.M.; Xu, Z.F.; Luo, X.; Liu, C.; et al. Proteomics profiling of epithelium-derived exosomes from nasal polyps revealed signaling functions affecting cellular proliferation. Respir. Med. 2020, 162, 105871. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, T.; Yan, Y.; Zhang, J.; Zhou, Y.; Pei, Y.; Yao, L.; You, B.; Chen, J. Exosomal miR-22-3p Derived from Chronic Rhinosinusitis with Nasal Polyps Regulates Vascular Permeability by Targeting VE-Cadherin. Biomed. Res. Int. 2020, 2020, 1237678. [Google Scholar] [CrossRef]
- Nocera, A.L.; Mueller, S.K.; Stephan, J.R.; Hing, L.; Seifert, P.; Han, X.; Lin, D.T.; Amiji, M.M.; Libermann, T.; Bleier, B.S. Exosome swarms eliminate airway pathogens and provide passive epithelial immunoprotection through nitric oxide. J. Allergy Clin. Immunol. 2018, 143, 1525–1535.e1. [Google Scholar] [CrossRef]
- Nocera, A.L.; Miyake, M.M.; Seifert, P.; Han, X.; Bleier, B.S. Exosomes mediate interepithelial transfer of functional P-glycoprotein in chronic rhinosinusitis with nasal polyps. Laryngoscope 2017, 127, E295–E300. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Nocera, A.L.; Workman, A.; Libermann, T.; Dillon, S.T.; Stegmann, A.; Wurm, J.; Iro, H.; Wendler, O.; Bleier, B.S. Significant polyomic and functional upregulation of the PAPP-A/IGFBP-4/5/IGF-1 axis in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2020, 10, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Lee, C.H.; Liang, S.S.; Hung, C.C.; Wu, Y.R.; Chien, C.Y.; Lee, C.H.; Chen, J.Y. Mucin 5AC is significantly upregulated in exosomes from the nasal lavage fluid and may promote the expression of COX-2, VEGF and MMP-9: An implication in nasal polyp pathogenesis. Rhinology 2021, 59, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, R.; Liang, W.; Qiu, H.; Yuan, T.; Wang, W.; Deng, H.; Kong, W.; Chen, J.; Bai, Y.; et al. Small extracellular vesicles facilitate epithelial-mesenchymal transition in chronic rhinosinusitis with nasal polyps via the miR-375-3p/QKI axis. Rhinology 2024, 62, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Logue, J.; Melville, V.M.; Ardanuy, J.; Frieman, M.B. 2′,3′ cyclic-nucleotide 3′-phosphodiesterase (CNP) inhibits SARS-CoV-2 virion assembly by blocking infection-induced mitochondria depolarization. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lee, S.W.; Kim, S.Y.; Moon, S.Y.; Yang, J.M.; Ha, E.K.; Jee, H.M.; Shin, J.I.; Cho, S.H.; Yon, D.K.; Suh, D.I. Estimating COVID-19 Infection and Severity Risks in Patients with Chronic Rhinosinusitis: A Korean Nationwide Cohort Study. J. Allergy Clin. Immunol. Pract. 2021, 9, 2262–2271.e2. [Google Scholar] [CrossRef] [PubMed]
- Waldstein, K.A.; Ganama, M.; Varga, S.M.; Tilley, S.; Hua, X. Topical Adenosine Inhibits Inflammation and Mucus Production in Viral Acute Rhinosinusitis. Laryngoscope 2022, 133, 2095–2103. [Google Scholar] [CrossRef]
- Hua, X.; Naselsky, W.C.; Bennett, W.D.; Ledent, C.; Senior, B.A.; Tilley, S.L. Adenosine increases nasal mucociliary clearance rate in mice through A2A and A2B adenosine receptors. Laryngoscope 2013, 123, 306–310. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, H.; Liu, Y.; Gao, K.; Zhang, J.; Fan, R.; Xie, S.; Xie, Z.; Wang, F.; Jiang, W. The Role of Serum Metabolomics in Distinguishing Chronic Rhinosinusitis With Nasal Polyp Phenotypes. Front. Mol. Biosci. 2020, 7, 593976. [Google Scholar] [CrossRef]
- DeConde, A.S.; Mace, J.C.; Levy, J.M.; Rudmik, L.; Alt, J.A.; Smith, T.L. Prevalence of polyp recurrence after endoscopic sinus surgery for chronic rhinosinusitis with nasal polyposis. Laryngoscope 2017, 127, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Ohno, I.; Lea, R.G.; Flanders, K.C.; Clark, D.A.; Banwatt, D.; Dolovich, J.; Denburg, J.; Harley, C.B.; Gauldie, J.; Jordana, M. Eosinophils in chronically inflamed human upper airway tissues express transforming growth factor beta 1 gene (TGF beta 1). J. Clin. Investig. 1992, 89, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Gudis, D.; Zhao, K.Q.; Cohen, N.A. Acquired cilia dysfunction in chronic rhinosinusitis. Am. J. Rhinol. Allergy 2012, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Shaari, J.; Claire, S.E.; Palmer, J.N.; Chiu, A.G.; Kennedy, D.W.; Cohen, N.A. Altered sinonasal ciliary dynamics in chronic rhinosinusitis. Am. J. Rhinol. 2006, 20, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Li, C.W.; Chao, S.S.; Yu, F.G.; Yu, X.M.; Liu, J.; Yan, Y.; Shen, L.; Gordon, W.; Shi, L.; et al. Impairment of cilia architecture and ciliogenesis in hyperplastic nasal epithelium from nasal polyps. J. Allergy Clin. Immunol. 2014, 134, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Chandra, R.K.; Pollak, J.; Kato, A.; Conley, D.B.; Peters, A.T.; Grammer, L.C.; Avila, P.C.; Kern, R.C.; Stewart, W.F.; et al. Incidence and associated premorbid diagnoses of patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2013, 131, 1350–1360. [Google Scholar] [CrossRef]
- Tatar, M.; Plevkova, J.; Brozmanova, M.; Pecova, R.; Kollarik, M. Mechanisms of the cough associated with rhinosinusitis. Pulm. Pharmacol. Ther. 2009, 22, 121–126. [Google Scholar] [CrossRef]
- Lai, L.Y.S.; Gracie, N.P.; Gowripalan, A.; Howell, L.M.; Newsome, T.P. SMAD proteins: Mediators of diverse outcomes during infection. Eur. J. Cell Biol. 2022, 101, 151204. [Google Scholar] [CrossRef]
- Kuo, C.L.; Yen, Y.C.; Chang, W.P.; Shiao, A.S. Association Between Middle Ear Cholesteatoma and Chronic Rhinosinusitis. JAMA Otolaryngol. Head Neck Surg. 2017, 143, 757–763. [Google Scholar] [CrossRef]
- Leichtle, A.; Klenke, C.; Ebmeyer, J.; Daerr, M.; Bruchhage, K.L.; Hoffmann, A.S.; Ryan, A.F.; Wollenberg, B.; Sudhoff, H. NOD-Like Receptor Signaling in Cholesteatoma. Biomed. Res. Int. 2015, 2015, 408169. [Google Scholar] [CrossRef]
- Ferguson, M. Rhinosinusitis in oral medicine and dentistry. Aust. Dent. J. 2014, 59, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Gu, Q. Expression and Clinical Significance of Mucin Gene in Chronic Rhinosinusitis. Curr. Allergy Asthma Rep. 2020, 20, 63. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Chu, H.S.; Lee, J.Y.; Hwang, S.J.; Lee, S.H.; Lee, H.M. Up-regulation of MUC5AC and MUC5B mucin genes in chronic rhinosinusitis. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Stoikes, N.F.; Dutton, J.M. The effect of endoscopic sinus surgery on symptoms of eustachian tube dysfunction. Am. J. Rhinol. 2005, 19, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Tangbumrungtham, N.; Patel, V.S.; Thamboo, A.; Patel, Z.M.; Nayak, J.V.; Ma, Y.; Choby, G.; Hwang, P.H. The prevalence of Eustachian tube dysfunction symptoms in patients with chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2018, 8, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Young, T.; Finn, L.; Kim, H. Nasal obstruction as a risk factor for sleep-disordered breathing. The University of Wisconsin Sleep and Respiratory Research Group. J. Allergy Clin. Immunol. 1997, 99, S757–S762. [Google Scholar] [CrossRef]
- Jiang, R.S.; Liang, K.L.; Hsin, C.H.; Su, M.C. The impact of chronic rhinosinusitis on sleep-disordered breathing. Rhinology 2016, 54, 75–79. [Google Scholar] [CrossRef]
- Kubota, T.; Fang, J.; Kushikata, T.; Krueger, J.M. Interleukin-13 and transforming growth factor-beta1 inhibit spontaneous sleep in rabbits. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R786–R792. [Google Scholar] [CrossRef]
- Alt, J.A.; Sautter, N.B.; Mace, J.C.; Detwiller, K.Y.; Smith, T.L. Antisomnogenic cytokines, quality of life, and chronic rhinosinusitis: A pilot study. Laryngoscope 2014, 124, E107–E114. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | NC Patients (n = 8) | CRSwNP Patients (n = 20) | CRSsNP Patients (n = 23) |
|---|---|---|---|
| Sex | |||
| Male | 6 | 12 | 15 |
| Female | 2 | 8 | 8 |
| Age | |||
| Range | 19–44 | 26–71 | 19–65 |
| Median | 32 | 48.5 | 35 |
| Allergy | 0 | 8 | 9 |
| Asthma | 1 | 3 | 2 |
| Aspirin sensitivity/ non-exacerbated respiratory disease (N-ERD) | 0 | 2 | 0 |
| Eosinophiles [%] (average ± SD) | 2.1 (±1.183) | 6.118 (±4.94) | 3.275 (±2.236) |
| Eosinophiles [103/μL] (average ± SD) | 0.129 (±0.063) | 0.427 (±0.404) | 0.229 (±0.178) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piszczatowska, K.; Czerwaty, K.; Dżaman, K.; Jermakow, N.; Brzost, J.; Kantor, I.; Ludwig, N.; Szczepański, M.J. Evaluation of CNPase and TGFβ1/Smad Signalling Pathway Molecule Expression in Sinus Epithelial Tissues of Patients with Chronic Rhinosinusitis with (CRSwNP) and without Nasal Polyps (CRSsNP). J. Pers. Med. 2024, 14, 894. https://doi.org/10.3390/jpm14090894
Piszczatowska K, Czerwaty K, Dżaman K, Jermakow N, Brzost J, Kantor I, Ludwig N, Szczepański MJ. Evaluation of CNPase and TGFβ1/Smad Signalling Pathway Molecule Expression in Sinus Epithelial Tissues of Patients with Chronic Rhinosinusitis with (CRSwNP) and without Nasal Polyps (CRSsNP). Journal of Personalized Medicine. 2024; 14(9):894. https://doi.org/10.3390/jpm14090894
Chicago/Turabian StylePiszczatowska, Katarzyna, Katarzyna Czerwaty, Karolina Dżaman, Natalia Jermakow, Jacek Brzost, Ireneusz Kantor, Nils Ludwig, and Mirosław J. Szczepański. 2024. "Evaluation of CNPase and TGFβ1/Smad Signalling Pathway Molecule Expression in Sinus Epithelial Tissues of Patients with Chronic Rhinosinusitis with (CRSwNP) and without Nasal Polyps (CRSsNP)" Journal of Personalized Medicine 14, no. 9: 894. https://doi.org/10.3390/jpm14090894
APA StylePiszczatowska, K., Czerwaty, K., Dżaman, K., Jermakow, N., Brzost, J., Kantor, I., Ludwig, N., & Szczepański, M. J. (2024). Evaluation of CNPase and TGFβ1/Smad Signalling Pathway Molecule Expression in Sinus Epithelial Tissues of Patients with Chronic Rhinosinusitis with (CRSwNP) and without Nasal Polyps (CRSsNP). Journal of Personalized Medicine, 14(9), 894. https://doi.org/10.3390/jpm14090894